
Downregulation of sCD40 and sCTLA4 in Recovered COVID-19 Patients with Comorbidities

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Material and Methods
2.1. SARS-CoV-2-Specific IgG Levels
2.2. Subjects
2.3. ELISA
2.4. Statistical Analysis
3. Results
3.1. Titers of SARS-CoV-2 N-Specific IgG
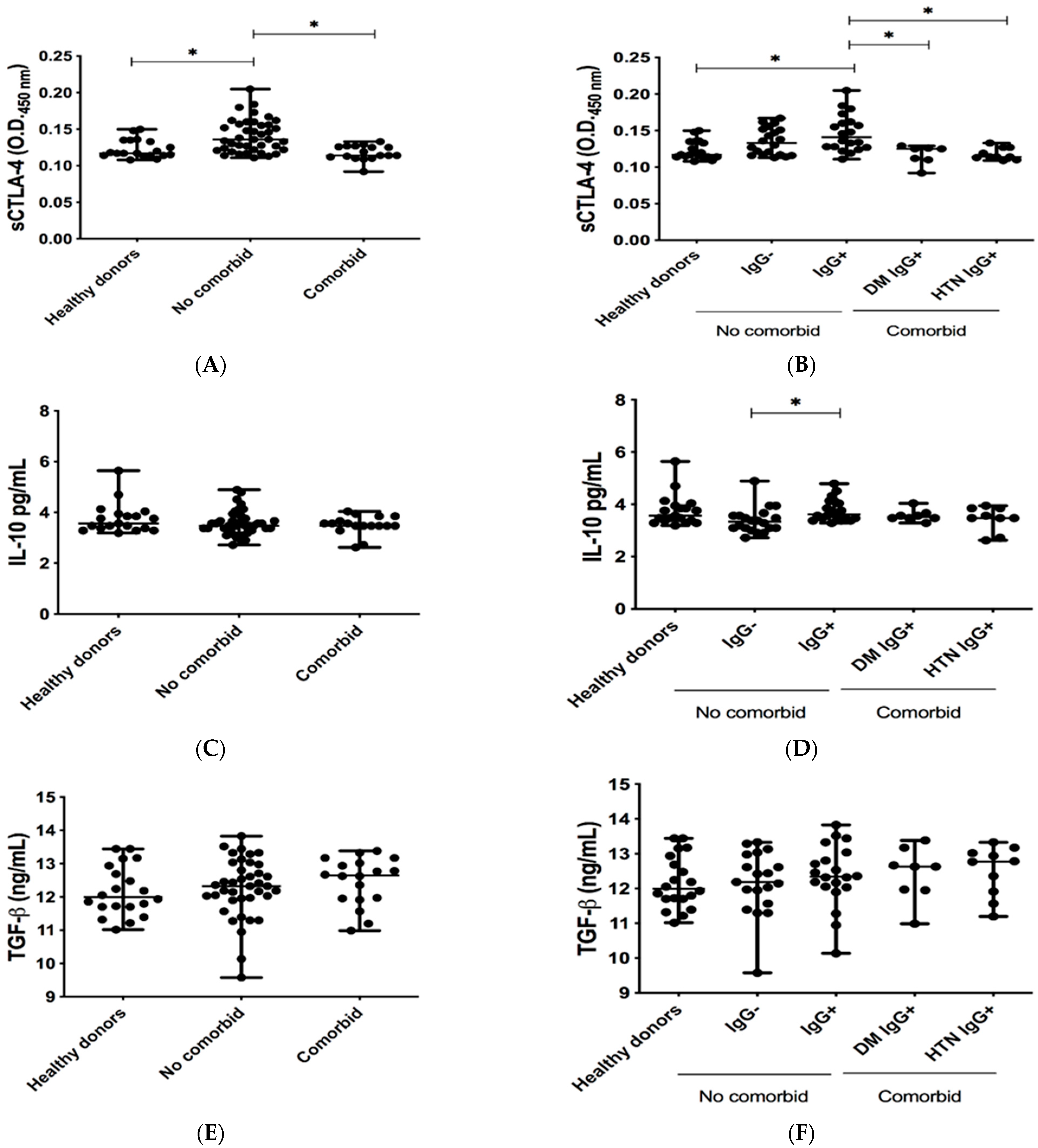
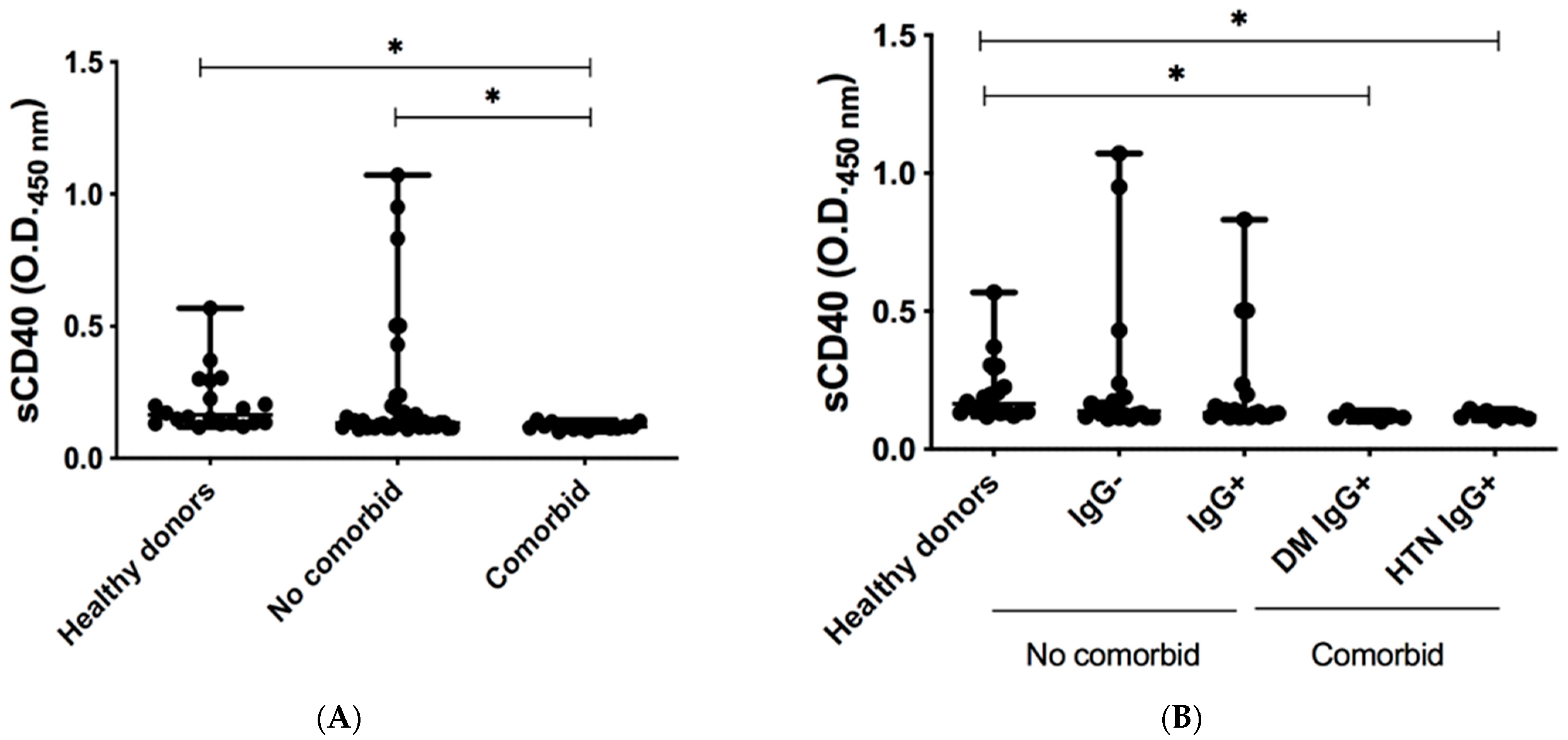
3.2. Levels of Co-Inhibitory Molecules in Serum of COVID-19-Recovered Subjects
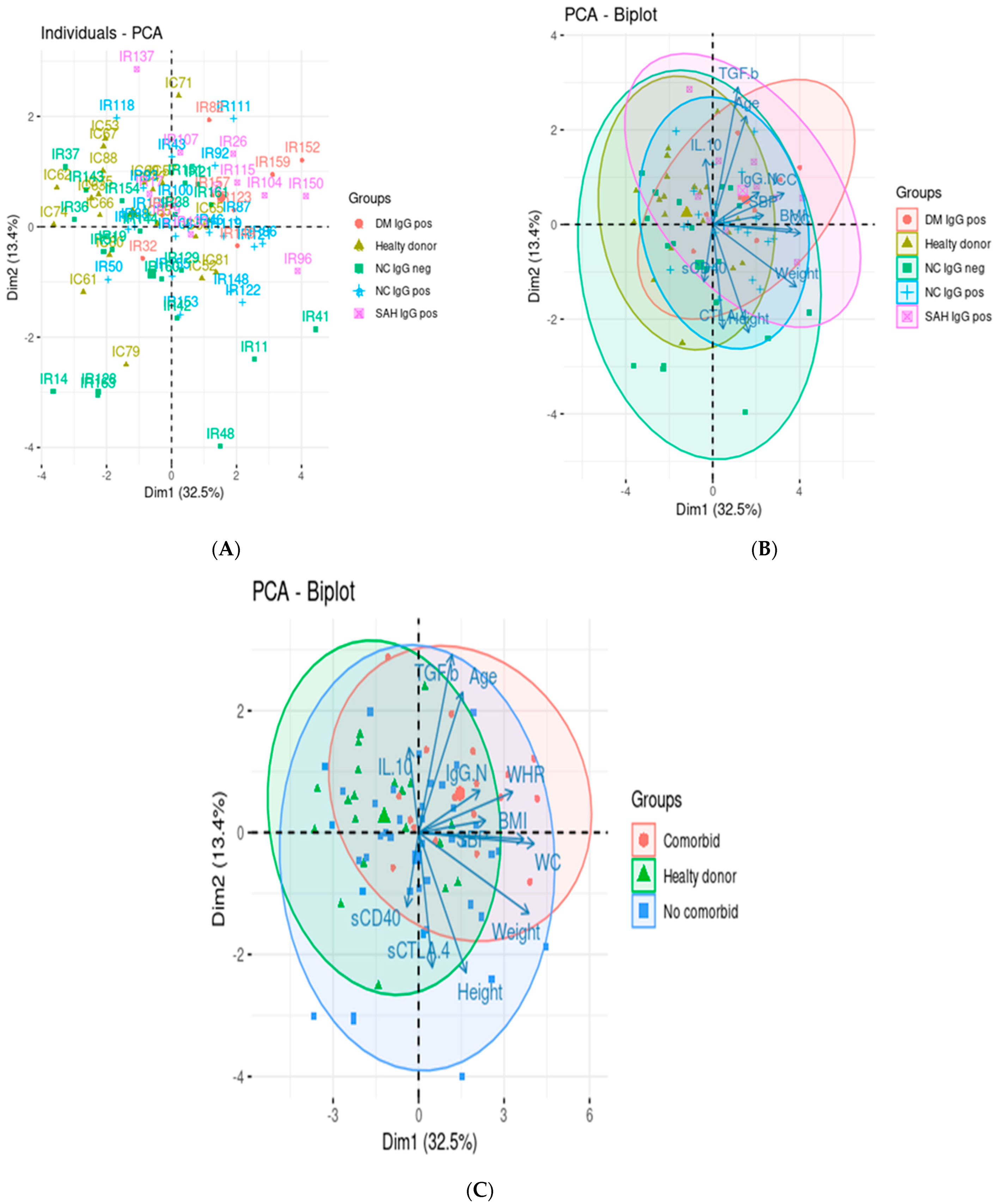
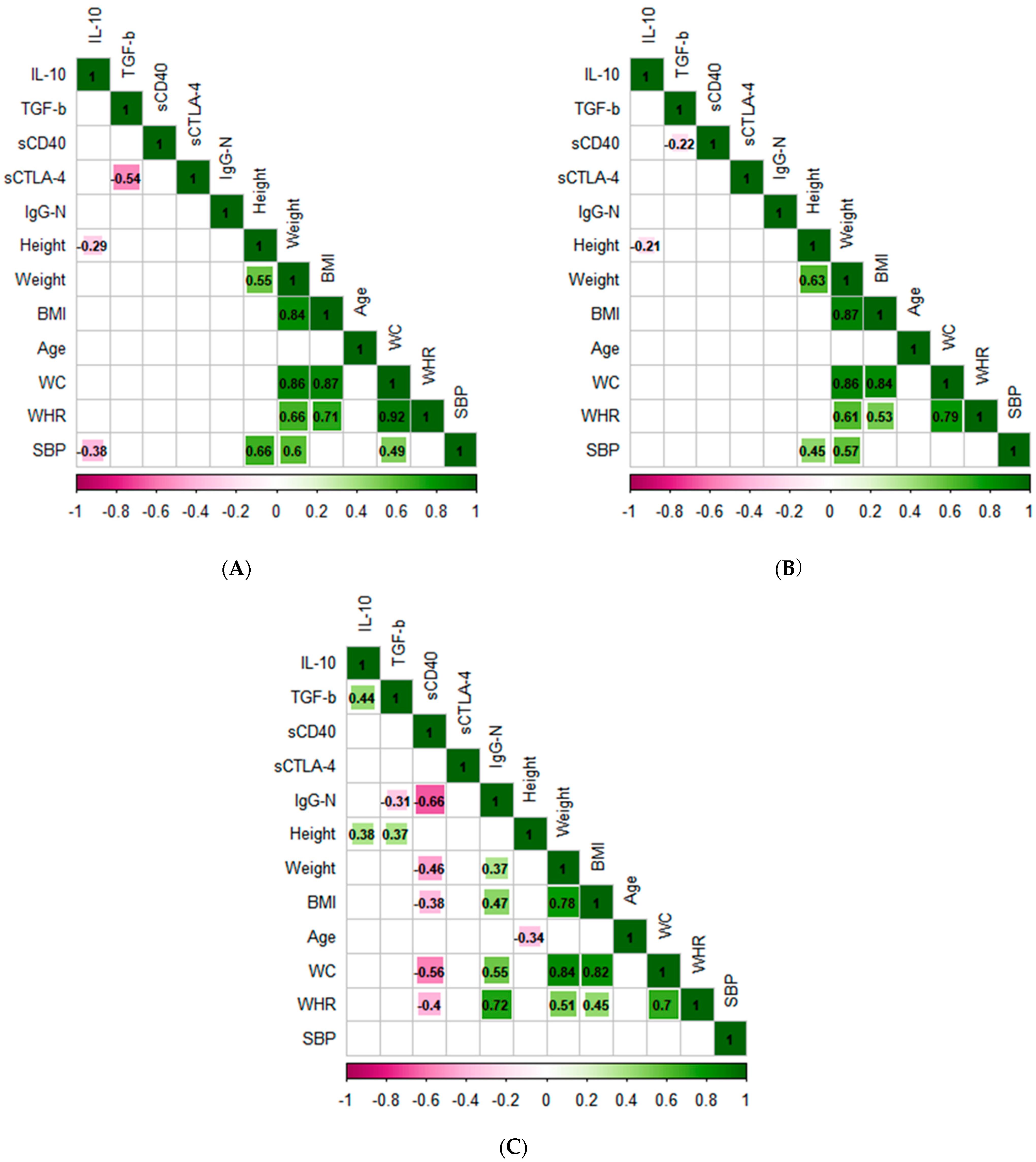
3.3. Principal Component Analysis and Correlation Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, T.; Wu, Q.; Zhang, Z. Probable Pangolin Origin of SARS-CoV-2 Associated with the COVID-19 Outbreak. Curr. Biol. 2020, 30, 1578. [Google Scholar] [CrossRef] [PubMed]
- Kermali, M.; Khalsa, R.K.; Pillai, K.; Ismail, Z.; Harky, A. The role of biomarkers in diagnosis of COVID-19—A systematic review. Life Sci. 2020, 254, 117788. [Google Scholar] [CrossRef] [PubMed]
- Webb, B.J.; Peltan, I.D.; Jensen, P.; Hoda, D.; Hunter, B.; Silver, A.; Starr, N.; Buckel, W.; Grisel, N.; Hummel, E.; et al. Clinical criteria for COVID-19-associated hyperinflammatory syndrome: A cohort study. Lancet Rheumatol. 2020, 2, e754–e763. [Google Scholar] [CrossRef]
- Weatherhead, J.E.; Clark, E.; Vogel, T.P.; Atmar, R.L.; Kulkarni, P.A. Inflammatory syndromes associated with SARS-CoV-2 infection: Dysregulation of the immune response across the age spectrum. J. Clin. Investig. 2020, 130, 6194–6197. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zheng, Y.; Gou, X.; Pu, K.; Chen, Z.; Guo, Q.; Ji, R.; Wang, H.; Wang, Y.; Zhou, Y. Prevalence of comorbidities and its effects in patients infected with SARS-CoV-2: A systematic review and meta-analysis. Int. J. Infect. Dis. 2020, 94, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Callender, L.A.; Curran, M.; Bates, S.M.; Mairesse, M.; Weigandt, J.; Betts, C.J. The Impact of Pre-existing Comorbidities and Therapeutic Interventions on COVID-19. Front. Immunol. 2020, 11, 1991. [Google Scholar] [CrossRef]
- Liu, J.; Li, S.; Liu, J.; Liang, B.; Wang, X.; Wang, H.; Li, W.; Tong, Q.; Yi, J.; Zhao, L.; et al. Longitudinal characteristics of lymphocyte responses and cytokine profiles in the peripheral blood of SARS-CoV-2 infected patients. EBioMedicine 2020, 55, 102763. [Google Scholar] [CrossRef]
- Zhang, C.; Wu, Z.; Li, J.W.; Zhao, H.; Wang, G.Q. Cytokine release syndrome in severe COVID-19: Interleukin-6 receptor antagonist tocilizumab may be the key to reduce mortality. Int. J. Antimicrob. Agents 2020, 55, 105954. [Google Scholar] [CrossRef]
- Qin, C.; Zhou, L.; Hu, Z.; Zhang, S.; Yang, S.; Tao, Y.; Xie, C.; Ma, K.; Shang, K.; Wang, W.; et al. Dysregulation of Immune Response in Patients With Coronavirus 2019 (COVID-19) in Wuhan, China. Clin. Infect. Dis. 2020, 71, 762–768. [Google Scholar] [CrossRef]
- Blanco-Melo, D.; Nilsson-Payant, B.E.; Liu, W.C.; Uhl, S.; Hoagland, D.; Møller, R.; Jordan, T.X.; Oishi, K.; Panis, M.; Sachs, D.; et al. Imbalanced Host Response to SARS-CoV-2 Drives Development of COVID-19. Cell 2020, 181, 1036–1045.e9. [Google Scholar] [CrossRef]
- Cohn, M. What roles do regulatory T cells play in the control of the adaptive immune response? Int. Immunol. 2008, 20, 1107–1118. [Google Scholar] [CrossRef] [PubMed]
- Lucca, L.E.; Dominguez-Villar, M. Modulation of regulatory T cell function and stability by co-inhibitory receptors. Nat. Rev. Immunol. 2020, 20, 680–693. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, Z.; Cao, W.; Wu, Q.; Yuan, Y.; Zhang, X. Regulatory T cells in COVID-19. Aging Dis. 2021, 12, 1545–1553. [Google Scholar] [CrossRef] [PubMed]
- Galvan-Pena, S.; Leon, J.; Chowdhary, K.; Michelson, D.A.; Vijaykumar, B.; Yang, L.; Magnuson, A.; Manickas-Hill, Z.; Piechocka-Trocha, A.; Worrall, D.P.; et al. Profound Treg perturbations correlate with COVID-19 severity. bioRxiv 2020. [Google Scholar] [CrossRef] [PubMed]
- Vick, S.C.; Frutoso, M.; Mair, F.; Konecny, A.J.; Greene, E.; Wolf, C.R.; Logue, J.K.; Boonyaratanakornkit, J.; Gottardo, R.; Schiffer, J.T.; et al. A differential regulatory T cell signature distinguishes the immune landscape of COVID-19 hospitalized patients from those hospitalized with other respiratory viral infections. medRxiv 2021. [Google Scholar] [CrossRef]
- Kratzer, B.; Trapin, D.; Ettel, P.; Körmöczi, U.; Rottal, A.; Tuppy, F.; Feichter, M.; Gattinger, P.; Borochova, K.; Dorofeeva, Y.; et al. Immunological imprint of COVID-19 on human peripheral blood leukocyte populations. Allergy 2021, 76, 751–765. [Google Scholar] [CrossRef]
- Liu, J.; Yang, X.; Wang, H.; Li, Z.; Deng, H.; Liu, J.; Xiong, S.; He, J.; Feng, X.; Guo, C.; et al. Analysis of the Long-Term Impact on Cellular Immunity in COVID-19-Recovered Individuals Reveals a Profound NKT Cell Impairment. mBio 2021, 12, e00085-21. [Google Scholar] [CrossRef]
- Rodda, L.B.; Netland, J.; Shehata, L.; Pruner, K.B.; Morawski, P.A.; Thouvenel, C.; Takehara, K.K.; Eggenberger, J.; Hemann, E.; Waterman, H.R.; et al. Functional SARS-CoV-2-specific immune memory persists after mild COVID-19. Res. Sq. 2020; preprint. [Google Scholar] [CrossRef]
- Xiang, T.; Liang, B.; Fang, Y.; Lu, S.; Li, S.; Wang, H.; Li, H.; Yang, X.; Shen, S.; Zhu, B.; et al. Declining Levels of Neutralizing Antibodies against SARS-CoV-2 in Convalescent COVID-19 Patients One Year Post Symptom Onset. Front. Immunol. 2021, 12, 708523. [Google Scholar] [CrossRef]
- Qin, X.; Shen, J.; Dai, E.; Li, H.; Tang, G.; Zhang, L.; Hou, X.; Lu, M.; Wu, X.; Duan, S.; et al. The seroprevalence and kinetics of IgM and IgG in the progression of COVID-19. BMC Immunol. 2021, 22, 14. [Google Scholar] [CrossRef]
- Macedo-Ojeda, G.; Muñoz-Valle, J.F.; Yokogawa-Teraoka, P.; Machado-Sulbarán, A.C.; Loza-Rojas, M.G.; García-Arredondo, A.C.; Tejeda-Constantini, R.; Vega-Magaña, A.N.; González-Estevez, G.; García-Chagollán, M.; et al. COVID-19 Screening by Anti-SARS-CoV-2 Antibody Seropositivity: Clinical and Epidemiological Characteristics, Comorbidities, and Food Intake Quality. Int. J. Environ. Res. Public Health 2021, 18, 8995. [Google Scholar] [CrossRef] [PubMed]
- Dolan, M.E.; Hill, D.P.; Mukherjee, G.; McAndrews, M.S.; Chesler, E.J.; Blake, J.A. Investigation of COVID-19 comorbidities reveals genes and pathways coincident with the SARS-CoV-2 viral disease. Sci. Rep. 2020, 10, 20848. [Google Scholar] [CrossRef] [PubMed]
- Lingel, H.; Meltendorf, S.; Billing, U.; Thurm, C.; Vogel, K.; Majer, C.; Prätsch, F.; Roggenbuck, D.; Heuft, H.G.; Hachenberg, T.; et al. Unique autoantibody prevalence in long-term recovered SARS-CoV-2-infected individuals. J. Autoimmun. 2021, 122, 102682. [Google Scholar] [CrossRef] [PubMed]
- Schub, D.; Klemis, V.; Schneitler, S.; Mihm, J.; Lepper, P.M.; Wilkens, H.; Bals, R.; Eichler, H.; Gärtner, B.C.; Becker, S.L.; et al. High levels of SARS-CoV-2-specific T cells with restricted functionality in severe courses of COVID-19. JCI Insight 2020, 5, e142167. [Google Scholar] [CrossRef]
- Ward, F.J.; Dahal, L.N.; Khanolkar, R.C.; Shankar, S.P.; Barker, R.N. Targeting the alternatively spliced soluble isoform of CTLA-4: Prospects for immunotherapy? Immunotherapy 2014, 6, 1073–1084. [Google Scholar] [CrossRef]
- Davis, I.M. SARS-CoV: Lessons learned; opportunities missed for SARS-CoV-2. Rev. Med. Virol. 2021, 31, 1–6. [Google Scholar] [CrossRef]
- Montalvo Villalba, M.C.; Valdés Ramírez, O.; Muné Jiménez, M.; Arencibia Garcia, A.; Martinez Alfonso, J.; González Baéz, G.; Roque Arrieta, R.; Rosell Simón, D.; Alvárez Gainza, D.; Sierra Vázquez, B.; et al. Interferon gamma, TGF-β1 and RANTES expression in upper airway samples from SARS-CoV-2 infected patients. Clin. Immunol. 2020, 220, 108576. [Google Scholar] [CrossRef]
- Han, H.; Ma, Q.; Li, C.; Liu, R.; Zhao, L.; Wang, W.; Zhang, P.; Liu, X.; Gao, G.; Liu, F.; et al. Profiling serum cytokines in COVID-19 patients reveals IL-6 and IL-10 are disease severity predictors. Emerg. Microbes Infect. 2020, 9, 1123–1130. [Google Scholar] [CrossRef]
- Chen, W. A potential treatment of COVID-19 with TGF-β blockade. Int. J. Biol. Sci. 2020, 16, 1954–1955. [Google Scholar] [CrossRef]
- Agita, A.; Alsagaff, M.T. Inflammation, Immunity, and Hypertension. Acta Med. Indones. 2017, 49, 158–165. [Google Scholar]
- Paust, S.; Cantor, H. Regulatory T cells and autoimmune disease. Immunol. Rev. 2005, 204, 195–207. [Google Scholar] [CrossRef] [PubMed]
- Pan, P.Y.; Ma, G.; Weber, K.J.; Ozao-Choy, J.; Wang, G.; Yin, B.; Divino, C.M.; Chen, S.H. Immune stimulatory receptor CD40 is required for T-cell suppression and T regulatory cell activation mediated by myeloid-derived suppressor cells in cancer. Cancer Res. 2010, 70, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Liu, Y.; Lau, Y.L.; Tu, W. CD40-activated B cells are more potent than immature dendritic cells to induce and expand CD4(+) regulatory T cells. Cell. Mol. Immunol. 2010, 7, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Tu, W.; Lau, Y.L.; Zheng, J.; Liu, Y.; Chan, P.L.; Mao, H.; Dionis, K.; Schneider, P.; Lewis, D.B. Efficient generation of human alloantigen-specific CD4+ regulatory T cells from naive precursors by CD40-activated B cells. Blood 2008, 112, 2554–2562. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.A.; Kogan, S.; Amano, S.U.; Wang, M.; Dagdeviren, S.; Friedline, R.H.; Aouadi, M.; Kim, J.K.; Czech, M.P. CD40 deficiency in mice exacerbates obesity-induced adipose tissue inflammation, hepatic steatosis, and insulin resistance. Am. J. Physiol. Endocrinol. Metab. 2013, 304, E951–E963. [Google Scholar] [CrossRef]
- Cipollone, F.; Chiarelli, F.; Davì, G.; Ferri, C.; Desideri, G.; Fazia, M.; Iezzi, A.; Santilli, F.; Pini, B.; Cuccurullo, C.; et al. Enhanced soluble CD40 ligand contributes to endothelial cell dysfunction in vitro and monocyte activation in patients with diabetes mellitus: Effect of improved metabolic control. Diabetologia 2005, 48, 1216–1224. [Google Scholar] [CrossRef]
- Linna, H.; Suija, K.; Rajala, U.; Herzig, K.H.; Karhu, T.; Jokelainen, J.; Keinänen-Kiukaanniemi, S.; Timonen, M. The association between impaired glucose tolerance and soluble CD40 ligand: A 15-year prospective cohort study. Aging Clin. Exp. Res. 2016, 28, 1243–1249. [Google Scholar] [CrossRef]
- Wong, C.K.; Ho, A.W.; Tong, P.C.; Yeung, C.Y.; Chan, J.C.; Kong, A.P.; Lam, C.W. Aberrant expression of soluble co-stimulatory molecules and adhesion molecules in type 2 diabetic patients with nephropathy. J. Clin. Immunol. 2008, 28, 36–43. [Google Scholar] [CrossRef]
- Sonmez, A.; Dogru, T.; Yilmaz, M.I.; Ocal, R.; Ozgurtas, T.; Kilic, S.; Eyileten, T.; Tasci, I.; Erbil, K.; Kocar, I.H. Soluble CD40 ligand levels in patients with hypertension. Clin. Exp. Hypertens. 2005, 27, 629–634. [Google Scholar] [CrossRef]
- Lamine, L.B.; Turki, A.; Al-Khateeb, G.; Sellami, N.; Amor, H.B.; Sarray, S.; Jailani, M.; Ghorbel, M.; Mahjoub, T.; Almawi, W.Y. Elevation in Circulating Soluble CD40 Ligand Concentrations in Type 2 Diabetic Retinopathy and Association with its Severity. Exp. Clin. Endocrinol. Diabetes 2020, 128, 319–324. [Google Scholar] [CrossRef]
- Yngen, M.; Östenson, C.G.; Hu, H.; Li, N.; Hjemdahl, P.; Wallén, N.H. Enhanced P-selectin expression and increased soluble CD40 Ligand in patients with Type 1 diabetes mellitus and microangiopathy: Evidence for platelet hyperactivity and chronic inflammation. Diabetologia 2004, 47, 537–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Uden, D.; Koudstaal, T.; van Hulst, J.A.C.; van den Bosch, T.P.P.; Vink, M.; Bergen, I.M.; Lila, K.A.; van den Bosch, A.E.; Bresser, P.; Kool, M.; et al. Evidence for a Role of CCR6+ T Cells in Chronic Thromboembolic Pulmonary Hypertension. Front. Immunol. 2022, 13, 861450. [Google Scholar] [CrossRef] [PubMed]
- Wolf, D.; Jehle, F.; Michel, N.A.; Bukosza, E.N.; Rivera, J.; Chen, Y.C.; Hoppe, N.; Dufner, B.; Rodriguez, A.O.; Colberg, C.; et al. Coinhibitory suppression of T cell activation by CD40 protects against obesity and adipose tissue inflammation in mice. Circulation 2014, 129, 2414–2425. [Google Scholar] [CrossRef] [PubMed]
- Guilherme, A.; Virbasius, J.V.; Puri, V.; Czech, M.P. Adipocyte dysfunctions linking obesity to insulin resistance and type 2 diabetes. Nat. Rev. Mol. Cell Biol. 2008, 9, 367–377. [Google Scholar] [CrossRef]
- Van Gaal, L.F.; Mertens, I.L.; De Block, C.E. Mechanisms linking obesity with cardiovascular disease. Nature 2006, 444, 875–880. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics of Subjects | Healthy Donors (HD) n = 20 | Non-Comorbid (NC) n = 40 | Comorbid (C) n = 18 | Reference Range |
|---|---|---|---|---|
| Gender (F/M) | 13/7 | 23/17 | 10/8 | |
| Age (y) | 37.40 ± 9.87 $ | 37.08 ± 10.43 $ | 51.50 ± 10.63 $,*,+ | |
| Comorbid (n) | ||||
| Diabetes mellitus (DM) | - | - | 8 | |
| Hypertension (HTN) | - | - | 10 | |
| Anthropometric Data | ||||
| Height (m) | 1.60 + 0.38 # | 1.64 + 0.39 # | 1.66 + 0.38 # | |
| Weight (kg) | 62.25 + 52.30 # | 73.05 + 73.20 # | 84.80 + 83.62 #,* | |
| BMI (kg/m2) | 24.98 + 24.67 # | 26.56 + 21.70 # | 30.01 + 22.02 #,* | |
| WC (cm) | 80.50 + 39.0 # | 92.0 + 74.0 # | 97.0 + 41.0 #,* | |
| WHR | 0.84 ± 0.09 $ | 0.87 ± 0.09 $ | 0.91 ± 0.07 $,* | |
| SBP (mmHg) | 117.0 + 22.0 # | 98.50 + 104.0 #,* | 130.0 + 50.0 #,+ | |
| Antibody Test Results | ||||
| IgG-N Cut-of index | 0.066 + 0.12 # | 5.29 + 153.4 # | 62.81 + 151.5 #,*,+ | <1.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Méndez-Frausto, G.; Godina-González, S.; Rivas-Santiago, C.E.; Nungaray-Anguiano, E.; Mendoza-Almanza, G.; Rivas-Santiago, B.; Galván-Tejada, C.E.; Gonzalez-Curiel, I.E. Downregulation of sCD40 and sCTLA4 in Recovered COVID-19 Patients with Comorbidities. Pathogens 2022, 11, 1128. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11101128
Méndez-Frausto G, Godina-González S, Rivas-Santiago CE, Nungaray-Anguiano E, Mendoza-Almanza G, Rivas-Santiago B, Galván-Tejada CE, Gonzalez-Curiel IE. Downregulation of sCD40 and sCTLA4 in Recovered COVID-19 Patients with Comorbidities. Pathogens. 2022; 11(10):1128. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11101128
Chicago/Turabian StyleMéndez-Frausto, Gwendolyne, Susana Godina-González, César E. Rivas-Santiago, Edna Nungaray-Anguiano, Gretel Mendoza-Almanza, Bruno Rivas-Santiago, Carlos E. Galván-Tejada, and Irma E. Gonzalez-Curiel. 2022. "Downregulation of sCD40 and sCTLA4 in Recovered COVID-19 Patients with Comorbidities" Pathogens 11, no. 10: 1128. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11101128