
Liver Proteome Alterations in Red Deer (Cervus elaphus) Infected by the Giant Liver Fluke Fascioloides magna
 ,
,
Abstract
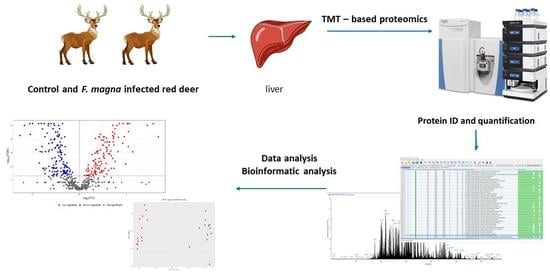
:
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Sample Preparation
2.3. LC-MS/MS Analysis
2.4. Statistical and Bioinformatic Analysis
3. Results
3.1. F. magna Infection Diagnosis and Liver Inspection
3.2. Proteomics Analysis
3.3. Functional Enrichment Analysis
4. Discussion
4.1. Alterations in Translation Processes during F. magna Infection
4.2. Alterations in Liver Metabolism during F. magna Infection
4.3. Host-Parasite Interface during F. magna Infection
4.4. Fibrosis-Related Protein Alterations during F. magna Infection
4.5. Oxidative Stress and Drug Metabolism Changes during F. magna Infection
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fürst, T.; Keiser, J.; Utzinger, J. Global burden of human food-borne trematodiasis: A systematic review and meta-analysis. Lancet Infect. Dis. 2012, 12, 210–221. [Google Scholar] [CrossRef] [PubMed]
- Petney, T.N.; Sithithaworn, P.; Andrews, R.H.; Webster, J.P. Foodborne trematodes: A diverse and challenging group of neglected parasites. Trans. R. Soc. Trop. Med. Hyg. 2016, 110, 1–3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vale, N.; Gouveia, M.J.; Gärtner, F.; Brindley, P.J. Oxysterols of helminth parasites and pathogenesis of foodborne hepatic trematodiasis caused by Opisthorchis and Fasciola species. Parasitol. Res. 2020, 119, 1443–1453. [Google Scholar] [CrossRef] [PubMed]
- Králová-Hromadová, I.; Juhásová, Ľ.; Bazsalovicsová, E. The Giant Liver Fluke, Fascioloides magna: Past, Present and Future Research; Springer International Publishing: Heidelberg, Germany, 2016; p. 106. [Google Scholar]
- Pybus, M.J. Liver Flukes. In Parasitic Diseases of Wild Mammals, 2nd ed.; Samuel, W.M., Pybus, M.J., Kocan, A.A., Eds.; Wiley Online Library: Hoboken, NJ, USA, 2001; pp. 121–149. [Google Scholar]
- Daszak, P.; Cunningham, A.A.; Hyatt, A.D. Emerging infectious diseases of wildlife--threats to biodiversity and human health. Science 2000, 287, 443–449. [Google Scholar] [CrossRef]
- Vilà, M.; Hulme, P.E. Non-native Species, Ecosystem Services, and Human Well-Being. In Impact of Biological Invasions on Ecosystem Services; Vilà, M., Hulme, P.E., Eds.; Springer International: Cham, Switzerland, 2017; pp. 1–14. [Google Scholar]
- Foreyt, W.J.; Todd, A.C. Effects of six fasciolicides against Fascioloides magna In white-tailed deer. J. Wildl. Dis. 1976, 12, 361–366. [Google Scholar] [CrossRef]
- Marinculić, A.; Džakula, N.; Janicki, Z.; Hardy, Z.; Lučinger, S.; Živičnjak, T. Appearance of American liver fluke (Fascioloides magna, Bassi, 1875) in Croatia—A case report. Vet. Arhiv. 2002, 72, 319–325. [Google Scholar]
- Slavica, A.; Florijančić, T.; Janicki, Z.; Konjević, D.; Severin, K.; Marinculić, A.; Pintur, K. Treatment of fascioloidosis (Fascioloides magna, Bassi, 1875) in free ranging and captive red deer (Cervus elaphus L.) at eastern Croatia. Vet. Arhiv. Suppl. 2006, 76, 9–19. [Google Scholar]
- Konjević, D.; Janicki, Z.; Calmels, P.; Jan, D.S.; Marinculić, A.; Šimunović, M.; Pavlak, M.; Krapinec, K.; Poljak, Z. Evaluation of factors affecting the efficacy of treatment against Fascioloides magna in wild red deer population. Vet. Ital. 2018, 54, 33–39. [Google Scholar]
- Šimunović, M. Influence of Epizootiologic Factors on Control of Deer Fascioloidosis in Baranja. Ph.D. Thesis, University of Zagreb, Veterinary Faculty, Zagreb, Croatia, 2017. [Google Scholar]
- Bennett, A.P.S.; Robinson, M.W. Trematode Proteomics: Recent Advances and Future Directions. Pathogens 2021, 10, 348. [Google Scholar] [CrossRef]
- McManus, D.P. Recent progress in the development of liver fluke and blood fluke vaccines. Vaccines 2020, 8, 553. [Google Scholar] [CrossRef]
- Molina-Hernández, V.; Mulcahy, G.; Pérez, J.; Martínez-Moreno, Á.; Donnelly, S.; O’Neill, S.M.; Dalton, J.P.; Cwiklinski, K. Fasciola hepatica vaccine: We may not be there yet but we’re on the right road. Vet. Parasitol. 2015, 208, 101–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silvane, L.; Celias, D.P.; Romagnoli, P.A.; Maletto, B.A.; Sanchez Vallecillo, M.F.; Chiapello, L.S.; Palma, S.D.; Allemandi, D.A.; Sanabria, R.E.F.; Pruzzo, C.I.; et al. A Vaccine Based on Kunitz-Type Molecule Confers Protection Against Fasciola hepatica Challenge by Inducing IFN-γ and Antibody Immune Responses Through IL-17A Production. Front. Immunol. 2020, 11, 2087. [Google Scholar] [CrossRef]
- Cantacessi, C.; Mulvenna, J.; Young, N.D.; Kasny, M.; Horak, P.; Aziz, A.; Hofmann, A.; Loukas, A.; Gasser, R.B. A deep exploration of the transcriptome and “excretory/secretory” proteome of adult Fascioloides magna. Mol. Cell. Proteom. 2012, 11, 1340–1353. [Google Scholar] [CrossRef] [PubMed]
- Kuleš, J.; Lovrić, L.; Gelemanović, A.; Beer Ljubić, B.; Rubić, I.; Bujanić, M.; Konjević, D. Complementary liver and serum protein profile in wild boars infected by the giant liver fluke Fascioloides magna using tandem mass tags quantitative approach. J. Proteom. 2021, 247, 104332. [Google Scholar] [CrossRef] [PubMed]
- Cwiklinski, K.; Robinson, M.W.; Donnelly, S.; Dalton, J.P. Complementary transcriptomic and proteomic analyses reveal the cellular and molecular processes that drive growth and development of Fasciola hepatica in the host liver. BMC Genom. 2021, 22, 46. [Google Scholar] [CrossRef] [PubMed]
- Haçarız, O.; Sayers, G.; Baykal, A.T. A proteomic approach to investigate the distribution and abundance of surface and internal Fasciola hepatica proteins during the chronic stage of natural liver fluke infection in cattle. J. Proteome Res. 2012, 11, 3592–3604. [Google Scholar] [CrossRef]
- Robinson, M.W.; Menon, R.; Donnelly, S.M.; Dalton, J.P.; Ranganathan, S. An integrated transcriptomics and proteomics analysis of the secretome of the helminth pathogen Fasciola hepatica: Proteins associated with invasion and infection of the mammalian host. Mol. Cell. Proteom. 2009, 8, 1891–1907. [Google Scholar] [CrossRef] [Green Version]
- Di Maggio, L.S.; Tirloni, L.; Pinto, A.F.M.; Diedrich, J.K.; Yates III, J.R.; Benavides, U.; Carmona, C.; Da Silva Vaz, I., Jr.; Berasain, P. Across intra-mammalian stages of the liver f luke Fasciola hepatica: A proteomic study. Sci. Rep. 2016, 6, 32796. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.K.; Hu, R.S.; Elsheikha, H.M.; Sheng, Z.A.; Zhang, W.Y.; Zheng, W.B.; Zhu, X.Q.; He, J.J. Global serum proteomic changes in water buffaloes infected with Fasciola gigantica. Parasites Vectors 2019, 12, 281. [Google Scholar] [CrossRef] [Green Version]
- Wagenknecht, E. Altersbestimmung des Erlegten Wildes; Neumann-Neudamm GmbH & Co. Kg: Melsungen, Germany, 1984. [Google Scholar]
- Mazeri, S.; Sargison, N.; Kelly, R.F.; Bronsvoort, B.M.D.; Handel, I. Evaluation of the Performance of Five Diagnostic Tests for Fasciola hepatica Infection in Naturally Infected Cattle Using a Bayesian No Gold Standard Approach. PLoS ONE 2016, 11, e0161621. [Google Scholar] [CrossRef] [Green Version]
- Cornell Wildlife Health Lab. Fascioloides magna. 2018. Available online: https://cwhl.vet.cornell.edu/disease/fascioloides-magna#collapse11 (accessed on 4 March 2022).
- Wiśniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods 2009, 6, 359. [Google Scholar] [CrossRef] [PubMed]
- Perez-Riverol, Y.; Bai, J.; Bandla, C.; García-Seisdedos, D.; Hewapathirana, S.; Kamatchinathan, S.; Kundu, D.J.; Prakash, A.; Frericks-Zipper, A.; Eisenacher, M.; et al. The PRIDE database resources in 2022: A hub for mass spectrometry-based proteomics evidences. Nucleic Acids Res. 2022, 50, D543–D552. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing 2020; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- The UniProt Consortium. UniProt: The universal protein knowledgebase in 2021. Nucleic Acids Res. 2020, 49, D480–D489. [Google Scholar]
- Mi, H.; Muruganujan, A.; Ebert, D.; Huang, X.; Thomas, P.D. PANTHER version 14: More genomes, a new PANTHER GO-slim and improvements in enrichment analysis tools. Nucleic Acids Res. 2018, 47, D419–D426. [Google Scholar] [CrossRef]
- Gillespie, M.; Jassal, B.; Stephan, R.; Milacic, M.; Rothfels, K.; Senff-Ribeiro, A.; Griss, J.; Sevilla, C.; Matthews, L.; Gong, C.; et al. The reactome pathway knowledgebase 2022. Nucleic Acids Res. 2021, 50, D687–D692. [Google Scholar] [CrossRef] [PubMed]
- Deslyper, G.; Doherty, D.G.; Carolan, J.C.; Holland, C.V. The role of the liver in the migration of parasites of global significance. Parasites Vectors 2019, 12, 531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNeilly, T.N.; Nisbet, A.J. Immune modulation by helminth parasites of ruminants: Implications for vaccine development and host immune competence. Parasite 2014, 21, 51. [Google Scholar] [CrossRef] [Green Version]
- Bhavsar, R.B.; Makley, L.N.; Tsonis, P.A. The other lives of ribosomal proteins. Hum. Genom. 2010, 4, 327. [Google Scholar] [CrossRef]
- Yen, H.C.; Xu, Q.; Chou, D.M.; Zhao, Z.; Elledge, S.J. Global protein stability profiling in mammalian cells. Science 2008, 322, 918–923. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, A.L.; Ciechanover, A. The ubiquitin-proteasome pathway and pathogenesis of human diseases. Annu. Rev. Med. 1999, 50, 57–74. [Google Scholar] [CrossRef]
- Benanti, J.A. Coordination of cell growth and division by the ubiquitin-proteasome system. Semin. Cell Dev. Biol. 2012, 23, 492–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Figueroa-Santiago, O.; Ana, M.E. Fasciola hepatica Fatty Acid Binding Protein Induces the Alternative Activation of Human Macrophages. Infect. Immun. 2014, 82, 5005–5012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tielens, A. Metabolism in Fasciolosis; Dalton, J., Ed.; CABI Publishing: Wallingford, UK, 1999; pp. 277–305. [Google Scholar]
- Huss, J.M.; Kelly, D.P. Nuclear receptor signaling and cardiac energetics. Circ. Res. 2004, 95, 568–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cwiklinski, K.; Jewhurst, H.; Mcveigh, P.; Barbour, T.; Maule, A.G.; Tort, J.; O’Neill, S.M.; Robinson, M.W.; Donnelly, S.; Dalton, J.P. Infection by the Helminth Parasite Fasciola hepatica Requires Rapid Regulation of Metabolic, Virulence, and Invasive Factors to Adjust to Its Mammalian Host. Mol. Cell. Proteom. 2018, 17, 792–809. [Google Scholar] [CrossRef] [PubMed]
- Berchtold, M.W.; Villalobo, A. The many faces of calmodulin in cell proliferation, programmed cell death, autophagy, and cancer. Biochim. Biophys. Acta (BBA) Mol. Cell Res. 2014, 1843, 398–435. [Google Scholar] [CrossRef]
- Rescher, U.; Gerke, V. Annexins—Unique membrane binding proteins with diverse functions. J. Cell Sci. 2004, 117 Pt 13, 2631–2639. [Google Scholar] [CrossRef] [Green Version]
- Gerke, V.; Creutz, C.E.; Moss, S.E. Annexins: Linking Ca2+ signalling to membrane dynamics. Nat. Rev. Mol. Cell Biol. 2005, 6, 449–461. [Google Scholar] [CrossRef]
- Hofmann, A.; Osman, A.; Leow, C.Y.; Driguez, P.; Mcmanus, D.P.; Jones, M.K. Parasite annexins—New molecules with potential for drug and vaccine development. BioEssays 2010, 32, 967–976. [Google Scholar] [CrossRef] [Green Version]
- Osterloh, A.; Breloer, M. Heat shock proteins: Linking danger and pathogen recognition. Med. Microbiol. Immunol. 2008, 197, 1–8. [Google Scholar] [CrossRef]
- Abaza, S. Heat shock proteins and parasitic diseases: Part 1: Helminths. Parasitol. United J. 2014, 7, 93–103. [Google Scholar] [CrossRef]
- Colaco, C.A.; Bailey, C.R.; Walker, K.B.; Keeble, J. Heat shock proteins: Stimulators of innate and acquired immunity. BioMed Res. Int. 2013, 2013, 461230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNulty, S.; Colaco, C.A.; Blandford, L.E.; Bailey, C.R.; Baschieri, S.; Todryk, S. Heat-shock proteins as dendritic cell-targeting vaccines—Getting warmer. Immunology 2013, 139, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Fagone, P.; Mangano, K.; Mammana, S.; Pesce, A.; Pesce, A.; Caltabiano, R.; Giorlandino, A.; Portale, T.R.; Cavalli, E.; Lombardo, G.a.G.; et al. Identification of novel targets for the diagnosis and treatment of liver fibrosis. Int. J. Mol. Med. 2015, 36, 747–752. [Google Scholar] [CrossRef] [Green Version]
- Delanote, V.; Vandekerckhove, J.; Gettemans, J. Plastins: Versatile modulators of actin organization in (patho)physiological cellular processes. Acta Pharmacol. Sin. 2005, 26, 769–779. [Google Scholar] [CrossRef]
- Janji, B.; Giganti, A.; De Corte, V.; Catillon, M.; Bruyneel, E.; Lentz, D.; Plastino, J.; Gettemans, J.; Friederich, E. Phosphorylation on Ser5 increases the F-actin-binding activity of L-plastin and promotes its targeting to sites of actin assembly in cells. J. Cell Sci. 2006, 119 Pt 9, 1947–1960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wabnitz, G.H.; Köcher, T.; Lohneis, P.; Stober, C.; Konstandin, M.H.; Funk, B.; Sester, U.; Wilm, M.; Klemke, M.; Samstag, Y. Costimulation induced phosphorylation of L-plastin facilitates surface transport of the T cell activation molecules CD69 and CD25. Eur. J. Immunol. 2007, 37, 649–662. [Google Scholar] [CrossRef]
- Honda, K. The biological role of actinin-4 (ACTN4) in malignant phenotypes of cancer. Cell Biosci. 2015, 5, 41. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Hsu, K.S.; Lim, J.H.; Bruggeman, L.A.; Kao, H.Y. α-Actinin 4 potentiates nuclear factor κ-light-chain-enhancer of activated B-cell (NF-κB) activity in podocytes independent of its cytoplasmic actin binding function. J. Biol. Chem. 2015, 290, 338–349. [Google Scholar] [CrossRef] [Green Version]
- Liem, R.K.H. Cytoskeletal Integrators: The Spectrin Superfamily. Cold Spring Harb. Perspect. Biol. 2016, 8, a018259. [Google Scholar] [CrossRef] [Green Version]
- Yang, P.; Yang, Y.; Sun, P.; Tian, Y.; Gao, F.; Wang, C.; Zong, T.; Li, M.; Zhang, Y.; Yu, T.; et al. βII spectrin (SPTBN1): Biological function and clinical potential in cancer and other diseases. Int. J. Biol. Sci. 2021, 17, 32–49. [Google Scholar] [CrossRef]
- Nassal, D.M.; Patel, N.J.; Unudurthi, S.D.; Shaheen, R.; Yu, J.; Mohler, P.J.; Hund, T.J. Ca2+/calmodulin kinase II–dependent regulation of βIV-spectrin modulates cardiac fibroblast gene expression, proliferation, and contractility. J. Biol. Chem. 2021, 297, 100893. [Google Scholar] [CrossRef] [PubMed]
- Wittmann, T.; Bokoch, G.M.; Waterman-Storer, C.M. Regulation of leading edge microtubule and actin dynamics downstream of Rac1. J. Cell Biol. 2003, 161, 845–851. [Google Scholar] [CrossRef] [PubMed]
- Byrne, K.M.; Monsefi, N.; Dawson, J.C.; Degasperi, A.; Bukowski-Wills, J.-C.; Volinsky, N.; Dobrzyński, M.; Birtwistle, M.R.; Tsyganov, M.; Kiyatkin, A.; et al. Bistability in the Rac1, PAK, and RhoA Signaling Network Drives Actin Cytoskeleton Dynamics and Cell Motility Switches. Cell Syst. 2016, 2, 38–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bottari, N.B.; Mendes, R.E.; Lucca, N.J.; Schwertz, C.I.; Henker, L.C.; Olsson, D.C.; Piva, M.M.; Sangoi, M.; Campos, L.P.; Moresco, R.N.; et al. Oxidative stress associated with pathological lesions in the liver of rats experimentally infected by Fasciola hepatica. Exp. Parasitol. 2015, 159, 24–28. [Google Scholar] [CrossRef]
- Becker, K.; Tilley, L.; Vennerstrom, J.L.; Roberts, D.; Rogerson, S.; Ginsburg, H. Oxidative stress in malaria parasite-infected erythrocytes: Host–parasite interactions. Int. J. Parasitol. 2004, 34, 163–189. [Google Scholar] [CrossRef]
- Kaya, S.; Sütçü, R.; Cetin, E.S.; Aridogan, B.C.; Delibaş, N.; Demirci, M. Lipid peroxidation level and antioxidant enzyme activities in the blood of patients with acute and chronic fascioliasis. Int. J. Infect. Dis. 2007, 11, 251–255. [Google Scholar] [CrossRef] [Green Version]
- Kolodziejczyk, L.; Siemieniuk, E.; Skrzydlewska, E. Antioxidant potential of rat liver in experimental infection with Fasciola hepatica. Parasitol. Res. 2005, 96, 367–372. [Google Scholar] [CrossRef]
- Kolodziejczyk, L.; Siemieniuk, E.; Skrzydlewska, E. Fasciola hepatica: Effects on the antioxidative properties and lipid peroxidation of rat serum. Exp. Parasitol. 2006, 113, 43–48. [Google Scholar] [CrossRef]
- Devine, C.; Brennan, G.P.; Lanusse, C.E.; Alvarez, L.I.; Trudgett, A.; Hoey, E.; Fairweather, I. Inhibition of cytochrome P450-mediated metabolism enhances ex vivo susceptibility of Fasciola hepatica to triclabendazole. Parasitology 2010, 137, 871–880. [Google Scholar] [CrossRef]
- Devine, C.; Brennan, G.P.; Lanusse, C.E.; Alvarez, L.I.; Trudgett, A.; Hoey, E.; Fairweather, I. Inhibition of triclabendazole metabolism in vitro by ketoconazole increases disruption to the tegument of a triclabendazole-resistant isolate of Fasciola hepatica. Parasitol. Res. 2011, 109, 981–995. [Google Scholar] [CrossRef]
- Devine, C.; Brennan, G.P.; Lanusse, C.E.; Alvarez, L.I.; Trudgett, A.; Hoey, E.; Fairweather, I. Potentiation of triclabendazole action in vivo against a triclabendazole-resistant isolate of Fasciola hepatica following its co-administration with the metabolic inhibitor, ketoconazole. Vet. Parasitol. 2012, 184, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Alvarez Rojas, C.A.; Ansell, B.R.E.; Hall, R.S.; Gasser, R.B.; Young, N.D.; Jex, A.R.; Scheerlinck, J.-P.Y. Transcriptional analysis identifies key genes involved in metabolism, fibrosis/tissue repair and the immune response against Fasciola hepatica in sheep liver. Parasites Vectors 2015, 8, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarez, L.I.; Solana, H.D.; Mottier, M.L.; Virkel, G.L.; Fairweather, I.; Lanusse, C.E. Altered drug influx/efflux and enhanced metabolic activity in triclabendazole-resistant liver flukes. Parasitology 2005, 131 Pt 4, 501–510. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Migration | Juvenile Fluke | Adult Fluke | Pseudocyst | Destroyed Pseudocyst | |
|---|---|---|---|---|---|
| Sample 1 | 0 | 0 | 2 | 1 | 0 |
| Sample 2 | 1 | 0 | 2 | 1 | 1 |
| Sample 3 | 1 | 1 | 2 | 1 | 0 |
| Sample 4 | 3 | 1 | 8 | 8 | 7 |
| Sample 5 | 2 | 12 | 1 | 3 | 65 |
| Sample 6 | 0 | 5 | 12 | 5 | 0 |
| Sample 7 | 3 | 21 | 25 | 6 | 0 |
| Sample 8 | 1 | 1 | 0 | 0 | 78 |
| Sample 9 | 6 | 6 | 4 | 11 | 11 |
| Sample 10 | 5 | 23 | 59 | 8 | 0 |
| Sample 11 | 0 | 1 | 2 | 1 | 1 |
| Sample 12 | 1 | 5 | 44 | 22 | 5 |
| Sample 13 | 7 | 3 | 0 | 14 | 7 |
| Accession | Gene ID | Description | p Value | FDR | FC | log2FC |
|---|---|---|---|---|---|---|
| Proteins with higher abundance in F. magna group | ||||||
| A0A6J0WZS6 | SLC27A2 | Very long-chain acyl-CoA synthetase-like | <0.001 | <0.001 | 2.77 | 1.47 |
| A0A6J0W5Y4 | CYP3A4 | Cytochrome P450 3A24 | <0.001 | <0.001 | 2.08 | 1.06 |
| A0A6J0ZFP6 | CYP4A11 | Taurochenodeoxycholic 6 alpha-hydroxylase-like isoform X2 | <0.001 | <0.001 | 2.08 | 1.06 |
| A0A5N3V0H2 | UGT2B4 | UDP-glucuronosyltransferase | <0.001 | 0.001 | 1.94 | 0.96 |
| A0A6J0YYJ9 | ACOT2 | Acyl-coenzyme A thioesterase 1-like isoform X1 | <0.001 | 0.001 | 1.84 | 0.88 |
| A0A6J0XYD9 | PCCA | Propanoyl-CoA:carbon dioxide ligase subunit alpha | <0.001 | 0.001 | 1.84 | 0.88 |
| A0A5N3XTI4 | HNRNPU | Heterogeneous nuclear ribonucleoprotein U (Scaffold attachment factor A) ** | 0.009 | 0.014 | 1.77 | 0.82 |
| A0A5J5MHR2 | TUBA1D | Tubulin alpha chain | <0.001 | <0.001 | 1.73 | 0.79 |
| A0A212DC96 | SULT2A1 | Sulfotransferase (Fragment) | 0.013 | 0.019 | 1.70 | 0.76 |
| A0A212DAL6 | FASN | 3-hydroxyacyl-[acyl-carrier-protein] dehydratase | 0.029 | 0.035 | 1.67 | 0.74 |
| Proteins with lower abundance in F. magna group | ||||||
| A0A6J0WX68 | H3C1 | Histone H3.1 ** | <0.001 | <0.001 | 0.50 | −0.99 |
| A0A5N3V8G4 | UQCRB | Cytochrome b-c1 complex subunit 7 | <0.001 | <0.001 | 0.50 | −1.00 |
| A0A6J0XG22 | UBC | Polyubiquitin-C | <0.001 | <0.001 | 0.48 | −1.05 |
| A0A6J0Z5P0 | RPL35A | 60S ribosomal protein L35a | <0.001 | <0.001 | 0.48 | −1.05 |
| A0A6J0WHA7 | TST | Sulfurtransferase | <0.001 | <0.001 | 0.47 | −1.10 |
| A0A6J0X0S3 | HSD17B14 | 17-beta-hydroxysteroid dehydrogenase 14 | 0.004 | 0.007 | 0.46 | −1.13 |
| A0A5N3WJW3 | CALM | Calmodulin ** | <0.001 | <0.001 | 0.46 | −1.13 |
| A0A5N3W733 | FTH1 | Ferritin | <0.001 | <0.001 | 0.35 | −1.51 |
| O46412 | SOD1 | Superoxide dismutase [Cu-Zn] | <0.001 | <0.001 | 0.35 | −1.52 |
| A0A6J0YAN6 | H2BC15 | Histone H2B | <0.001 | <0.001 | 0.29 | −1.77 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Šimonji, K.; Konjević, D.; Bujanić, M.; Rubić, I.; Farkaš, V.; Beletić, A.; Grbavac, L.; Kuleš, J. Liver Proteome Alterations in Red Deer (Cervus elaphus) Infected by the Giant Liver Fluke Fascioloides magna. Pathogens 2022, 11, 1503. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11121503
Šimonji K, Konjević D, Bujanić M, Rubić I, Farkaš V, Beletić A, Grbavac L, Kuleš J. Liver Proteome Alterations in Red Deer (Cervus elaphus) Infected by the Giant Liver Fluke Fascioloides magna. Pathogens. 2022; 11(12):1503. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11121503
Chicago/Turabian StyleŠimonji, Karol, Dean Konjević, Miljenko Bujanić, Ivana Rubić, Vladimir Farkaš, Anđelo Beletić, Lea Grbavac, and Josipa Kuleš. 2022. "Liver Proteome Alterations in Red Deer (Cervus elaphus) Infected by the Giant Liver Fluke Fascioloides magna" Pathogens 11, no. 12: 1503. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11121503