
The Role of Serratomolide-like Amino Lipids Produced by Bacteria of Genus Serratia in Nematicidal Activity




Abstract
:1. Introduction
2. Results
2.1. Amplification, Purification and Sequencing of swrW and swrA Genes
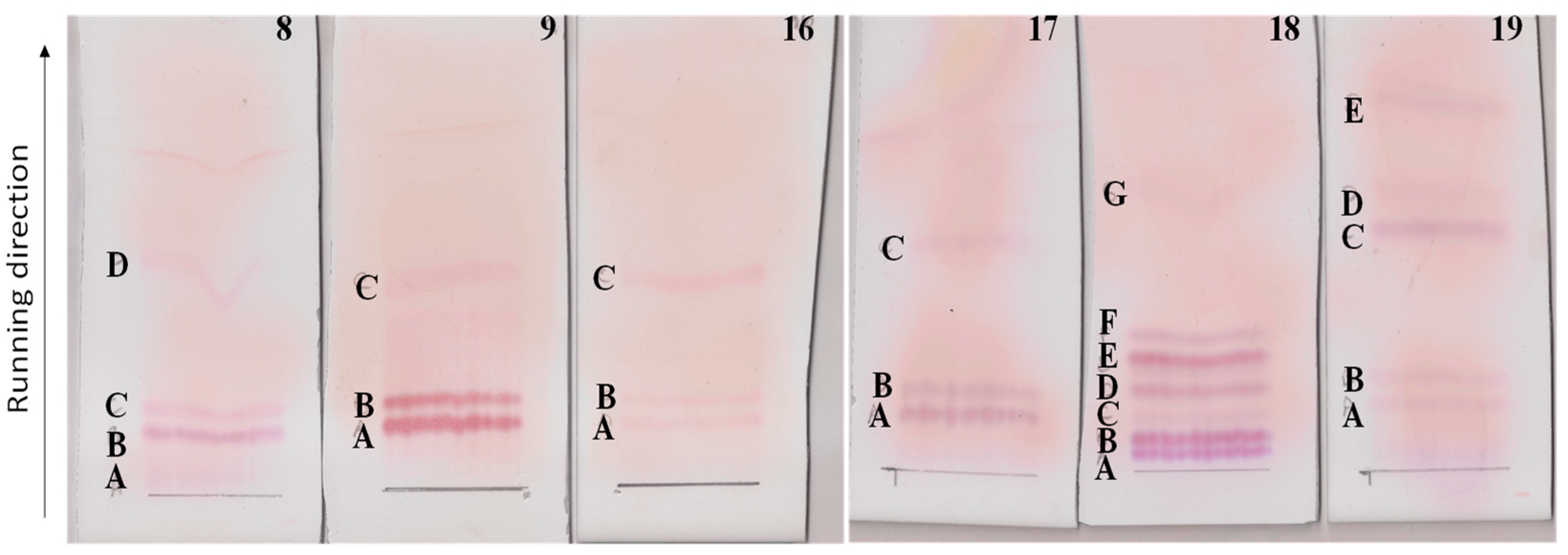
2.2. Lipopeptides Extraction and Purification
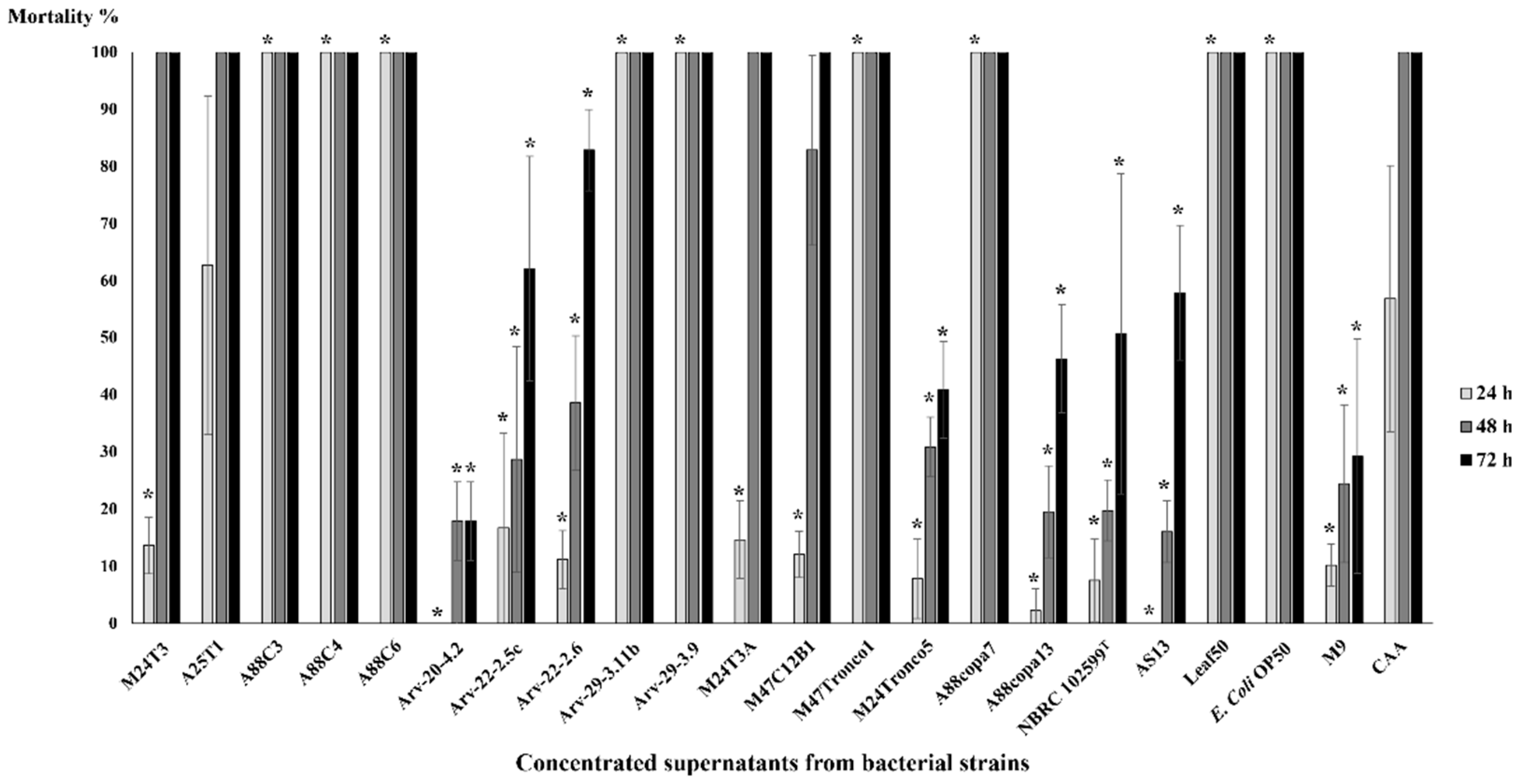
2.3. Mortality Tests in C. elegans
2.4. Mortality Tests in B. xylophilus
2.5. Attraction Tests in C. elegans
2.6. HPLC Analysis of Selected TLC Bands
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains, Media and Growth Conditions
4.2. DNA Extraction and Quantification
4.3. Primers Design, Polymerase Chain Reaction and DNA Sequence Analysis
4.4. Lipopeptides Extraction and Purification
4.5. Maintenance of C. elegans
4.6. Attraction Tests in C. elegans
4.7. Mortality Tests in C. elegans and B. xylophilus
4.8. Statistical Analysis
4.9. Nucleotide Sequence Accession Numbers
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhao, B.G.; Futai, K.; Sutherland, J.R.; Takeuchi, Y. (Eds.) Pine Wilt Disease; Springer: Tokyo, Japan, 2008; ISBN 978-4-431-75654-5. [Google Scholar]
- Jones, J.T.; Moens, M.; Mota, M.; Li, H.; Kikuchi, T. Bursaphelenchus xylophilus: Opportunities in comparative genomics and molecular host-parasite interactions. Mol. Plant Pathol. 2008, 9, 357–368. [Google Scholar] [CrossRef] [PubMed]
- Futai, K. Pine wilt in Japan: From First Incidence to the Present. 2008, pp. 5–12. Available online: https://0-link-springer-com.brum.beds.ac.uk/chapter/10.1007/978-4-431-75655-2_2 (accessed on 15 December 2021).
- Khan, F.A.; Gbadegesin, R.A. On the occurrence of nematode-induced pine wilt disease in Nigeria. Pakistan J. Nematol. 1991, 9, 57–58. [Google Scholar]
- Dwinnel, L.D. First report of pinewood nematode (Bursaphelenchus xylophilus) in Mexico. Plant Dis. 1993, 77, 846. [Google Scholar] [CrossRef]
- Mota, M.; Braasch, H.; Bravo, M.A.; Penas, A.C.; Burgermeister, W.; Metge, K.; Sousa, E. First report of Bursaphelenchus xylophilus in Portugal and in Europe. Nematology 1999, 1, 727–734. [Google Scholar] [CrossRef]
- Sousa, E.; Bonifácio, L.; Pires, J.; Penas, A.C.; Mota, M.; Bravo, M.A.; Naves, P. Bursaphelenchus xylophilus (Nematoda; Aphelenchoididae) associated with Monochamus galloprovincialis (Coleoptera; Cerambycidae) in Portugal. Nematology 2001, 3, 89–91. [Google Scholar] [CrossRef]
- Rodrigues, J.M.; Sousa, E.; Abrantes, I. Pine Wilt Disease historical overview. In Pine Wilt Disease in Europe: Biological Interactions and Integrated Management; Sousa, E., Vale, F., Abrantes, I., Eds.; FNAPF—Federação Nacional das Associações de Proprietários Florestais: Lisbon, Portugal, 2015; pp. 12–32. [Google Scholar]
- Fonseca, L.; Cardoso, J.M.S.; Lopes, A.; Pestana, M.; Abreu, F.; Nunes, N.; Mota, M.; Abrantes, I. The pinewood nematode, Bursaphelenchus xylophilus, in Madeira island. Helminthologia 2012, 49, 96–103. [Google Scholar] [CrossRef] [Green Version]
- Robertson, L.; Cobacho Arcos, S.; Escuer, M.; Santiago Merino, R.; Esparrago, G.; Abelleira, A.; Navas, A. Incidence of the pinewood nematode Bursaphelenchus xylophlius Steiner & Buhrer, 1934 (Nickle, 1970) in Spain. Nematology 2011, 13, 755–757. [Google Scholar] [CrossRef]
- de la Fuente, B.; Saura, S. Long-term projections of the natural expansion of the pine wood nematode in the Iberian Peninsula. Forests 2021, 12, 849. [Google Scholar] [CrossRef]
- Kikuchi, T.; Cotton, J.A.; Dalzell, J.J.; Hasegawa, K.; Kanzaki, N.; McVeigh, P.; Takanashi, T.; Tsai, I.J.; Assefa, S.A.; Cock, P.J.A.; et al. Genomic insights into the origin of parasitism in the emerging plant pathogen Bursaphelenchus xylophilus. PLoS Pathog. 2011, 7, e1002219. [Google Scholar] [CrossRef] [Green Version]
- Blaxter, M.L.; De Ley, P.; Garey, J.R.; Liu, L.X.; Scheldeman, P.; Vierstraete, A.; Vanfleteren, J.R.; Mackey, L.Y.; Dorris, M.; Frisse, L.M.; et al. A molecular evolutionary framework for the phylum Nematoda. Nature 1998, 392, 71–75. [Google Scholar] [CrossRef]
- Brenner, S. The genetics of Caenorhabditis elegans. Genetics 1974, 77, 71–94. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, T.; Jones, J.T.; Aikawa, T.; Kosaka, H.; Ogura, N. A family of glycosyl hydrolase family 45 cellulases from the pine wood nematode Bursaphelenchus xylophilus. FEBS Lett. 2004, 572, 201–205. [Google Scholar] [CrossRef] [Green Version]
- Proença, D.N.; Grass, G.; Morais, P.V. Understanding pine wilt disease: Roles of the pine endophytic bacteria and of the bacteria carried by the disease-causing pinewood nematode. Microbiologyopen 2017, 6, e00415. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Mota, M.; Vieira, P.; Butcher, R.A.; Sun, J. Interspecific communication between pinewood nematode, its insect vector, and associated microbes. Trends Parasitol. 2014, 30, 299–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- EPPO/OEPP Diagnostic protocols for regulated pests: Bursaphelenchus xylophilus. Bull. OEPP/EPPO 2009, 39, 344–353.
- Ikeda, T. Control program of Pine Wilt Disease for landscape conservation–The case of Amanohashidate, Kyoto, Japan. In Pine Wilt Disease: A Worldwide Threat to Forest Ecosystems; Springer: Dordrecht, The Netherlands, 2008; pp. 397–404. [Google Scholar]
- Petrini, O. Fungal endophytes of tree leaves. In Microbial Ecology of Leaves; Andrews, J., Hirano, S., Eds.; Brock/Springer series in contemporary bioscience; Springer: New York, NY, USA, 1991; pp. 179–197. ISBN 978-1-4612-3168-4. [Google Scholar]
- Proença, D.N.; Francisco, R.; Santos, C.V.; Lopes, A.; Fonseca, L.; Abrantes, I.M.O.; Morais, P.V. Diversity of bacteria associated with Bursaphelenchus xylophilus and other nematodes isolated from Pinus pinaster trees with pine wilt disease. PLoS ONE 2010, 5, e15191. [Google Scholar] [CrossRef] [Green Version]
- Alves, M.; Pereira, A.; Vicente, C.; Matos, P.; Henriques, J.; Lopes, H.; Nascimento, F.; Mota, M.; Correia, A.; Henriques, I. The role of bacteria in pine wilt disease: Insights from microbiome analysis. FEMS Microbiol. Ecol. 2018, 94, fiy077. [Google Scholar] [CrossRef]
- Vicente, C.S.L.; Nascimento, F.X.; Espada, M.; Barbosa, P.; Hasegawa, K.; Mota, M.; Oliveira, S. Characterization of bacterial communities associated with the pine sawyer beetle Monochamus galloprovincialis, the insect vector of the pinewood nematode Bursaphelenchus xylophilus. FEMS Microbiol. Lett. 2013, 347, 130–139. [Google Scholar] [CrossRef] [Green Version]
- Proença, D.N.; Schwab, S.; Vidal, M.S.; Baldani, J.I.; Xavier, G.R.; Morais, P.V. The nematicide Serratia plymuthica M24T3 colonizes Arabidopsis thaliana, stimulates plant growth, and presents plant beneficial potential. Brazilian J. Microbiol. 2019, 50, 777–789. [Google Scholar] [CrossRef]
- Proença, D.N.; Heine, T.; Senges, C.H.R.; Bandow, J.E.; Morais, P.V.; Tischler, D. Bacterial metabolites produced under iron limitation kill pinewood nematode and attract Caenorhabditis elegans. Front. Microbiol. 2019, 10, 2166. [Google Scholar] [CrossRef]
- Paiva, G.; Proença, D.N.; Francisco, R.; Verissimo, P.; Santos, S.S.; Fonseca, L.; Abrantes, I.M.O.; Morais, P.V. Nematicidal bacteria associated to pinewood nematode produce extracellular proteases. PLoS ONE 2013, 8, e79705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimazu, M. Use of microbes for control of Monochamus alternatus, vector of the invasive pinewood nematode. In Use of Microbes for Control and Eradication of Invasive Arthropods; Hajek, A.E., Glare, T.R., O’Callaghan, M., Eds.; Springer: Dordrecht, The Netherlands, 2009; Volume 6, pp. 141–157. ISBN 978-1-4020-8559-8. [Google Scholar]
- Matsuyama, T.; Fujita, M.; Yano, I. Wetting agent produced by Serratia marcescens. FEMS Microbiol. Lett. 1985, 28, 125–129. [Google Scholar] [CrossRef]
- Wasserman, H.H.; Keggi, J.J.; McKeon, J.E. The structure of Serratamolide1-3. J. Am. Chem. Soc. 1962, 84, 2978–2982. [Google Scholar] [CrossRef]
- Matsuyama, T.; Kaneda, K.; Nakagawa, Y.; Isa, K.; Hara-Hotta, H.; Yano, I. A novel extracellular cyclic lipopeptide which promotes flagellum-dependent and -independent spreading growth of Serratia marcescens. J. Bacteriol. 1992, 174, 1769–1776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuyama, T.; Murakami, T.; Fujita, M.; Fujita, S.; Yano, I. Extracellular vesicle formation and biosurfactant production by Serratia marcescens. Microbiology 1986, 132, 865–875. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Tanikawa, T.; Sato, Y.; Nakagawa, Y.; Matsuyama, T. Serratia marcescens gene required for surfactant serrawettin W1 production encodes putative aminolipid synthetase belonging to nonribosomal peptide synthetase family. Microbiol. Immunol. 2005, 49, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Pradel, E.; Zhang, Y.; Pujol, N.; Matsuyama, T.; Bargmann, C.I.; Ewbank, J.J. Detection and avoidance of a natural product from the pathogenic bacterium Serratia marcescens by Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 2007, 104, 2295–2300. [Google Scholar] [CrossRef] [Green Version]
- Su, C.; Xiang, Z.; Liu, Y.; Zhao, X.; Sun, Y.; Li, Z.; Li, L.; Chang, F.; Chen, T.; Wen, X.; et al. Analysis of the genomic sequences and metabolites of Serratia surfactantfaciens sp. nov. YD25T that simultaneously produces prodigiosin and serrawettin W2. BMC Genom. 2016, 17, 865. [Google Scholar] [CrossRef] [Green Version]
- Marques-Pereira, C.; Proença, D.N.; Morais, P.V. Genome sequences of Serratia strains revealed common genes in both serratomolides gene clusters. Biology. 2020, 9, 482. [Google Scholar] [CrossRef]
- Hirata, A.; Nakamura, K.; Nakao, K.; Kominami, Y.; Tanaka, N.; Ohashi, H.; Takano, K.T.; Takeuchi, W.; Matsui, T. Potential distribution of pine wilt disease under future climate change scenarios. PLoS ONE 2017, 12, e0182837. [Google Scholar] [CrossRef] [Green Version]
- Proença, D.N.; Fonseca, L.; Powers, T.O.; Abrantes, I.M.O.; Morais, P.V. Diversity of bacteria carried by pinewood nematode in USA and phylogenetic comparison with isolates from other countries. PLoS ONE 2014, 9, e105190. [Google Scholar] [CrossRef]
- Matsuyama, T.; Nakagawa, Y. Bacterial wetting agents working in colonization of bacteria on surface environments. Colloids Surf. B Biointerfaces 1996, 7, 207–214. [Google Scholar] [CrossRef]
- Nascimento, F.; Vicente, C.; Cock, P.; Tavares, M.; Rossi, M.; Hasegawa, K.; Mota, M. From plants to nematodes: Serratia grimesii BXF1 genome reveals an adaptation to the modulation of multi-species interactions. Microb. Genom. 2018, 4, e000178. [Google Scholar] [CrossRef] [PubMed]
- Roberts, D.P.; McKenna, L.F.; Lakshman, D.K.; Meyer, S.L.F.; Kong, H.; de Souza, J.T.; Lydon, J.; Baker, C.J.; Buyer, J.S.; Chung, S. Suppression of damping-off of cucumber caused by Pythium ultimum with live cells and extracts of Serratia marcescens N4-5. Soil Biol. Biochem. 2007, 39, 2275–2288. [Google Scholar] [CrossRef]
- Zhang, L.; Sun, J.; Hao, Y.; Zhu, J.; Chu, J.; Wei, D.; Shen, Y. Microbial production of 2,3-butanediol by a surfactant (serrawettin)-deficient mutant of Serratia marcescens H30. J. Ind. Microbiol. Biotechnol. 2010, 37, 857–862. [Google Scholar] [CrossRef] [PubMed]
- Weber, T.; Blin, K.; Duddela, S.; Krug, D.; Kim, H.U.; Bruccoleri, R.; Lee, S.Y.; Fischbach, M.A.; Müller, R.; Wohlleben, W.; et al. antiSMASH 3.0—A comprehensive resource for the genome mining of biosynthetic gene clusters. Nucleic Acids Res. 2015, 43, W237–W243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahul, S.; Chandrashekhar, P.; Hemant, B.; Chandrakant, N.; Laxmikant, S.; Satish, P. Nematicidal activity of microbial pigment from Serratia marcescens. Nat. Prod. Res. 2014, 28, 1399–1404. [Google Scholar] [CrossRef]
- Hellberg, J.E.E.U.; Matilla, M.A.; George, P.; Salmond, C. The broad-spectrum antibiotic, zeamine, kills the nematode worm Caenorhabditis elegans. Front. Microbiol. 2015, 6, 137. [Google Scholar] [CrossRef] [Green Version]
- Glater, E.E.; Rockman, M.V.; Bargmann, C.I. Multigenic natural variation underlies Caenorhabditis elegans olfactory preference for the bacterial pathogen Serratia marcescens. G3 (Bethesda) 2014, 4, 265–276. [Google Scholar] [CrossRef] [Green Version]
- Gibson, A.K.; Morran, L.T. A Model for Evolutionary Ecology of Disease: The case for Caenorhabditis nematodes and their natural parasites. J. Nematol. 2017, 49, 357–372. [Google Scholar] [CrossRef] [Green Version]
- Pujol, N.; Link, E.M.; Liu, L.X.; Kurz, C.L.; Alloing, G.; Tan, M.W.; Ray, K.P.; Solari, R.; Johnson, C.D.; Ewbank, J.J. A reverse genetic analysis of components of the Toll signaling pathway in Caenorhabditis elegans. Curr. Biol. 2001, 11, 809–821. [Google Scholar] [CrossRef] [Green Version]
- Heise, P.; Liu, Y.; Degenkolb, T.; Vogel, H.; Schäberle, T.F.; Marzinelli, E.M. Antibiotic-producing beneficial bacteria in the gut of the burying beetle Nicrophorus vespilloides. Front. Microbiol. 2019, 10, 1178. [Google Scholar] [CrossRef] [Green Version]
- Bargmann, C.I. Chemosensation in C. elegans. WormBook 2006, 1, 1–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Worthy, S.E.; Haynes, L.; Chambers, M.; Bethune, D.; Kan, E.; Chung, K.; Ota, R.; Taylor, C.J.; Glater, E.E. Identification of attractive odorants released by preferred bacterial food found in the natural habitats of C. elegans. PLoS ONE 2018, 13, e0201158. [Google Scholar] [CrossRef] [PubMed]
- Shanmugam, G.; Dubey, A.; Ponpandian, L.N.; Rim, S.O.; Seo, S.-T.; Bae, H.; Jeon, J. Genomic insights into nematicidal activity of a bacterial endophyte, Raoultella ornithinolytica MG against Pine Wilt Nematode. Plant Pathol. J. 2018, 34, 250. [Google Scholar] [CrossRef] [PubMed]
- Zečić, A.; Dhondt, I.; Braeckman, B.P. The nutritional requirements of Caenorhabditis elegans. Genes Nutr. 2019, 14, 15. [Google Scholar] [CrossRef] [PubMed]
- Proença, D.N.; Francisco, R.; Kublik, S.; Scholer, A.; Vestergaard, G.; Schloter, M.; Morais, P.V. The microbiome of endophytic, wood colonizing bacteria from pine trees as affected by Pine Wilt Disease. Sci. Rep. 2017, 7, 4205. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Korenblum, E.; de Araujo, L.; Guimarães, C.; de Souza, L.M.; Sassaki, G.; Abreu, F.; Nitschke, M.; Lins, U.; Freire, D.M.; Barreto-Bergter, E.; et al. Purification and characterization of a surfactin-like molecule produced by Bacillus sp. H2O-1 and its antagonistic effect against sulfate reducing bacteria. BMC Microbiol. 2012, 12, 252. [Google Scholar] [CrossRef] [Green Version]
- Kügler, J.H.; Muhle-Goll, C.; Hansen, S.H.; Völp, A.R.; Kirschhöfer, F.; Kühl, B.; Brenner-Weiss, G.; Luy, B.; Syldatk, C.; Hausmann, R. Glycolipids produced by Rouxiella sp. DSM 100043 and isolation of the biosurfactants via foam-fractionation. AMB Express 2015, 5, 82. [Google Scholar] [CrossRef]
- Stiernagle, T. Maintenance of C. elegans. WormBook 2006, 2, 51–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsueh, Y.; Gronquist, M.R.; Schwarz, E.M.; Nath, R.D.; Lee, C.; Gharib, S.; Schroeder, F.C.; Sternberg, P.W. Nematophagous fungus Arthrobotrys oligospora mimics olfactory cues of sex and food to lure its nematode prey. eLife 2017, 6, e20023. [Google Scholar] [CrossRef] [PubMed]
- R Core Development Team. R: A Language and Environment for Statistical Computing, R Version 4.1.2. 2021. Available online: https://www.r-project.org/ (accessed on 15 December 2021).











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Strain | PCR Code | PCR Product Length | Accession Number | Serratia Species | Gene | Identity % | Cover % | Accession Number |
|---|---|---|---|---|---|---|---|---|
| Serratia sp. Arv-20-4.2 | 8 | 555 | OK500210 | Serratia marcescens | swrW | 98.38% | 100% | AB193098 |
| Serratia sp. Arv-22-2.5c | 9 | 438 | OK500211 | Serratia marcescens | swrW | 98.87% | 100% | AB193098 |
| Serratia sp. Arv-29-3.11b | 11 | 447 | OK500212 | Serratia marcescens | swrW | 98.88% | 100% | AB193098 |
| Serratia sp. NBRC 102599T | 18 | 430 | OK500213 | Serratia marcescens | swrW | 72.52% | 100% | AB193098 |
| Serratia sp. AS13 | 19 | 452 | OK500214 | Serratia marcescens | swrW | 73.46% | 96% | AB193098 |
| Serratia sp. A88copa7 | 16 | 357 | OK500215 | Serratia liquefaciens | swrA | 71.27% | 94% | AF039572 |
| Serratia sp. A88copa13 | 17 | 455 | OK500216 | Serratia liquefaciens | swrA | 69.36% | 99% | AF039572 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marques-Pereira, C.; Proença, D.N.; Morais, P.V. The Role of Serratomolide-like Amino Lipids Produced by Bacteria of Genus Serratia in Nematicidal Activity. Pathogens 2022, 11, 198. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11020198
Marques-Pereira C, Proença DN, Morais PV. The Role of Serratomolide-like Amino Lipids Produced by Bacteria of Genus Serratia in Nematicidal Activity. Pathogens. 2022; 11(2):198. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11020198
Chicago/Turabian StyleMarques-Pereira, Catarina, Diogo Neves Proença, and Paula V. Morais. 2022. "The Role of Serratomolide-like Amino Lipids Produced by Bacteria of Genus Serratia in Nematicidal Activity" Pathogens 11, no. 2: 198. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11020198