1. Introduction
Cancer remains the most dangerous and widespread non-contagious disease that affects people around the world. Until now, traditional anti-cancer therapies such as chemotherapy, radiotherapy, resection, and radiosurgery have provided hope for the patient, but have often resulted in severe cytotoxicity. New solutions, such as anti-cancer therapy using viruses, is a promising method of treating many types of cancer. Since the introduction of the virus into the body stimulates an adaptive immune response, this strategy is considered as an immunotherapy [
1]. It uses both genetically modified, and naturally occurring viruses that selectively replicate and lead to the degradation of cancer cells without affecting normal cells [
2].
Equid alphaherpesvirus 1 (EHV-1) is a member of the subfamily
Alphaherpesvirinae and a major factor of the upper respiratory tract diseases, mare abortion, and neuropathies in horses. Like other alpha-herpesviruses (e.g., human herpesvirus type 1(HHV-1), human herpesvirus type 2 (HHV-2), varicella-zoster virus (VZV), pseudorabies virus (PRV), bovine herpesvirus type 1 (BHV-1)), it exhibits neurotropism and causes latent infection of the host cells [
3,
4]. A virus that adversely affects animal cells has been shown to be potentially helpful for humans. Courchesne et al. (2012) discovered the oncolytic potential of this virus and set out to investigate its effect on glioblastoma multiforme cells. One of the results of this groundbreaking study was the understanding of how the virus enters the glioblastoma cell via the Major Histocompatibility Complex (MHC-1) receptor [
5]. However, the changes in the cytoskeleton structure of A172 cells (human glioblastoma cell line) have not yet been examined. By expanding knowledge in this area, cytoskeleton can be used as a new immunotherapeutic target for oncolytic viruses.
The following structures are responsible for the movement of cancer cells: invadopodia, lamellipodia, and filopodia and depend on the reorganization of the actin cytoskeleton. On the other hand, actin-binding proteins (ABP) are responsible for controlling the polymerization and depolymerization process. Actin is a highly conserved protein that is found extensively in eukaryotic cells. There are two main forms—globular G-actin (monomer) and fibrillar F-actin (long-chain polymer) [
6]. The state of dynamic equilibrium between F- and G-actin results from changes in ABP in neoplastic cells, which results in incorrect regulation [
7].
The host cells cytoskeleton is also a crucial during replication and spreading of viruses [
8]. One of the first components of the cytoskeleton that the virus encounters in the process of penetrating the cell is actin network [
9]. Filamentous actin is responsible for some of the key functions of the cell such as cell integrity, formations of stress fibers, cell protrusions (filopodia and lamellipodia), the perinuclear and cell cortex. In context of viral replication, the most important feature, is polymerization of actin filaments which motors the crawling locomotion of eukaryotic cells and supports spread of viral particles. The layer of dense microfilaments beneath the plasma membrane (the cortical actin meshwork) is responsible for creating a physical barrier against viral infection. However, viruses use this structure to facilitate the processes of entry and post-entry pathways. In addition, actin filaments also influence the organization of surface receptors and membrane structures that mediate viral attachment, highlighting the complex and essential role of actin in early infection [
9,
10,
11]. In case of herpesviruses, it is known that they interact with actin filaments at various stages of their replication cycle, causing polymerization or fragmentation of filaments [
12,
13,
14,
15,
16]. EHV-1 influence on cytoskeleton was previously examined in Vero cells (african green monkey kidney), ED cell culture (equine dermis cells), and primary murine neurons. The results clearly showed that changes that occur under EHV-1 infection are depended on cells origination [
17,
18,
19,
20].
There are only a few data on alpha-herpesviruses infection and its effect on the cytoskeleton of glioblastoma multiforme cells. Most of the analysis focused on early-stage, in vitro studies investigating the aspect of genetic and molecular alterations in protein or marker expression. Clinical trials using herpesviruses, in particular HHV-1 (human herpesvirus type 1), have been conducted for years [
21,
22]. Application of EHV-1 as an oncolytic virus in the treatment of glioma cell lines was already proposed by Courchesne et al. (2012) [
5] and White and Frampton (2013) [
23]. In our previous study, we have demonstrated that animal virus EHV-1 productively replicated in the human adenocarcinoma cell line (A549) without the need for adaptation. Real-time PCR analysis and immunofluorescence assay showed that EHV-1 infected and caused lysis of human lung cancer cells [
20].
In the present study, we have evaluated the oncolytic potential of three EHV-1 strains on glioblastoma multiforme cells. Our results determined that EHV-1 productive infection presumably inhibits the migration of A172 cells by annihilation of actin cytoskeleton. Inhibition of A172 cell migration by EHV-1 infection may be an essential discovery relevant to glioblastoma multiforme therapy limiting tumor metastasis.
3. Discussion
The changes occurring in the cell cytoskeleton are closely related to the cell’s physiological state and the progression of tumor development [
24]. The three main components of the cytoskeleton are microtubules (MT), microfilaments (actin filaments, MF), and intermediate filaments (IF). This structure undergoes continuous remodeling and is responsible for migration, intracellular transport, and movement of organelles between cells [
25]. In the context of our research, we focused on EHV-1—microfilaments interaction. Actin filaments are involved in the internalization of membrane vesicles, protrusion of filopodia and lamellipodia, chemotaxis, cytokinesis and, most importantly, cell migration [
10]. They are also utilized by cells undergoing carcinogenesis and play an essential role in their migration. Tumor metastasis is still a vast problem, especially in malignant brain cancers. This phenomenon is remarkably relevant to GBM and is associated with increased actin polymerization so that the cells form protrusions and can migrate beyond the tumor mass area [
26]. Spatially controlled assembly of actin filaments generates lamellipodia, filopodia, and pseudopodia, which are used by cells to explore the extracellular space during invasion and metastasis [
27]. Taking it into consideration, we have discovered, for the first time, that EHV-1 inhibits migration of human glioblastoma cells (A172) by depolymerization of the actin cytoskeleton. This finding is crucial because GBM cells are notorious for their invasiveness, ability to develop resistance to chemo- and radiotherapy, and form secondary site tumors [
26]. Previous studies conducted by Courchesne and Frampton (2011; 2013) confirmed that EHV-1 has many beneficial properties, such as a well-defined growth cycle that results in the death of infected cells and a lack of pre-existing immunity to the virus, that make it a potential new anti-cancer agent. They assessed the oncolytic properties of the designed EHV-1 strains against malignant glioma—recombinant EHV-1 LacZ derived from RacL11 (L11ΔgIΔgE) with use of MHC-I as a cell entry receptor [
5,
23].
Data on alpha-herpesvirus infection and its effect on the cytoskeleton of glioblastoma multiforme cells are scarce. Most analyses focus on the preliminary research phase, in vitro studies in which genetic and molecular changes regarding protein or marker expression are investigated. Herpesvirus, adenovirus, vaccinia virus, reovirus, parvovirus, poliovirus, measles virus, replicating retrovirus vector, and Newcastle disease virus (NDV) are presently being tested in malignant glioma patients for safety and efficacy at different clinical phases. For years, clinical trials have been conducted using herpesviruses, particularly HSV-1 derivatives—G207, G47Δ, rQNestin34.5v.2, or C134. The G47Δ is the first OV approved for CNS tumor therapy and is a registered treatment for malignant glioma in Japan’s MHLW [
21,
22]. Unfortunately, no studies have described the changes occurring in the actin cytoskeleton of EHV-1-induced glioblastoma multiforme cells to date.
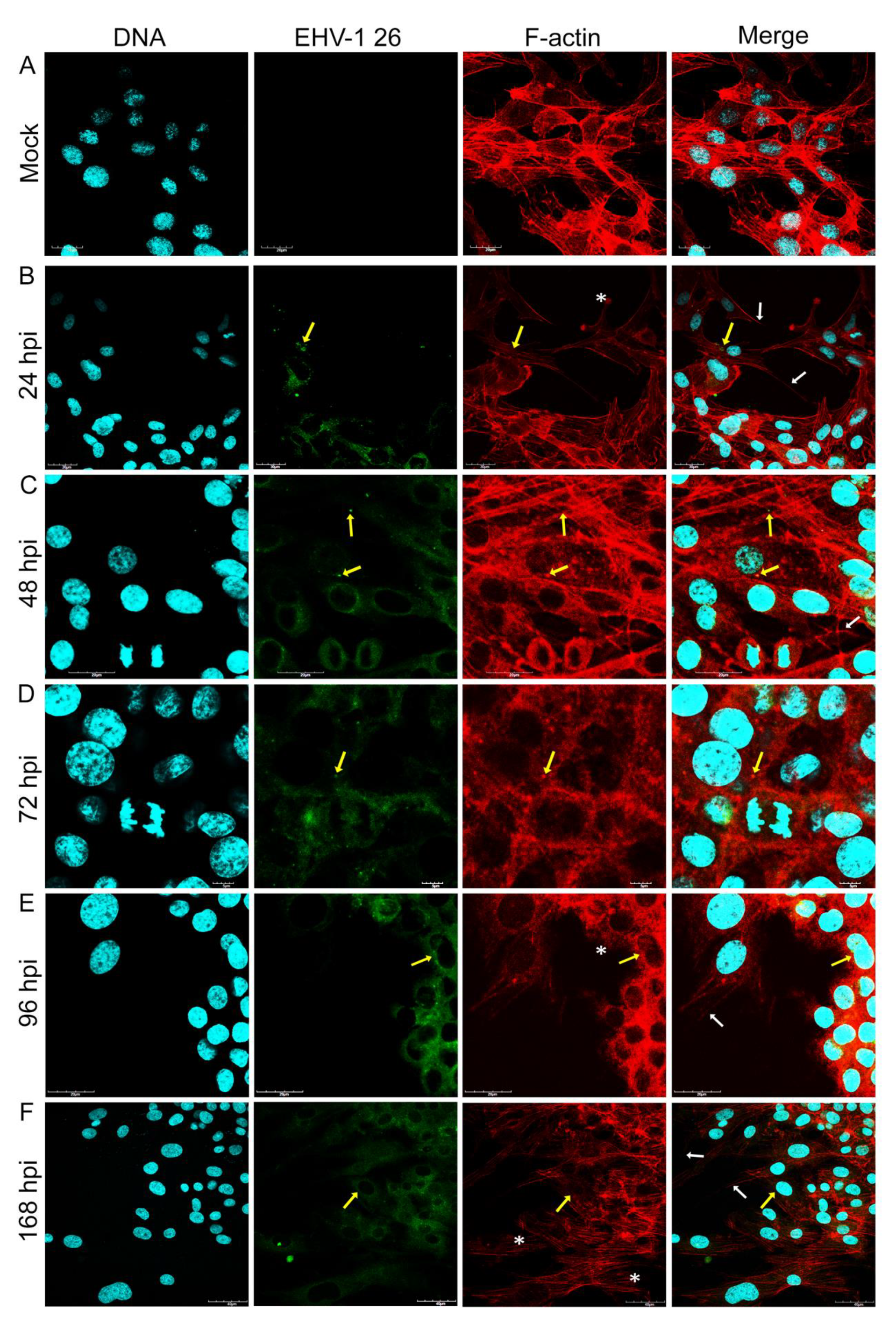
Our study provided the evaluation of the equid alphaherpesvirus 1 infection by confocal microscopy, JuLI™ Br live-time imaging, and Oris™FLEX kit, which revealed several changes in glioblastoma cells morphology and movement in case of all EHV-1 strains used. Analyzing the obtained data, we have observed not only the reorganization and depolarization of the actin cytoskeleton after infection with EHV-1 strains, but we have also noted the inhibition of cell motility. The changes observed by confocal microscopy were manifested by the formation of elongated actin protrusions—filopodia, intercellular bridges, and even cytoneme-like structures up to 48 hpi. It can be inferred that EHV-1 uses actin filaments as channels to transport virions between adjacent cells without driving the cell to the lysis pathway. Interestingly, during infection with Rac-H and Jan-E, cells have lost the ability to form protrusions after 48 hpi with visible depolymerization of actin and disruption of the filamentary network. In contrast, cells infected with EHV-1 26 after 72 hpi were characterized by the emergence of new protrusions with present viral antigen.
Particularly in the context of GBM, tunneling nanotubes (TNTs) are significant actin structures. These structures, which can be easily mistaken for filopodia, are responsible for cell-to-cell communication, transmission of signaling molecules in dendritic cells, small molecules (ions), macromolecules (proteins, nucleic acids), or even cell organelles (mitochondria, lysosomes) [
28,
29]. These structures are present in small amounts in cells of healthy, adult individuals. With the onset of disease and inflammation, their amount increases significantly. TNTs proliferate in response to increased oxidative stress caused by viral, bacterial infection, prions, or the process of carcinogenesis. Then, bacterial/viral protein, membrane fragments, or organelles may be exchanged [
30].
Considering that TNTs are activated in pathological states, it could be speculated that their increased amount appears in the initial times after EHV-1 infection and is controlled by the virus to facilitate the spread of progeny virions between cells. Subsequently, when the processes of polarization and depolarization of actin particles are completely disrupted by infection, GBM cells are unable to form protrusions which prevent them from migrating and exchanging signaling particles or metabolites.
Thus, it could be assumed that appropriate modulation of TNT structures and other mentioned actin structures by an agent such as EHV-1 infection could somehow reduce tumor growth and metastasis. This mechanism could be combined with the idea described in the new research investigating adaptation of TNTs by GBM cells to spread cell resistance to temozolomide (TMZ) and radiotherapy (IR) treatments. GMB utilizes structures as “highways” through which TMZ resistance can be spread via TNT-mediated transfer of MGMT (DNA repair enzyme O
6-methylguanine-DNA methyltransferase) [
30]. Thus, TNTs become a therapeutic target, and EHV-1 infection allows limiting/stopping the proliferation of these structures.
Obtained results were confirmed by Real-Time PCR replication analysis. The EHV-1 26 DNA copy number was significantly higher in samples collected from cells after 48 hpi. It could be assumed that EHV-1 26 in human GBM cells undergoes a specific latency state typical for herpesviruses. A specific knock-down of latency-related gene transcripts (LATs) should be considered in the context of potential use and further research of this strain. A different phenomenon was observed for Rac-H and Jan-E strains. In this case, after 48 hpi, the GBM cells do not regain the ability to form protrusions or intercellular bridges. The disappearance of actin filaments in the area of focal adhesion sites may indicate impaired migratory ability.
This hypothesis was confirmed by the analysis performed using the Oris™FLEX cell migration assessment kit. Analyzing the wound healing percentage diagrams and snapshots of real-time observation (JuLI™Br) of cell culture revealed that infection with all EHV-1 strains decreased the migration activity by inducing malformation in the actin cytoskeleton of the A172 cells. It is likely that EHV-1-infected glioma cells, particularly Jan-E and Rac-H strains, depolymerize actin filaments and thus cannot form cell protrusions that allow cells to move. The changes in actin cytoskeleton morphology compared to other neural cells of normal phenotype maybe since the hypoxic (hypoxia) environment coordinates changes in cytoskeleton dynamics, thus promoting migration of these cells. In addition, cell migration is often associated with excessive dynamic polymerization of actin at the leading edge of cells at focal adhesion sites, especially in cell protrusions [
26]. Those EHV-1 strains modify actin polymerization, initially promoting migration and then gradually affecting depolymerization of the actin cytoskeleton leading to lysis of tumor cells, thus blocking proliferation of A172 human glioblastoma multiforme cells. It was seen in the gradual lowering of confluence percentage on the wound healing diagram. It is crucial for GBM cells to rapidly polymerize actin, especially at the leading edges of the cell, to form protrusions. The other therapeutic strategies were examining the hypoxic environment and overexpression of the CTTN gene to disable this mechanism [
26].
The latest studies performed by Chodkowski et al. (2021) also revealed the possible oncolytic potential of EHV-1 Jan-E. The results indicated productive replication in the human adenocarcinoma cell line (A549) without additional adaptation of EHV-1. Real-time PCR analysis and immunofluorescence assay showed that EHV-1 infected and caused lysis of human lung cancer cells [
20]. Compering confocal analysis with our findings, we also have noted that in cells that did not undergo lysis accumulation, of viral antigens in the cytoplasm, accompanied by increased density of actin fibers, occurred. Despite different cell lines, comparable viral influence on actin cytoskeleton was detected. Also, a similar cytopathic effect was manifested in the A549 cell line, such as cell destruction, focal degeneration, and plaque formation. In the case of the A172 cell line, there were no sights of syncytia formation.
For the first time, the oncolytic potential of EHV-1 was described by Courchesne (2012), White and Frampton (2013), however, no observation of cytoskeletal or motile disturbances was investigated [
5,
23]. Detailed analysis of infection with a modified strain of EHV-1 (strain L11ΔgIΔgE, deletion of gI and gE glycoproteins, and embedded LacZ cassette) on five glioblastoma multiforme cell lines (A172, Hs 683, LN-18, SNB19, U251) was conducted to assess the oncolytic potential of the virus. EHV-1 productively infected four cell lines, and replication levels were positively correlated with glioma cell death. These findings were previously confirmed by Kurtz et al. (2010), who demonstrated that EHV-1 uses the MHC-1 complex to enter glioblastoma multiforme cells [
31]. Moreover, in cell lines A172 and LN-18 high level of expression of this complex was detected, thus confirming the high sensitivity of these lines to EHV-1 infection (supported by MTT cell viability assay) [
5,
23].
In other studies, utilization the histone deacetylase (HDAC) inhibitor and valproic acid (VPA) enhanced EHV-1 L11ΔgIΔgE infection and caused limited glioma progression, thus improving the therapeutic potential of this virus. The study revealed that treating two human glioma cell lines (U251 and SNB19) with VPA resulted in significantly more efficient virus entry, establishing a practical approach in using OVs in synergistic combination therapies to maximize therapeutic outcome [
23].
In conclusion, oncolytic virus (OV) therapy designates a promising new immune treatment strategy targeting cancer, particularly those located within the CNS. Oncolytic viruses can replicate in tumor cells but not in non-cancerogenic cells, resulting in lysis of the tumor-forming cell mass. Therefore, an effective viral vector used for potential therapeutic therapy should be safe, replicate rapidly, and lead to cell lysis. Many established researchers have conducted
in vitro and
in vivo studies using oncolytic viruses, including EHV-1, as vectors for anti-cancer therapy, combining viruses with drug therapy or using them alone [
5,
23,
31,
32,
33]. In contrast to already used HHV-1, EHV-1 is an animal virus that might naturally avoid the problem of rapid virus elimination by antibodies or memory cytotoxic T lymphocytes resulting from pre-existing immunity [
23,
34].
Our studies revealed another favorable property of EHV-1 virus proving its oncolytic potential. All strains of EHV-1 have affected polymerization of the actin cytoskeleton, thus blocking proliferation and migration of A172 human glioblastoma multiforme cells.
4. Materials and Methods
4.1. Cell Culture and Virus Maintenance
Human glioblastoma cell line A172 [ATCC CRL-1620™] was cultured in Dulbecco’s Modified Eagle Medium (DMEM) with 10% inactivated fetal bovine serum (FBS) and 40 mg/mL of streptomycin, penicillin (Gibco, Life Technologies, Waltham MA, USA) at 37 °C with 5% CO
2. Three different EHV-1 stains were used: non-neuropathogenic, reference strain Rac-H (149th passage in ED cells); non-neuropathogenic Jan-E (12th passage in ED cells) strain isolated from aborted fetus (neuropathogenicity confirmed by PCR-RFLP neuropathogenic/non-neuropathogenic discrimination test) [
35]; neuropathogenic EHV-1 strain (EHV-1 26, 15th passage in Vero cells) isolated from aborted fetus in Hungary in 2004 (neuropathogenicity confirmed by PriProET technique) [
36]. All strains were obtained from the virus collection of the Virology Laboratory at Warsaw University of Life Sciences (SGGW). The viruses were propagated in equine dermal (ED) and Vero cell cultures grown in Eagle’s minimum essential medium (MEM) (Gibco Life Technologies).
4.2. Infecting Cells with EHV-1
A172 cells were infected with Rac-H, Jan-E, and EHV-1 26 strains (MOI = 1.0) for 60 min at 37 °C. After incubation, the inoculum was removed, washed with PBS and fresh culture medium was added. Subsequently, infected cells were incubated for 24, 48, 72, 96, and 168 h at 37 °C with 5% CO2.
4.3. Immunofluorescence Staining and Imaging
The direct immunofluorescence method was used to visualize cell structures and viral antigen. After incubation with the viruses, A172 cells were washed twice in PBS (Sigma-Aldrich, Darmstadt, Germany), then fixed in 3.7% PFA (paraformaldehyde) for 30 min. After fixation the cells were washed twice with PBS solution and further incubated with 0.5% Tween/PBS solution for 5 min at room temperature. Subsequently, the culture was washed twice with PBS solution. Actin filaments were stained with 50 µL of TRITC-labelled phalloidin conjugate (500 ng/mL; Sigma-Aldrich) and incubated for 60 min in a wet chamber. In order to visualize the viral antigen of EHV-1 strains FITC-labelled anti-EHV-1 antibody (1:16; RPK Gamakon conjugate; Mevak, Nitra, Slovakia) was used. Cells infected with EHV-1 strains were covered with the conjugate and incubated for 30 min at 37 °C. Cell nuclei were stained with Hoechst 33258 (Thermo Fisher, Waltham MA, USA) for 2 min. Afterward, cover slips were mounted on microscope slides using ProLong Gold Antifade Mounting Medium (Thermo Fisher). Confocal images were acquired in a confocal microscope (Fluoview FV10i, Olympus, Warsaw, Poland) and analyzed using FV10i software (Olympus), ImageJ (NIH Image, version 1.53a, Bethesda, MD, USA), and Adobe Photoshop CS6 software (Adobe Systems Incorporated, ver. 13.0, San Jose, CA, USA).
4.4. Quantitative Real-Time PCR
Viral DNA was isolated from samples collected after 24, 48, 72, 96, 168 hpi, separately from A172 cells and cell culture medium. The quantity of the EHV-1 DNA in all samples was determined using quantitative real-time PCR (qPCR) with fluorescent TaqMan probes, complementary for the sequence within the amplified products. For amplification of viral DNA, the primers specific for the glycoprotein B (gB) gene were used [gB-1, 5′–AAA CAA AGA GCG GAC CCT AT–3′; gB-2, 5′–TCC GTG AAA ATC TCG TTC TC–3′]. Amplification was performed with LightCycler TaqMan Master Kit (Roche Diagnostics, Mannheim, Germany). To obtain a standard curve, Jan-E EHV-1 strain was serially diluted in sterile deionized water to give ranges of CCID
50 = 10
6. DNA of the Rac-H strain was used as a positive control. The negative control was water included in the LightCycler TaqMan Master kit. Fluorescence level was measured at 530 nm wavelength. Tests were run on the LightCycler 2.0 instrument (Roche Diagnostics, Mannheim, Germany) according to the in-house quantitative method [
3,
37].
4.5. Real-Time Cell Growth Analise JuLi™ Br
To determine cellular growth and morphology of glioblastoma cells infected with various EHV-1 strains, JuLI TM Br Live Cell—system for bright-field analysis (NanoEnTek, Seoul, Korea 2015) was used. The A172 cell cultures that reached > 95% confluency were infected with Jan-E, Rac-H, and EHV-1 26 strains as previously described. Images were captured for 168 h with 30 min intervals. The results were obtained and analyzed using JuLI TM Br PC software. Uninfected cells were used as a negative control. All images were captured at a ×40 magnification.
4.6. Migration Assay—The OrisTM Cell Migration Assembly Kit
The technology utilizes a 96-well plate populated with silicone-based cell seeding stoppers which exclude cells from attaching to a central zone.
A172 cells were seeded at a density of 5 × 10
4 into an Oris™ 96-well plate with attached Oris™ silicon stoppers (Platypus Technologies, LLC, Madison, WI, USA). After the cells were seeded and allowed to adhere, the stoppers were removed to reveal a 2 mm diameter exclusion zone into which cells may then migrate [
38]. After 18 h of attachment and reaching full confluency, the stoppers were removed. Then the cell cultures were infected with three EHV-1 strains for 1 h at 37 °C. Afterwards, the cell cultures were washed twice with PBS and cultured with fresh serum-free medium for 168 h at 37 °C, 5% CO
2. The Oris™ plate was placed under the JuLI™ Br—Live Cell Movie Analyzer (NanoEnTek) (
Figure 8). During the incubation period of the migration test, the JuLI™ Br camera has taken photos in 5 min intervals. The complete cell growth movie was cut to obtain from 0 to168 h post-infection with each EHV-1 strain to collect the final image. To evaluate the stage of migration inhibition, the wound healing diagrams were analyzed.
4.7. Statistical Analysis
The results were statistically evaluated by one-way analysis of variation (ANOVA) using the Student-Newman-Keuls multiple comparisons test and the Turkey-Kramer multiple comparisons test. These analyses were performed using GraphPad InStatTM version 3.0 software (GraphPad Software Inc., San Diego, CA, USA). Statistical differences were interpreted as significant at p ≤ 0.05, highly significant at p ≤ 0.01, extremely significant at p ≤ 0.001 and not significant at p > 0.05.


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}








