
Hagnosa longicapillata, gen. nov., sp. nov., a New Sordariaceous Ascomycete in the Indoor Environment, and the Proposal of Hagnosaceae fam. nov.



Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
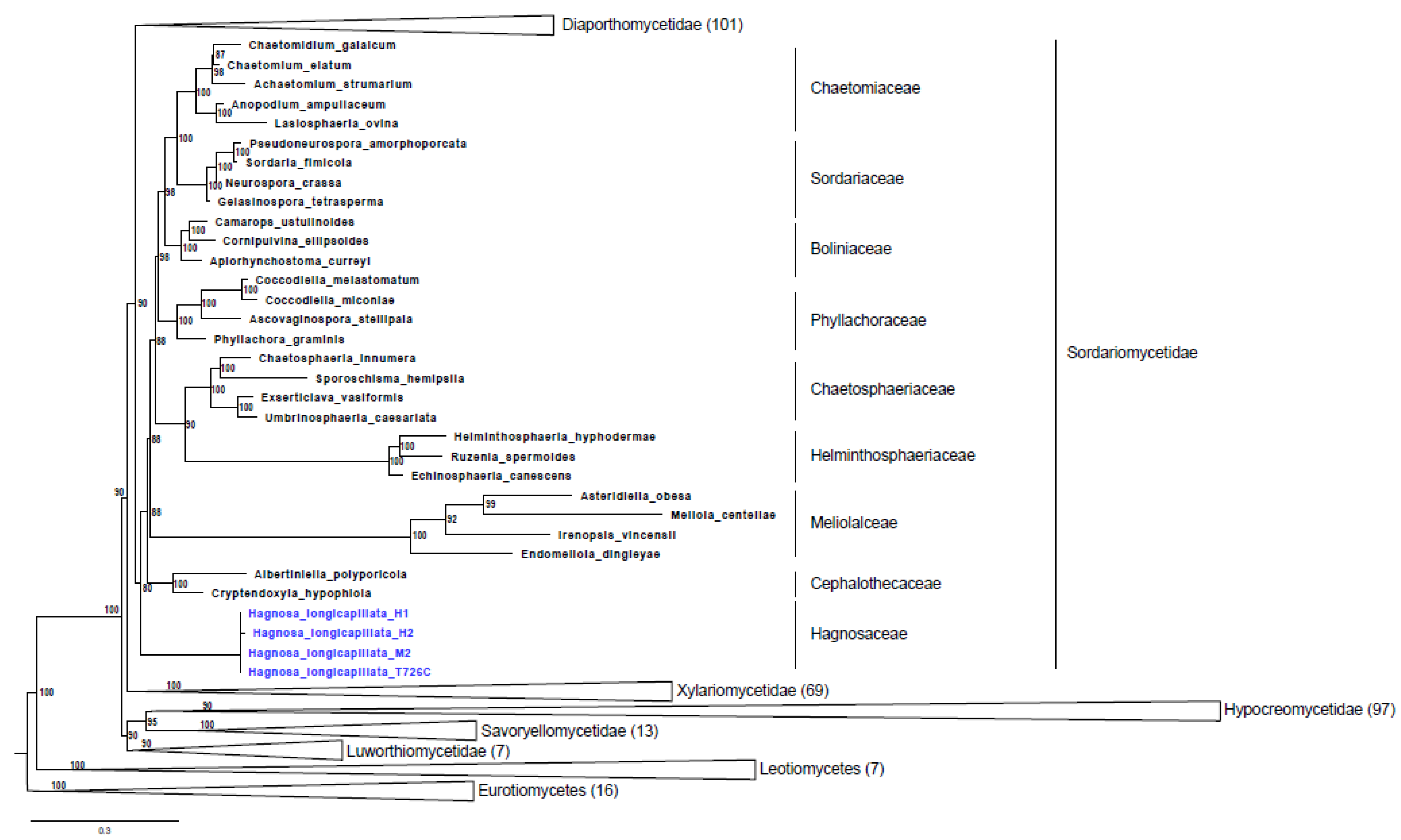
2.1. Phylogenetic Analysis
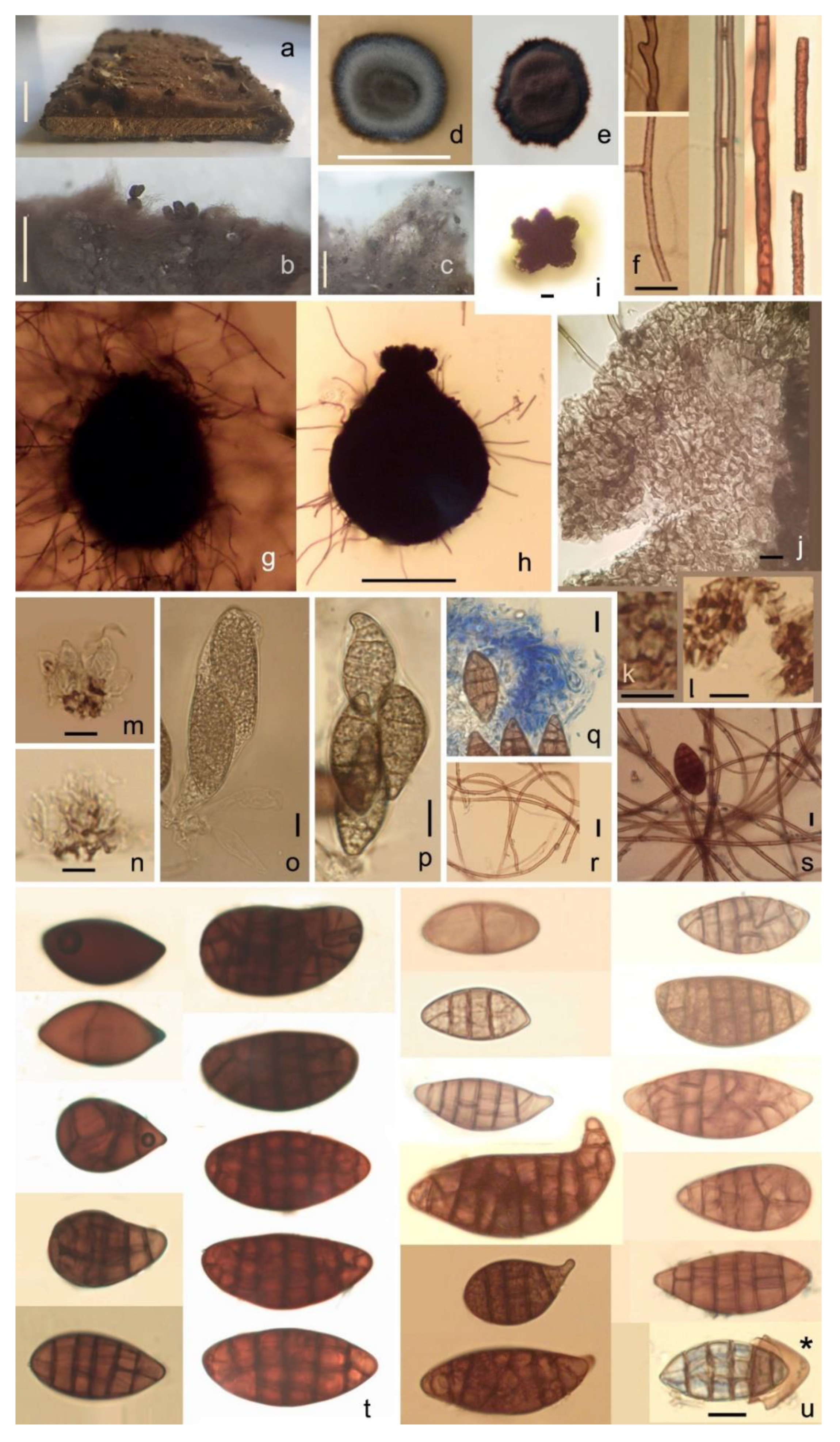
2.2. Taxonomy
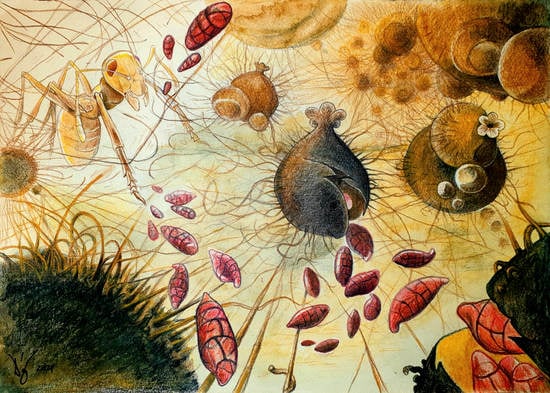
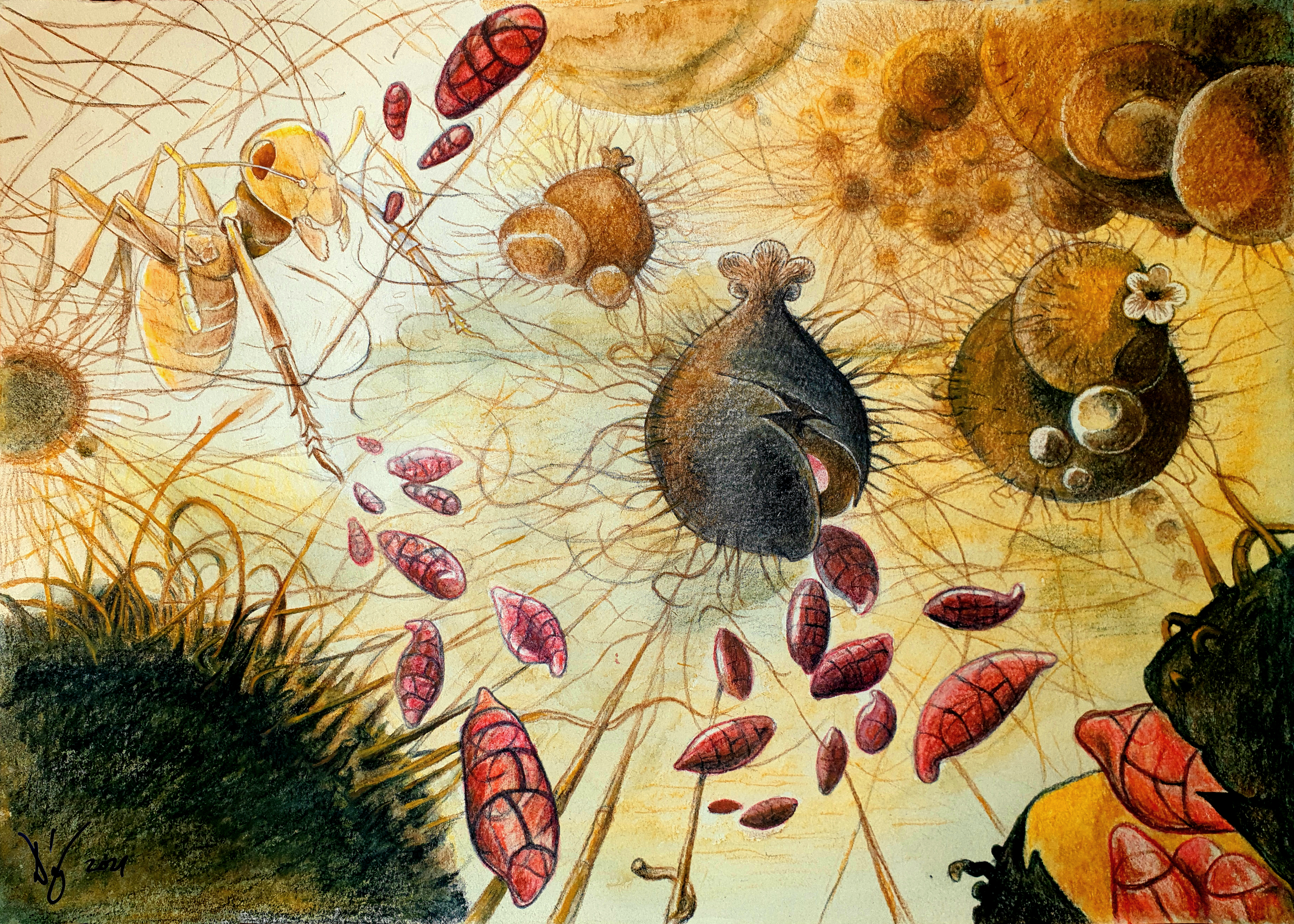
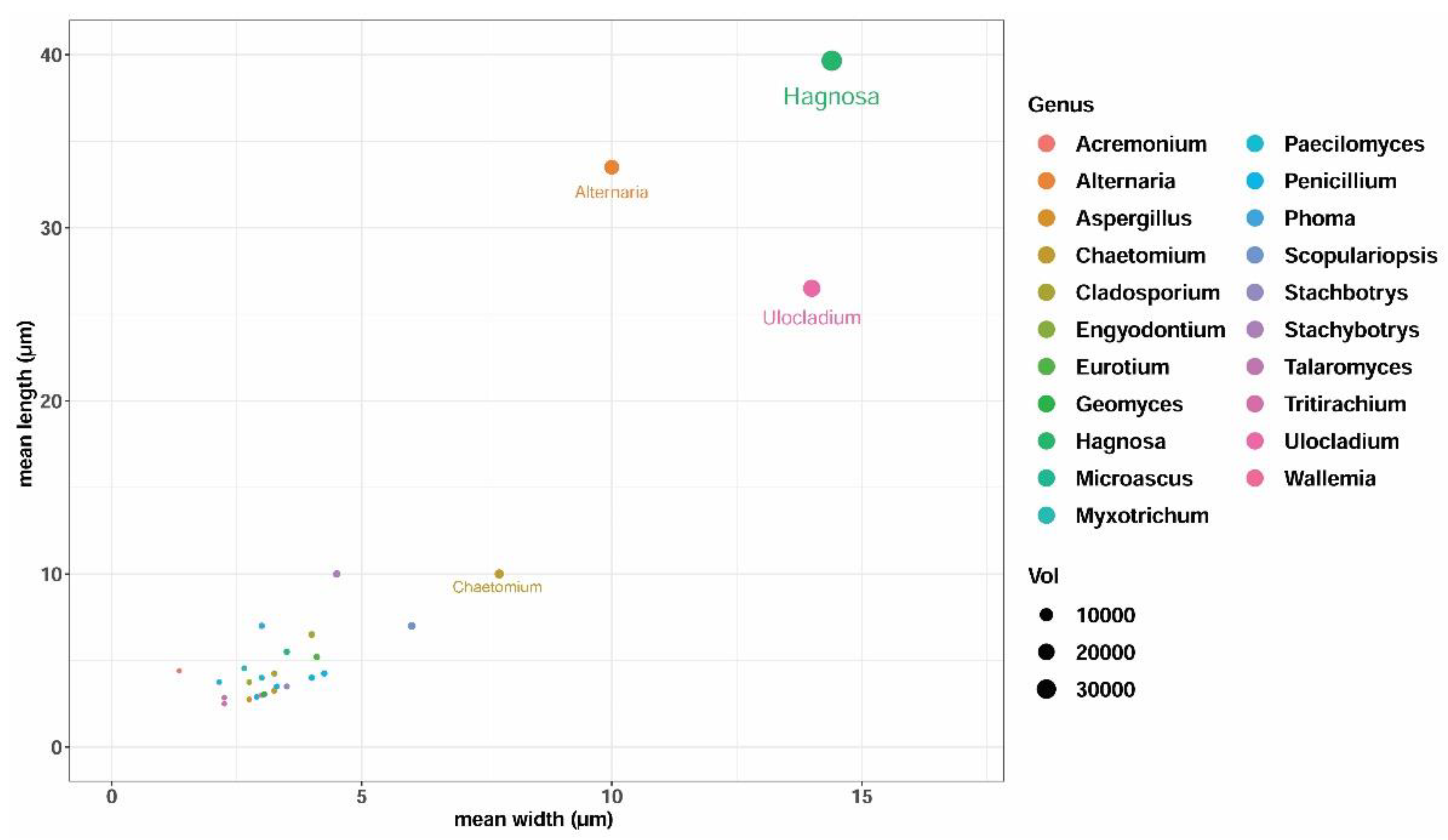
2.3. Fungal Ecology and Dispersal
3. Discussion
4. Materials and Methods
4.1. Isolation
4.2. Molecular Phylogeny
4.3. Ecological Experiments
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Flannigan, B.; Samson, R.A.; Miller, J.D. Microorganisms in Home and Indoor Work Environments: Diversity, Health Impacts, Investigation and Control; CRC Press: London, UK, 2002. [Google Scholar]
- Li, D.W.; Yang, C.S. Notes on indoor fungi I: New records and noteworthy fungi from indoor environments. Mycotaxon 2004, 89, 473–488. [Google Scholar]
- Hongsanan, S.; Maharachchikumbura, S.S.; Hyde, K.D.; Samarakoon, M.C.; Jeewon, R.; Zhao, Q.; Bahkali, A.H. An updated phylogeny of Sordariomycetes based on phylogenetic and molecular clock evidence. Fungal Divers. 2017, 84, 25–41. [Google Scholar] [CrossRef]
- Sánchez, R.M.; Miller, A.N.; Bianchinotti, M.V. New species of Capronia (Herpotrichiellaceae, Ascomycota) from Patagonian forests, Argentina. Plant Fungal Syst. 2019, 64, 81–90. [Google Scholar] [CrossRef] [Green Version]
- Müller, E.; Petrini, O.; Fisher, P.J.; Samuels, G.J.; Rossman, A.Y. Taxonomy and anamorphs of the Herpotrichiellaceae with notes on generic synonymy. Trans. Br. Mycol. Soc. 1987, 88, 63–74. [Google Scholar] [CrossRef]
- Kirk, P.M.; Cannon, P.F.; Minter, D.W.; Stalpers, J.A. Dictionary of the Fungi, 10th ed.; CAB International: Wallingford, UK, 2008. [Google Scholar]
- Eriksson, O.E.; Yue, J.Z. An amended description and disposition of the genus Thyridium. Syst. Ascomycetum 1989, 8, 9–16. [Google Scholar]
- Checa, J.; Blanco, M.N.; Moreno, G. Contributions to the family Thyridiaceae. New data on Sphaeria mutabilis. Mycotaxon 2013, 125, 149–164. [Google Scholar] [CrossRef]
- Huhndorf, S.M.; Miller, A.N.; Fernández, F.A. Molecular systematics of the Sordariales: The order and the family Lasiosphaeriaceae redefined. Mycologia 2004, 96, 368–387. [Google Scholar] [CrossRef]
- Réblova, M.; Barr, M.E.; Samuels, G.J. Chaetosphaeriaceae, a new family for Chaetosphaeria and its relatives. Sydowia 1999, 51, 49–70. [Google Scholar]
- Maharachchikumbura, S.S.; Hyde, K.D.; Jones, E.G.; McKenzie, E.H.C.; Bhat, J.D.; Dayarathne, M.C.; Wijayawardene, N.N. Families of Sordariomycetes. Fungal Divers. 2016, 79, 1–317. [Google Scholar] [CrossRef]
- Magyar, D.; Kállai, Z.; Sipiczki, M.; Dobolyi, C.; Sebők, F.; Beregszászi, T.; Bihari, Z.; Kredics, L.; Oros, G. Survey of viable airborne fungi in wine cellars of Tokaj, Hungary. Aerobiologia 2018, 34, 171–185. [Google Scholar] [CrossRef]
- de Hoog, G.S.; Guarro, J.; Gené, J.; Figueras, M.J. Atlas of Clinical Fungi, 2nd ed.; Centraalbureau Voor Schimmelcultures: Utrecht, The Netherlands, 2000. [Google Scholar]
- Samson, R.A.; Houbraken, J.; Thrane, U.; Frisvad, J.C.; Andersen, B. Food and Indoor Fungi; Westerdijk Fungal Biodiversity Institute: Utrecht, The Netherlands, 2019. [Google Scholar]
- Gregory, P.H. The Microbiology of the Atmosphere; Leonard Hill: London, UK, 1961; p. 83. [Google Scholar]
- Ingold, C.T. Fungal Spores: Their Liberation and Dispersal; Clarendon Press: Oxford, UK, 1971; p. 197. [Google Scholar]
- Bánhegyi, J.; Tóth, S.; Ubrizsy, G.; Vörös, J. Identification Book of Microfungi in Hungary; Akadémiai Kiadó: Budapest, Hungary, 1985; pp. 5–511. [Google Scholar]
- Kedves, O.; Kocsubé, S.; Bata, T.; Andersson, M.A.; Salo, J.M.; Mikkola, R.; Salonen, H.; Szűcs, A.; Kedves, A.; Kónya, Z.; et al. Chaetomium-like species from European indoor environments include Dichotomopilus finlandicus sp. nov. Pathogens 2021, 10, 1133. [Google Scholar] [CrossRef] [PubMed]
- von Arx, J.A.; Guarro, J.; Figueras, M.J. The ascomycete genus Chaetomium. Beih. Zur Nova Hedwig. 1986, 84, 162. [Google Scholar]
- Hölldobler, B.E.; Wilson, E.O. The Ants; The Belknap Press of Harvard University Press: Cambridge, MA, USA, 1990; p. 746. [Google Scholar]
- Pereira, H.; Detrain, C. Pathogen avoidance and prey discrimination in ants. R. Soc. Open Sci. 2020, 7, 191705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maraun, M.; Migge, S.; Schaefer, M.; Scheu, S. Selection of microfungal food by six oribatid mite species (Oribatida, Acari) from two different beech forests. Pedobiologia 1998, 42, 232–240. [Google Scholar]
- Maraun, M.; Martens, H.; Migge, S.; Theenhaus, A.; Scheu, S. Adding to ‘the enigma of soil animal diversity’: Fungal feeders and saprophagous soil invertebrates prefer similar food substrates. Eur. J. Soil Biol. 2003, 39, 85–95. [Google Scholar] [CrossRef]
- Renker, C.; Otto, P.; Schneider, K.; Zimdars, B.; Maraun, M.; Buscot, F. Oribatid mites as potential vectors for soil microfungi: Study of mite-associated fungal species. Microb. Ecol. 2005, 50, 518–528. [Google Scholar] [CrossRef]
- Schneider, K.; Maraun, M. Feeding preferences among dark pigmented fungal taxa (“Dematiacea”) indicate limited trophic niche differentiation of oribatid mites (Oribatida, Acari). Pedobiologia 2005, 49, 61–67. [Google Scholar] [CrossRef]
- Bailey, I. Some relations between ants and fungi. Ecology 1920, 1, 174–189. [Google Scholar] [CrossRef]
- Guedes, F.L.A.; Attili-Angelis, D.; Pagnocca, F.C. Selective isolation of dematiaceous fungi from the workers of Atta laevigata (Formicidae: Attini). Folia Microbiol. 2012, 57, 21–26. [Google Scholar] [CrossRef]
- Magyar, D.; Vass, M.; Li, D.W. Dispersal Strategies of Microfungi. In Biology of Microfungi; Li, D.W., Ed.; Springer International Publishing: London, UK, 2016; pp. 315–371. [Google Scholar]
- Baral, H.O. Vital versus herbarium taxonomy: Morphological differences between living and dead cells of Ascomycetes, and their taxonomic implications. Mycotaxon 1992, 44, 333–390. [Google Scholar]
- Puchkov, E. Image analysis in microbiology: A review. J. Comput. Commun. 2016, 4, 8. [Google Scholar] [CrossRef] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- O’Mahony, E.M.; Tay, W.T.; Paxton, R.J. Multiple rRNA variants in a single spore of the microsporidian Nosema bombi. J. Eukaryot. Microbiol. 2007, 54, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes—application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Toh, H. Recent developments in the MAFFT multiple sequence alignment program. Brief. Bioinform. 2008, 9, 286–298. [Google Scholar] [CrossRef] [Green Version]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; Von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A. Fig Tree, Ver. 1.3.1. 2009. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 10 February 2022).
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer-Verlag: New York, NY, USA, 2016; Available online: https://ggplot2.tidyverse.org (accessed on 31 March 2022).
- Chen, J. Advancement on techniques for the separation and maintenance of the red imported fire ant colonies. Insect Sci. 2007, 14, 1–4. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Magyar, D.; Tartally, A.; Merényi, Z. Hagnosa longicapillata, gen. nov., sp. nov., a New Sordariaceous Ascomycete in the Indoor Environment, and the Proposal of Hagnosaceae fam. nov. Pathogens 2022, 11, 593. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11050593
Magyar D, Tartally A, Merényi Z. Hagnosa longicapillata, gen. nov., sp. nov., a New Sordariaceous Ascomycete in the Indoor Environment, and the Proposal of Hagnosaceae fam. nov. Pathogens. 2022; 11(5):593. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11050593
Chicago/Turabian StyleMagyar, Donát, András Tartally, and Zsolt Merényi. 2022. "Hagnosa longicapillata, gen. nov., sp. nov., a New Sordariaceous Ascomycete in the Indoor Environment, and the Proposal of Hagnosaceae fam. nov." Pathogens 11, no. 5: 593. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11050593