
Inhibitor of Cysteine Protease of Plasmodium malariae Regulates Malapains, Endogenous Cysteine Proteases of the Parasite

 , ,
, ,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
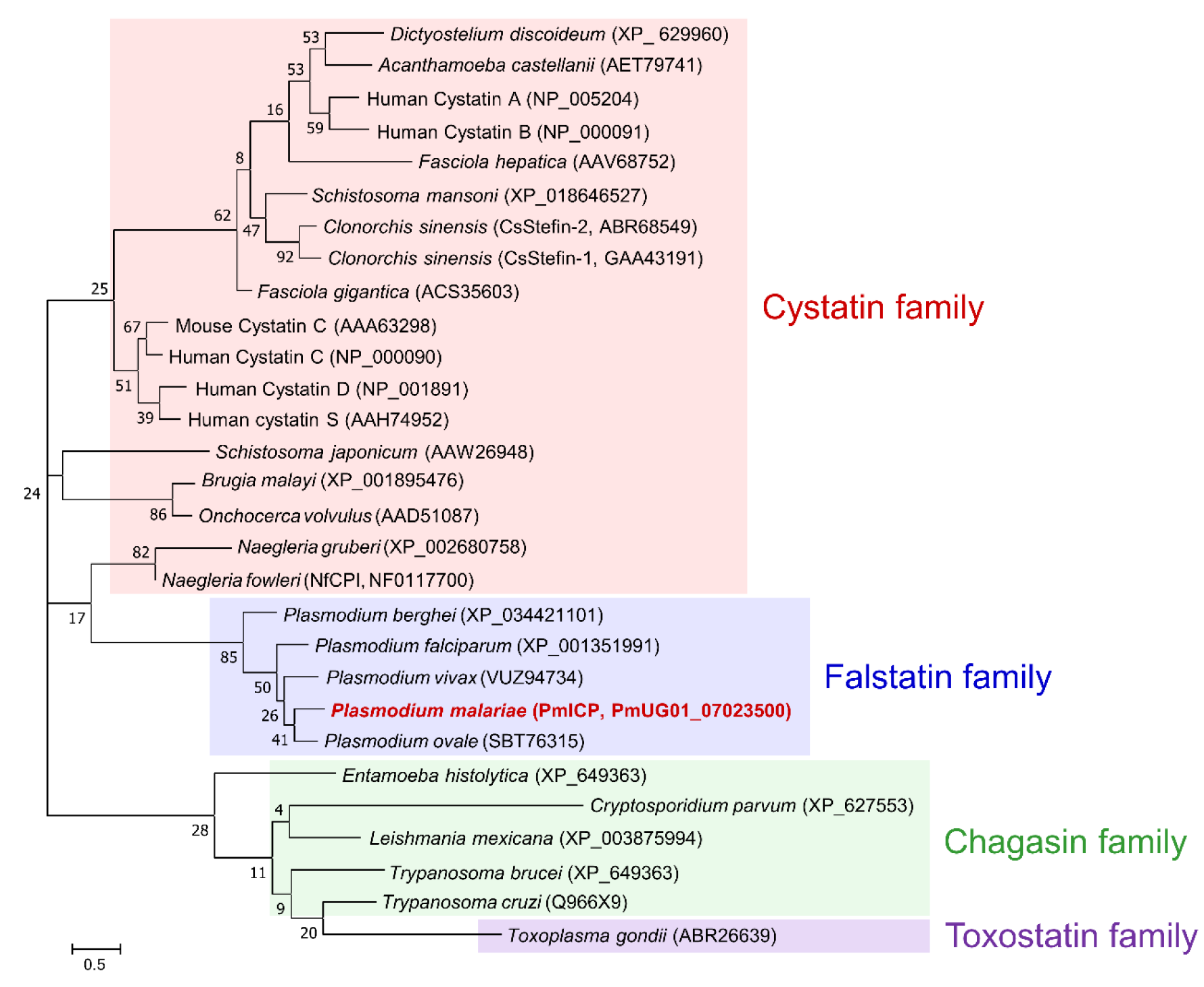
2.1. Sequence Analysis of the Gene Encoding PmICP
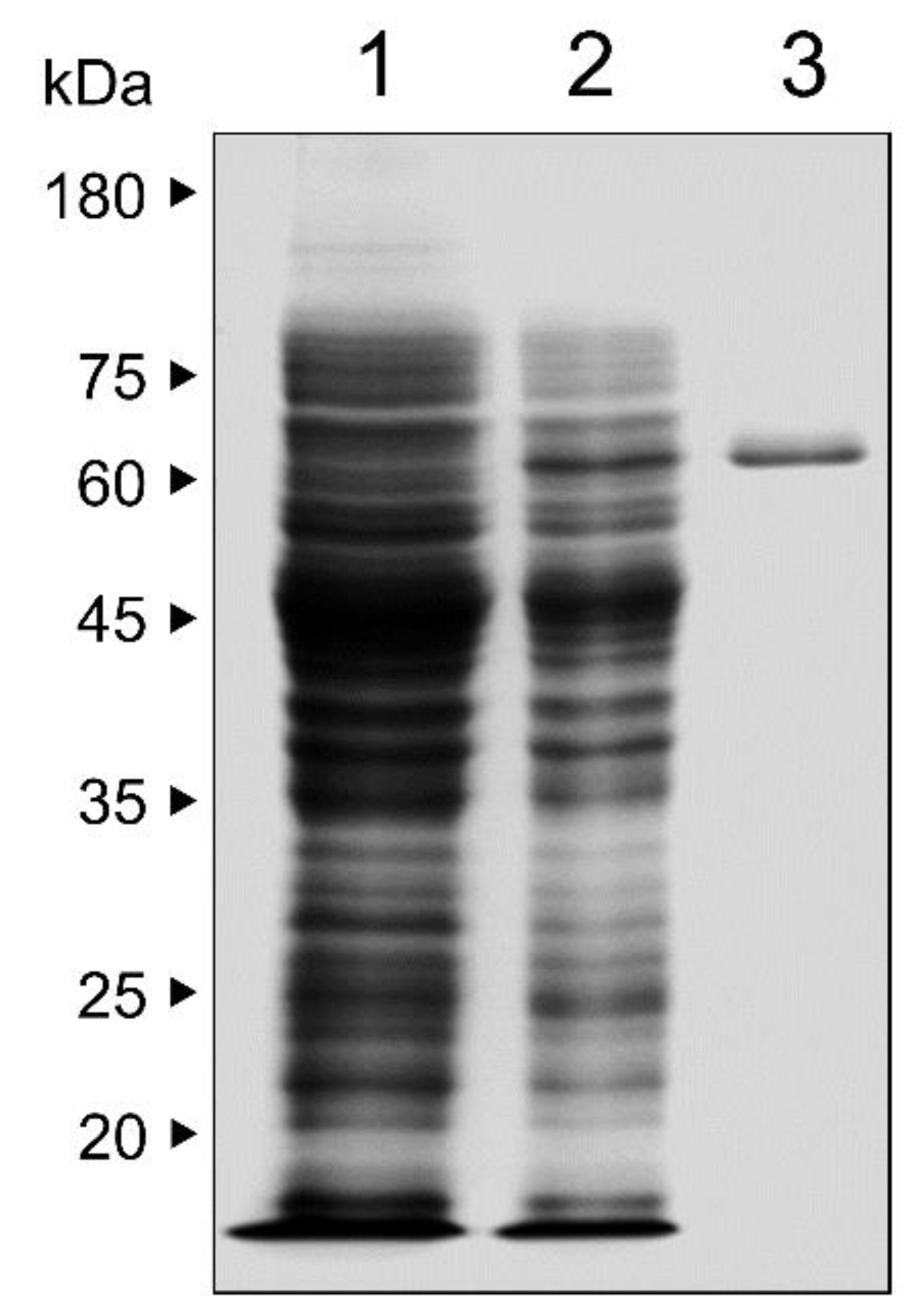
2.2. Expression and Purification of Recombinant PmICP
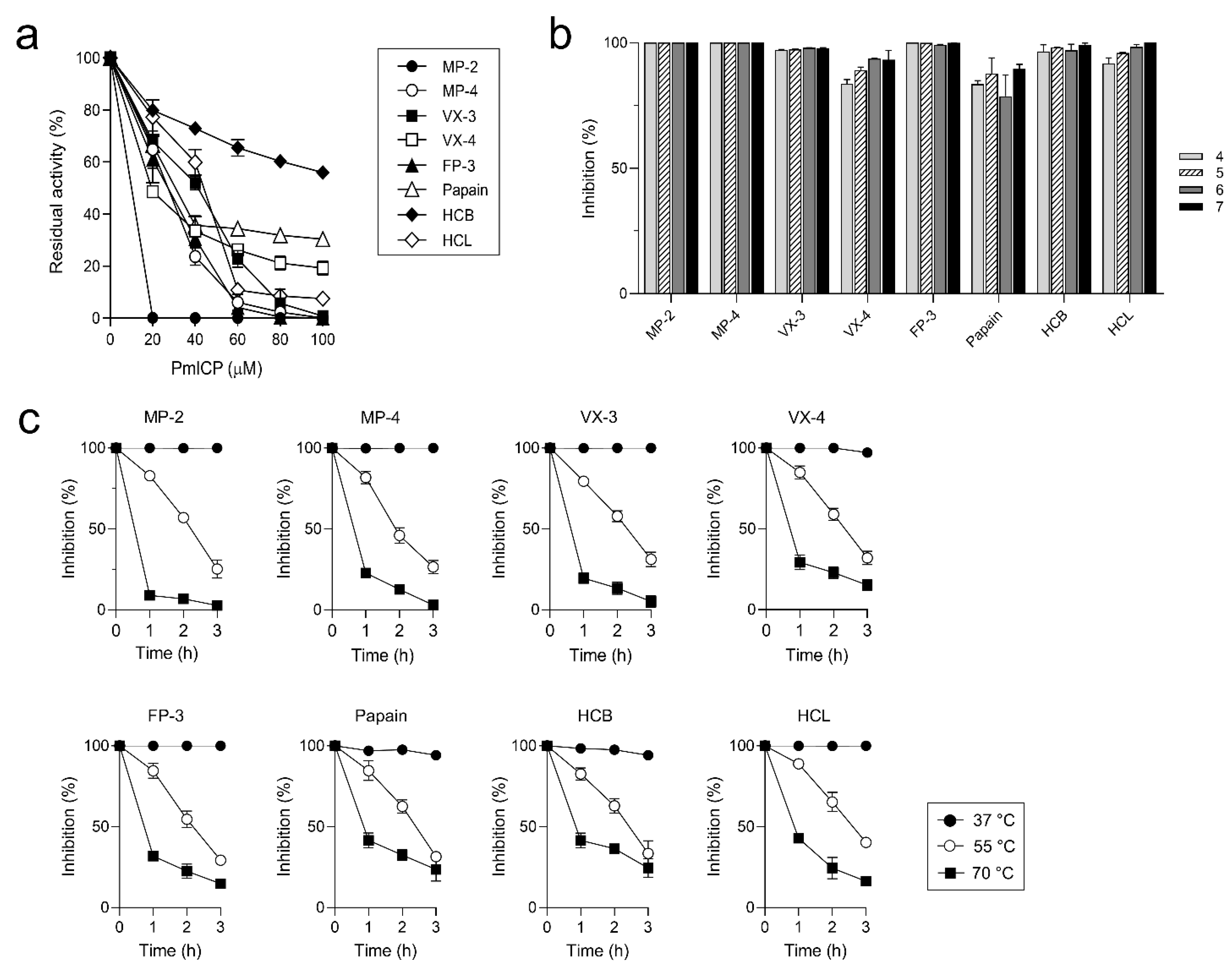
2.3. Biochemical Properties of PmICP
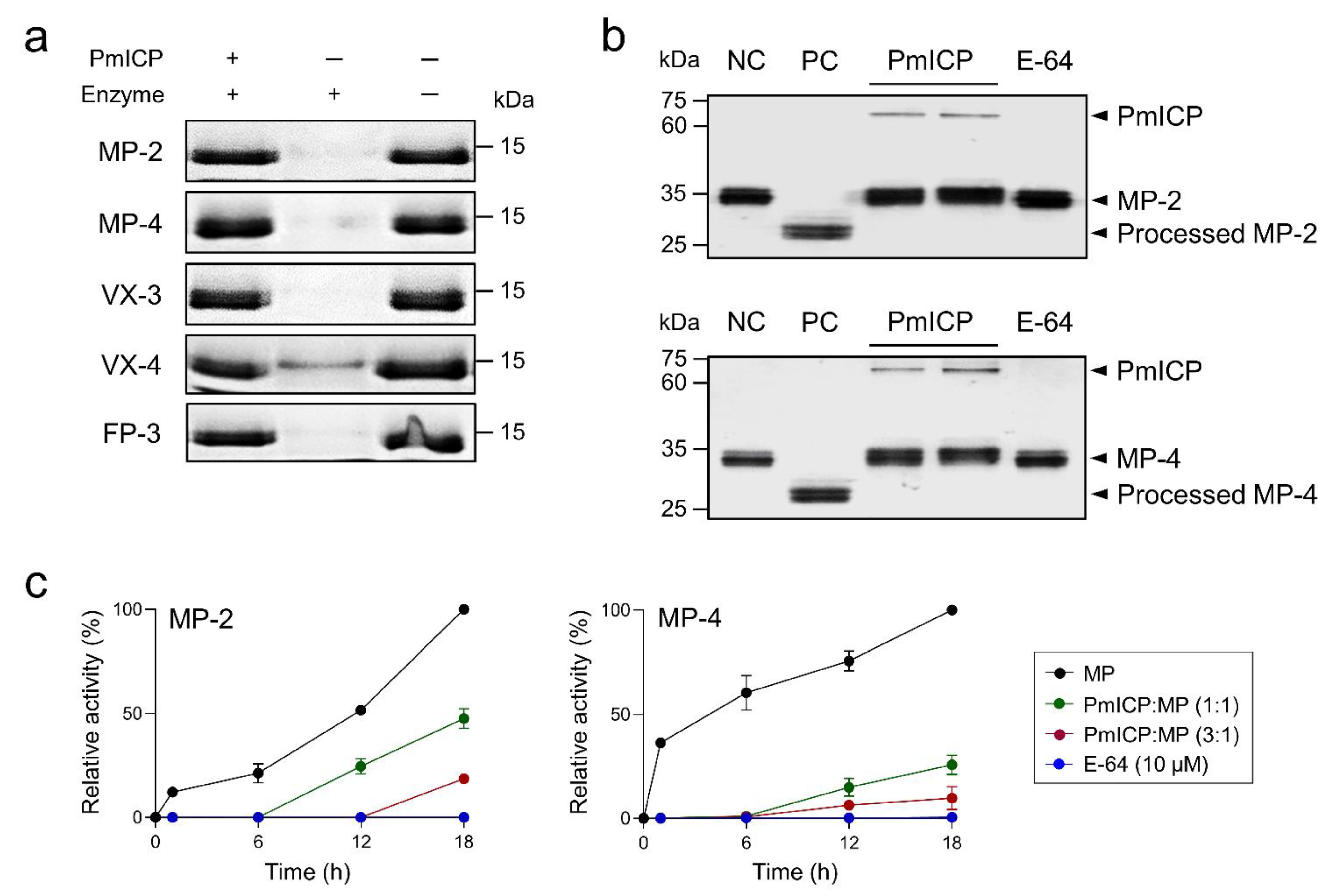
2.4. PmICP as a Regulatory Molecule of MPs
3. Discussion
4. Materials and Methods
4.1. Cloning of a Gene Encoding PmICP
4.2. Expression and Purification of Recombinant PmICP
4.3. Inhibitory Activity Assay
4.4. Characterization of Biochemical Properties of PmICP
4.5. Inhibition of Hemoglobin Hydrolysis by MPs
4.6. Regulatory Activity of PmICP on Maturation of MPs
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rosenthal, P.J. Falcipains and other cysteine proteases of malaria parasites. Adv. Exp. Med. Biol. 2011, 712, 30–48. [Google Scholar] [CrossRef] [PubMed]
- Rosenthal, P.J. Falcipain cysteine proteases of malaria parasites: An update. Biochim. Biophys. Acta-Proteins Proteom. 2020, 1868, 140362. [Google Scholar] [CrossRef] [PubMed]
- Qidwai, T. Hemoglobin degrading proteases of Plasmodium falciparum as antimalarial drug targets. Curr. Drug Targets 2015, 16, 1133–1141. [Google Scholar] [CrossRef]
- Ettari, R.; Previti, S.; Di Chio, C.; Zappalà, M. Falcipain-2 and Falcipain-3 Inhibitors as Promising Antimalarial Agents. Curr. Med. Chem. 2020, 28, 3010–3031. [Google Scholar] [CrossRef]
- Sijwali, P.S.; Rosenthal, P.J. Gene disruption confirms a critical role for the cysteine protease Falcipain-2 in hemoglobin hydrolysis by Plasmodium falciparum. Proc. Natl. Acad. Sci. USA 2004, 101, 4384–4389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sijwali, P.S.; Koo, J.; Singh, N.; Rosenthal, P.J. Gene disruptions demonstrate independent roles for the four falcipain cysteine proteases of Plasmodium falciparum. Mol. Biochem. Parasitol. 2006, 150, 96–106. [Google Scholar] [CrossRef] [PubMed]
- Blackman, M.J. Malarial proteases and host cell egress: An “emerging” cascade. Cell. Microbiol. 2008, 10, 1925–1934. [Google Scholar] [CrossRef] [Green Version]
- Greenbaum, D.C.; Baruch, A.; Grainger, M.; Bozdech, Z.; Medzihradszky, K.F.; Engel, J.; DeRisi, J.; Holder, A.A.; Bogyo, M. A role for the protease Falcipain 1 in host cell invasion by the human malaria parasite. Science 2002, 298, 2002–2006. [Google Scholar] [CrossRef] [Green Version]
- Hopp, C.S.; Bennett, B.L.; Mishra, S.; Lehmann, C.; Hanson, K.K.; Lin, J.W.; Rousseau, K.; Carvalho, F.A.; van der Linden, W.A.; Santos, N.C.; et al. Deletion of the rodent malaria ortholog for Falcipain-1 highlights differences between hepatic and blood stage merozoites. PLoS Pathog. 2017, 13, e1006586. [Google Scholar] [CrossRef] [Green Version]
- Na, B.K.; Shenai, B.R.; Sijwali, P.S.; Choe, Y.; Pandey, K.C.; Singh, A.; Craik, C.S.; Rosenthal, P.J. Identification and biochemical characterization of vivapains, cysteine proteases of the malaria parasite Plasmodium vivax. Biochem. J. 2004, 378, 529–538. [Google Scholar] [CrossRef]
- Na, B.K.; Bae, Y.A.; Zo, Y.G.; Choe, Y.; Kim, S.H.; Desai, P.V.; Avery, M.A.; Craik, C.S.; Kim, T.S.; Rosenthal, P.J.; et al. Biochemical properties of a novel cysteine protease of Plasmodium vivax, vivapain-4. PLoS Negl. Trop. Dis. 2010, 4, e849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prasad, R.; Atul; Soni, A.; Puri, S.K.; Sijwali, P.S. Expression, characterization, and cellular localization of knowpains, papain-like cysteine proteases of the Plasmodium knowlesi malaria parasite. PLoS ONE 2012, 7, e51619. [Google Scholar] [CrossRef] [PubMed]
- Lê, H.G.; Kang, J.M.; Võ, T.C.; Yoo, W.G.; Lee, K.H.; Na, B.K. Biochemical properties of two Plasmodium Malariae cysteine proteases, malapain-2 and malapain-4. Microorganisms 2022, 10, 193. [Google Scholar] [CrossRef] [PubMed]
- Pandey, K.C.; Singh, N.; Arastu-Kapur, S.; Bogyo, M.; Rosenthal, P.J. Falstatin, a cysteine protease inhibitor of Plasmodium falciparum, Facilitates Erythrocyte Invasion. PLoS Pathog. 2006, 2, e117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rennenberg, A.; Lehmann, C.; Heitmann, A.; Witt, T.; Hansen, G.; Nagarajan, K.; Deschermeier, C.; Turk, V.; Hilgenfeld, R.; Heussler, V.T. Exoerythrocytic Plasmodium parasites secrete a cysteine protease inhibitor involved in sporozoite invasion and capable of blocking cell death of host hepatocytes. PLoS Pathog. 2010, 6, e1000825. [Google Scholar] [CrossRef] [Green Version]
- Pei, Y.; Miller, J.L.; Lindner, S.E.; Vaughan, A.M.; Torii, M.; Kappe, S.H.I. Plasmodium yoelii inhibitor of cysteine proteases is exported to exomembrane structures and interacts with yoelipain-2 during asexual blood-stage development. Cell. Microbiol. 2013, 15, 1508–1526. [Google Scholar] [CrossRef] [Green Version]
- Hansen, G.; Heitmann, A.; Witt, T.; Li, H.; Jiang, H.; Shen, X.; Heussler, V.T.; Rennenberg, A.; Hilgenfeld, R. Structural basis for the regulation of cysteine-protease activity by a new class of protease inhibitors in Plasmodium. Structure 2011, 19, 919–929. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.X.; Pandey, K.C.; Scharfstein, J.; Whisstock, J.; Huang, R.K.; Jacobelli, J.; Fletterick, R.J.; Rosenthal, P.J.; Abrahamson, M.; Brinen, L.S.; et al. The structure of chagasin in complex with a cysteine protease clarifies the binding mode and evolution of an inhibitor family. Structure 2007, 15, 535–543. [Google Scholar] [CrossRef] [Green Version]
- Kang, J.M.; Ju, H.L.; Yu, J.R.; Sohn, W.M.; Na, B.K. Cryptostatin, a chagasin-family cysteine protease inhibitor of Cryptosporidium parvum. Parasitology 2012, 139, 1029–1037. [Google Scholar] [CrossRef]
- Kang, J.M.; Lee, K.H.; Sohn, W.M.; Na, B.K. identification and functional characterization of csstefin-1, a cysteine protease inhibitor of Clonorchis sinensis. Mol. Biochem. Parasitol. 2011, 177, 126–134. [Google Scholar] [CrossRef]
- Thaí, T.L.; Kang, J.M.; Lê, H.G.; Lee, J.; Yoo, W.G.; Shin, H.J.; Sohn, W.M.; Na, B.K. Fowlerstefin, a cysteine protease inhibitor of Naegleria fowleri, induces inflammatory responses in BV-2 microglial cells in vitro. Parasites Vectors 2020, 13, 41. [Google Scholar] [CrossRef] [PubMed]
- Boysen, K.E.; Matuschewski, K. Inhibitor of cysteine proteases is critical for motility and infectivity of Plasmodium sporozoites. MBio 2013, 4, e00874-13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehmann, C.; Heitmann, A.; Mishra, S.; Burda, P.C.; Singer, M.; Prado, M.; Niklaus, L.; Lacroix, C.; Ménard, R.; Frischknecht, F.; et al. A cysteine protease inhibitor of Plasmodium berghei is essential for exo-erythrocytic development. PLoS Pathog. 2014, 10, e1004336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, J.M.; Cho, P.Y.; Moe, M.; Lee, J.; Jun, H.; Lee, H.W.; Ahn, S.K.; Kim, T.I.; Pak, J.H.; Myint, M.K.; et al. Comparison of the diagnostic performance of microscopic examination with nested polymerase chain reaction for optimum malaria diagnosis in Upper Myanmar. Malar. J. 2017, 16, 119. [Google Scholar] [CrossRef] [Green Version]
- Lê, H.G.; Ham, A.J.; Kang, J.M.; Võ, T.C.; Naw, H.; Sohn, H.J.; Shin, H.J.; Na, B.K. A novel cysteine protease inhibitor of Naegleria fowleri that is specifically expressed during encystation and at mature cysts. Pathogens 2021, 10, e1004336. [Google Scholar] [CrossRef]
- Sijwali, P.S.; Shenai, B.R.; Gut, J.; Singh, A.; Rosenthal, P.J. Expression and characterization of the Plasmodium falciparum haemoglobinase Falcipain-3. Biochem. J. 2001, 360, 481–489. [Google Scholar] [CrossRef]
- Kang, J.M.; Ju, H.L.; Lee, K.H.; Kim, T.S.; Pak, J.H.; Sohn, W.M.; Na, B.K. Identification and characterization of the second cysteine protease inhibitor of Clonorchis sinensis (CsStefin-2). Parasitol. Res. 2014, 113, 47–58. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lê, H.G.; Kang, J.-M.; Võ, T.C.; Nguyễn, T.D.; Jung, M.; Shin, M.K.; Yoo, W.G.; Na, B.-K. Inhibitor of Cysteine Protease of Plasmodium malariae Regulates Malapains, Endogenous Cysteine Proteases of the Parasite. Pathogens 2022, 11, 605. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11050605
Lê HG, Kang J-M, Võ TC, Nguyễn TD, Jung M, Shin MK, Yoo WG, Na B-K. Inhibitor of Cysteine Protease of Plasmodium malariae Regulates Malapains, Endogenous Cysteine Proteases of the Parasite. Pathogens. 2022; 11(5):605. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11050605
Chicago/Turabian StyleLê, Hương Giang, Jung-Mi Kang, Tuấn Cường Võ, Thảo Dương Nguyễn, Myunghwan Jung, Min Kyoung Shin, Won Gi Yoo, and Byoung-Kuk Na. 2022. "Inhibitor of Cysteine Protease of Plasmodium malariae Regulates Malapains, Endogenous Cysteine Proteases of the Parasite" Pathogens 11, no. 5: 605. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11050605