
Species Diversity, Habitat Distribution, and Blood Meal Analysis of Haematophagous Dipterans Collected by CDC-UV Light Traps in the Dominican Republic

 ,
,
Abstract
:1. Introduction
2. Results
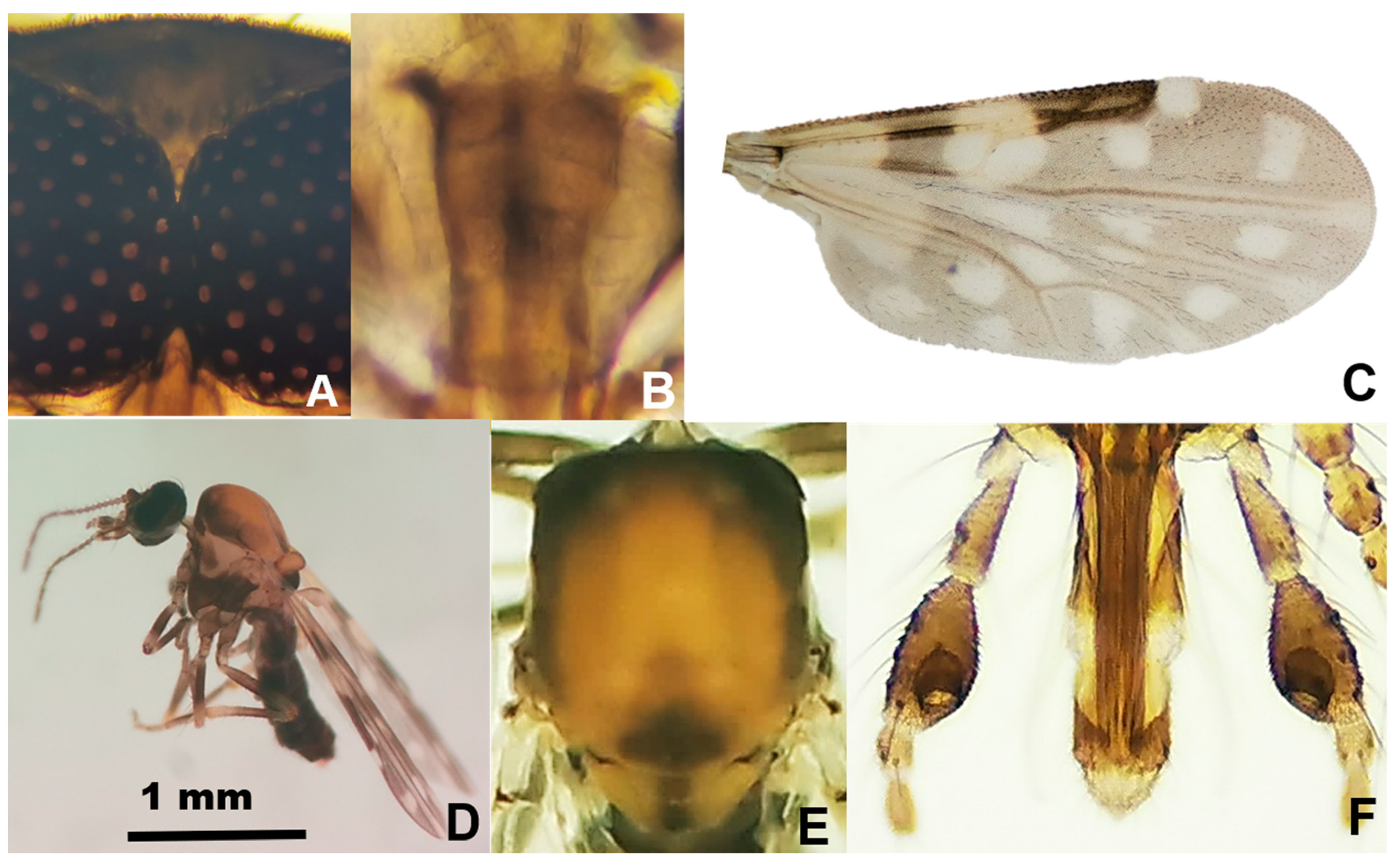
2.1. Species Composition Based on Morphological Features
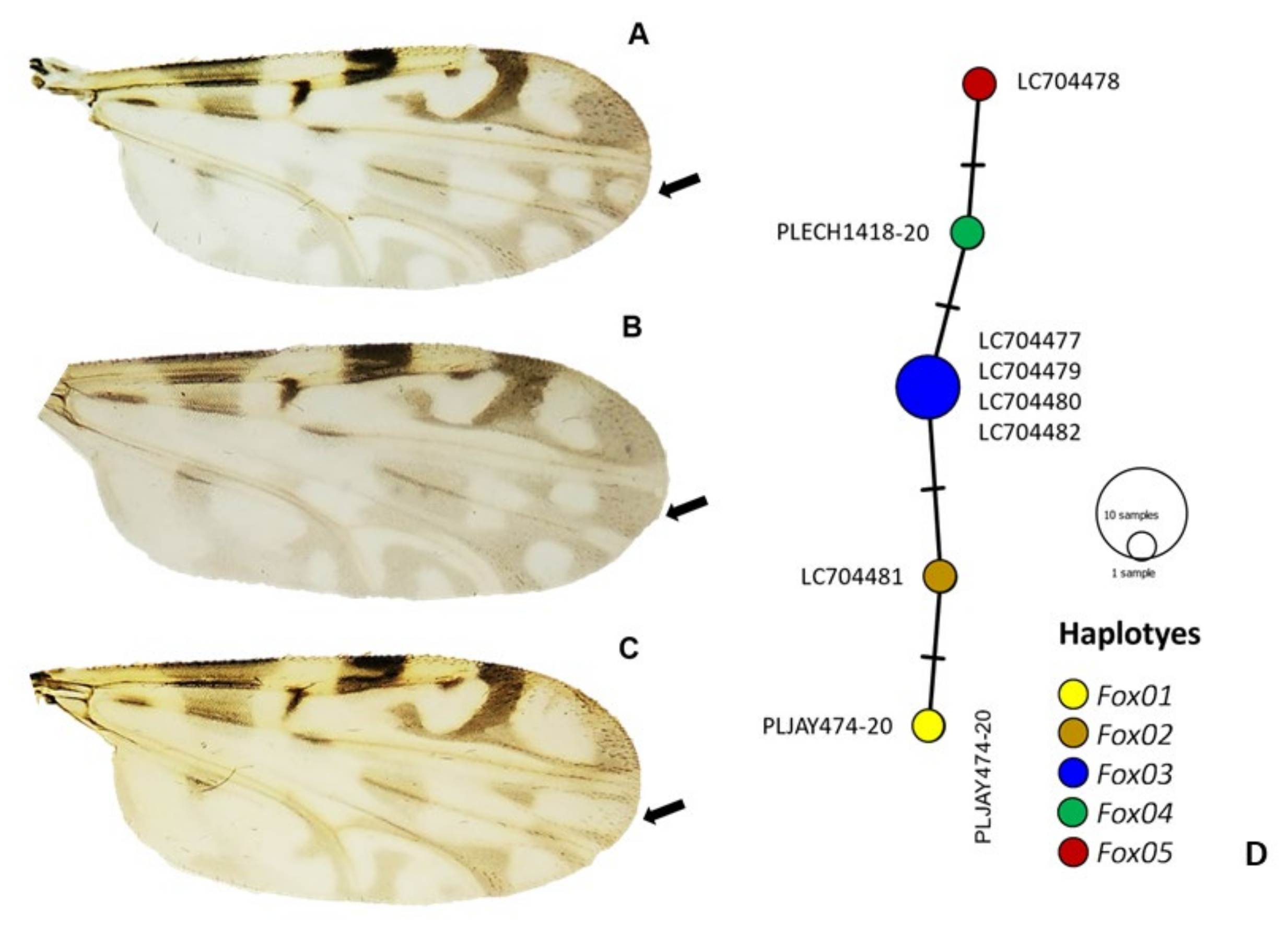
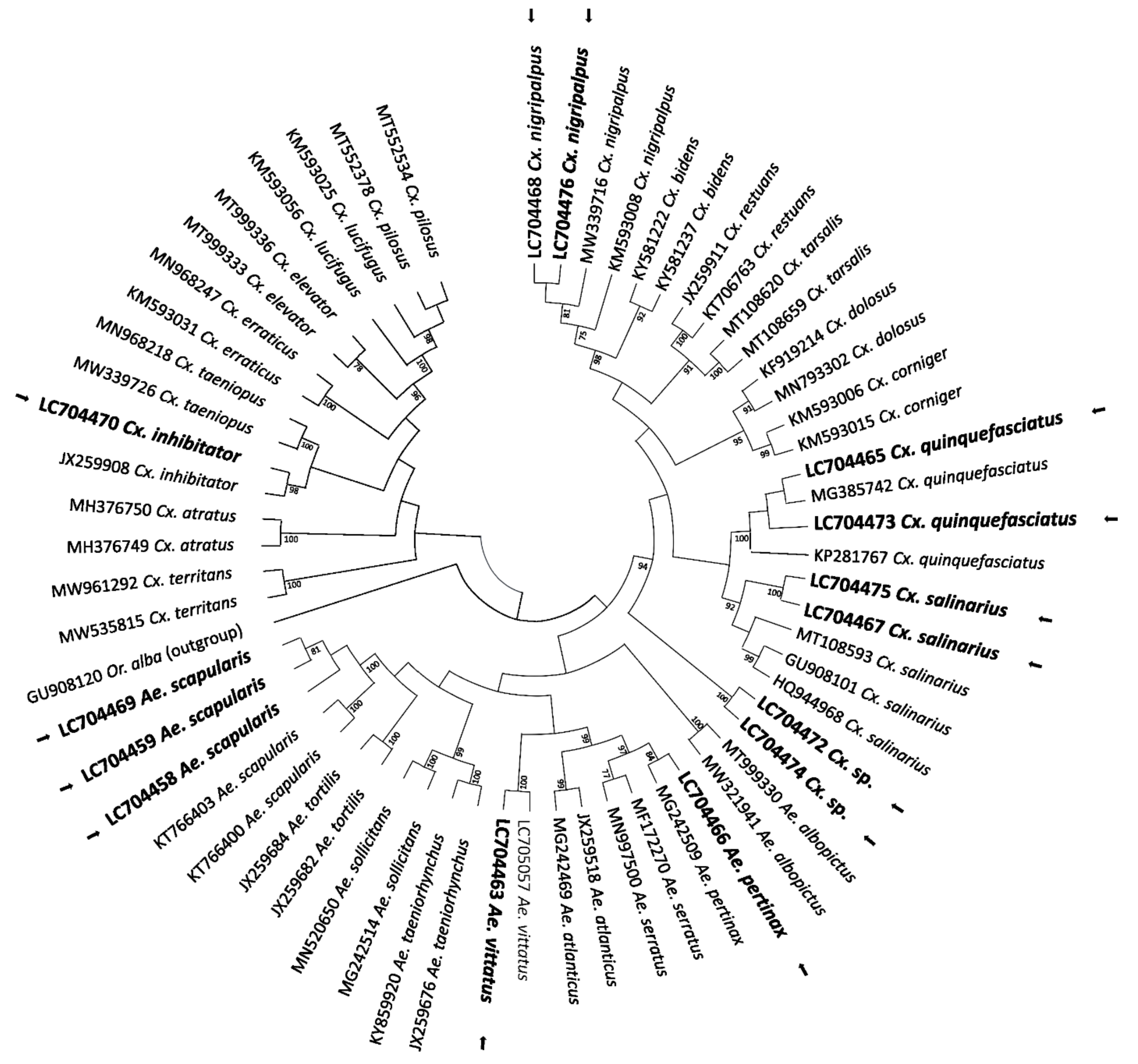
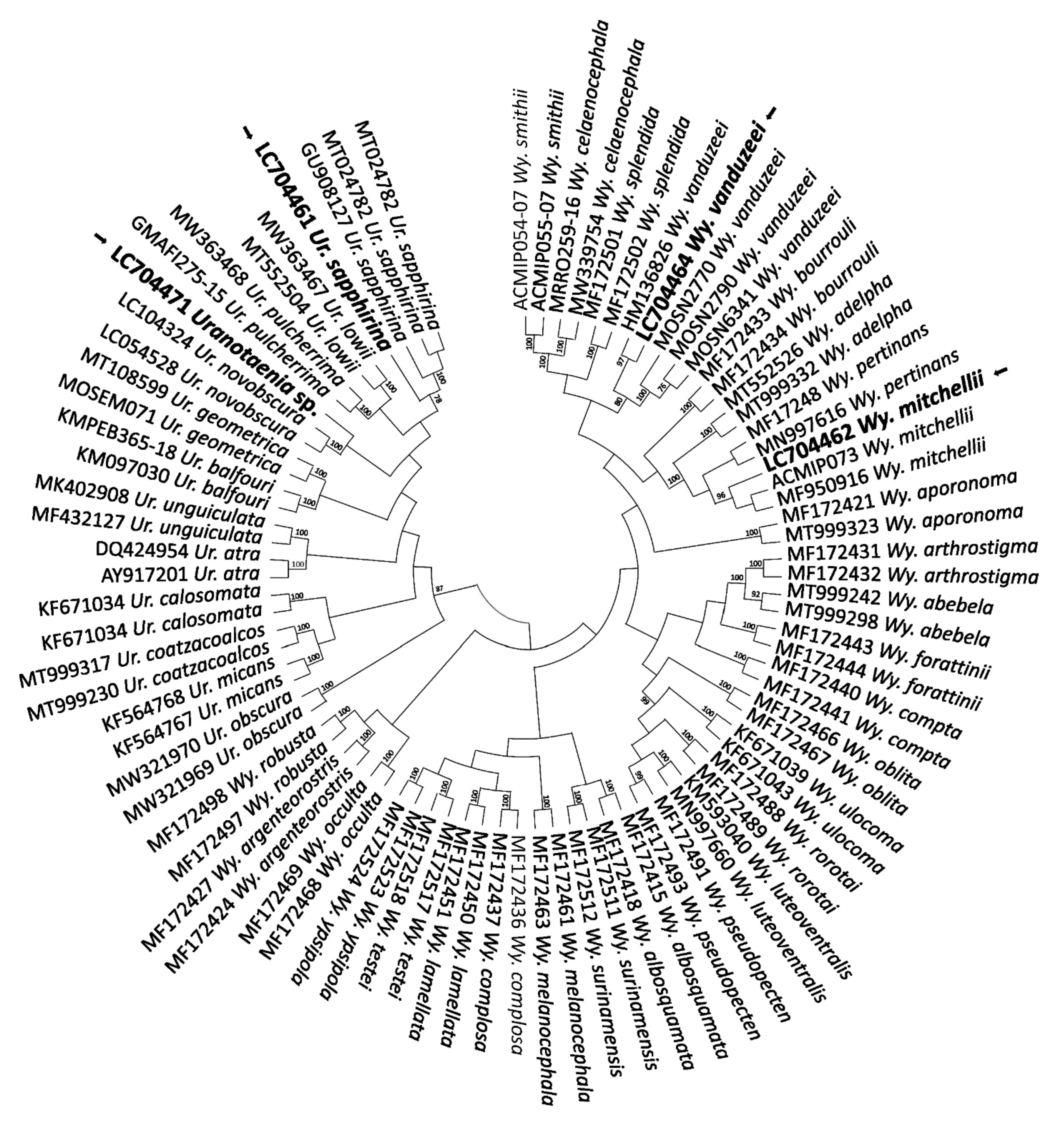
2.2. COI-Based DNA Barcodes and Phylogenetic Analysis
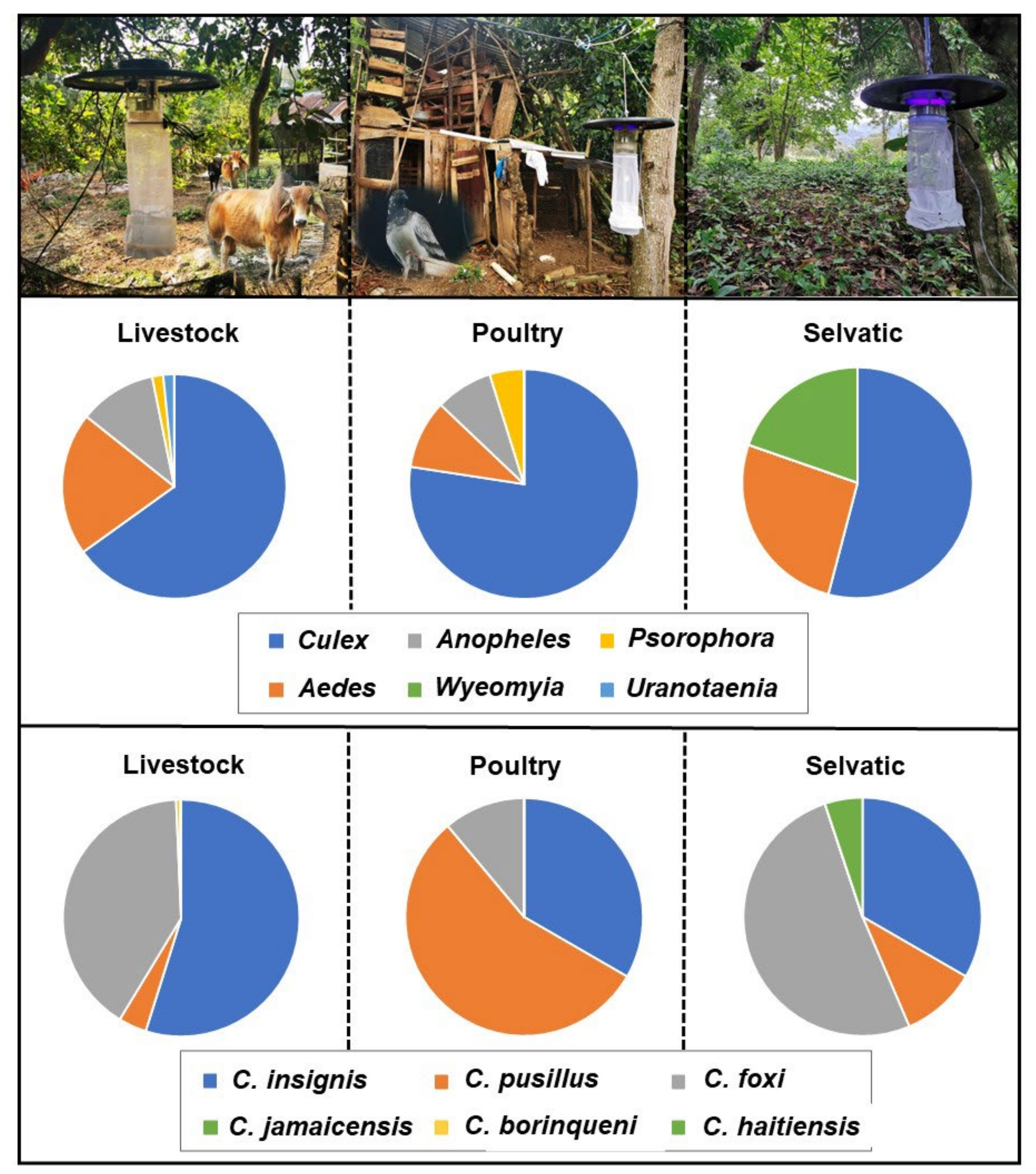
2.3. Spatial Distribution
2.4. Blood Meal Analysis
3. Discussion
4. Materials and Methods
4.1. Study Area
4.2. Entomological Survey
4.3. Morphological Species Identification
4.4. Molecular Species Identification
4.5. Identification of Host Blood Meal Sources
4.6. Analysis of Sequences and Statistical Analysis
4.7. Evolutionary Analysis by the Maximum Likelihood Method
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alarcón-Elbal, P.M.; Paulino-Ramírez, R.; Diéguez-Fernández, L.; Fimia-Duarte, R.; Guerrero, K.A.; González, M. Arbovirosis transmitidas por mosquitos (Diptera: Culicidae) en la República Dominicana: Una revisión. Biologist 2017, 1, 193–219. [Google Scholar] [CrossRef] [Green Version]
- Marcondes, C.B.; Contigiani, M.; Gleiser, R.M.; Reisen, W. Emergent and reemergent arboviruses in South America and the Caribbean: Why so many and why now? J. Med. Entomol. 2017, 54, 509–532. [Google Scholar] [CrossRef] [PubMed]
- Sosa, M.A.R.; Rueda, J.; Bautista, Y.E.V.; Fimia-Duarte, R.; de Prada, M.B.; Guerrero, K.A.; Alarcón-Elbal, P.M. Diversidad de mosquitos (Diptera: Culicidae) de Jarabacoa, República Dominicana. Graellsia 2019, 75, e084. [Google Scholar]
- Rodríguez Sosa, M.A.; Rueda, J.; Pichardo Rodríguez, R.J.; Vásquez Bautista, Y.E.; Durán Tiburcio, J.C.; Fimia-Duarte, R.; Alarcón-Elbal, P.M. Primera cita de Culex interrogator (Diptera: Culicidae) para La Hispaniola y actualización del listado de mosquitos de Jarabacoa, República Dominicana. Novit. Caribaea 2020, 16, 110–121. [Google Scholar] [CrossRef]
- Becker, N.; Petrić, D.; Zgomba, M.; Boase, C.; Madon, M.B.; Dahl, C.; Kaiser, A. Mosquitoes. Identification, Ecology and Control, 3rd ed.; Springer International Publishing: Cham, Switzerland, 2020. [Google Scholar]
- Francis, S.; Frank, C.; Buchanan, L.; Green, S.; Stennett-Brown, R.; Gordon-Strachan, G.; Rubio-Palis, Y.; Grant, C.; Alexander-Lindo, R.L.; Nwokocha, C.; et al. Challenges in the control of neglected insect vector diseases of human importance in the Anglo-Caribbean. One Health 2021, 13, 100316. [Google Scholar] [CrossRef]
- Diéguez Fernández, L.; Monzón Muñoz, M.V.; Juárez Sandoval, J.A.; Barrios Barrios, D.Y.; Barrientos Juárez, M.E.; Barrios, Y.H.; Rodríguez-Sosa, M.A.; Vásquez-Bautista, Y.E.; Alarcón-Elbal, P.M. Caracterización de la infestación de viviendas por tres especies de mosquitos (Diptera: Culicidae) de importancia médica en países de bajos y medios ingresos: Recomendaciones para su control doméstico. Ciencia y Salud 2021, 5, 49–66. [Google Scholar] [CrossRef]
- Sudeep, A.B.; Shil, P. Aedes vittatus (Bigot) Mosquito: An emerging threat to hublic Health. J. Vector Borne Dis. 2017, 54, 295–300. [Google Scholar] [CrossRef]
- Díaz-Martínez, I.; Diéguez-Fernández, L.; Santana-Águila, B.; de la Paz, E.M.A.; Ruiz-Domínguez, D.; Alarcón-Elbal, P.M. New introduction of Aedes vittatus (Diptera: Culicidae) into the East-Central region of Cuba: Ecological characterization and medical relevance. Interam. J. Med. Health 2021, 4, 1–6. [Google Scholar]
- Alarcón-Elbal, P.M.; Rodríguez-Sosa, M.A.; Newman, B.C.; Sutton, W.B. The First record of Aedes vittatus (Diptera: Culicidae) in the Dominican Republic: Public health implications of a potential invasive mosquito species in the Americas. J. Med. Entomol. 2020, 57, 2016–2021. [Google Scholar] [CrossRef]
- Raccurt, C.P. Mansonella ozzardi and Its Vectors in the New World: An update with emphasis on the current situation in Haiti. J. Helminthol. 2018, 92, 655–661. [Google Scholar] [CrossRef]
- Lowrie, R.C.; Raccurt, C.P. Assessment of Culicoides barbosai as a vector of Mansonella ozzardi in Haiti. Am. J. Trop. Med. Hyg. 1984, 33, 1275–1277. [Google Scholar] [CrossRef]
- Raccurt, C.P.; Brasseur, P.; Boncy, J. Mansonelliasis, a neglected parasitic disease in Haiti. Mem. Inst. Oswaldo Cruz. 2014, 109, 709–711. [Google Scholar] [CrossRef] [Green Version]
- Elbadry, M.A.; Durães-Carvalho, R.; Blohm, G.M.; Stephenson, C.J.; Loeb, J.C.; White, S.K.; Telisma, T.; Chavannes, S.; Beau De Rochars, V.M.; Salemi, M.; et al. Orthobunyaviruses in the Caribbean: Melao and Oropouche virus infections in school children in Haiti in 2014. PLoS Negl. Trop. Dis. 2021, 15, e009494. [Google Scholar] [CrossRef]
- Thompson, L.H.; Mo, C.L.; Oviedo, M.T.; Homan, E.J. Prevalence and Incidence of Bluetongue Viruses in the Caribbean Basin: Serologic and Virologic Findings. In Proceedings of the Second International Symposium, Paris, France, 17–21 June 1992; Walton, T.E., Osburn, B.I., Eds.; CRC Press Inc.: Boca Raton, FL, USA; pp. 106–113. [Google Scholar]
- Garcia-Rejon, J.E.; Blitvich, B.J.; Farfan-Ale, J.A.; Loroño-Pino, M.A.; Chi Chim, W.A.; Flores-Flores, L.F.; Rosado-Paredes, E.; Baak-Baak, C.; Perez-Mutul, J.; Suarez-Solis, V.; et al. Host-feeding preference of the mosquito, Culex quinquefasciatus, in Yucatan State, Mexico. J. Insect Sci. 2010, 10, 32. [Google Scholar] [CrossRef] [Green Version]
- Kent, R.J. Molecular methods for arthropod bloodmeal identification and applications to ecological and vector-borne disease studies. Mol. Ecol. Resour. 2009, 9, 4–18. [Google Scholar] [CrossRef]
- Alarcón-Elbal, P.M.; Sandiford, S.L. One health entomology in the Insular Caribbean: Time to bet on prevention. Interam. J. Med. Health 2021, 4, e202101001. [Google Scholar] [CrossRef]
- Mitchell, C.J.; Taylor, S.A.; Christensen, H.A. Identification of blood meals from engorged dipterans collected in the Dominican Republic during an Eastern Equine encephalitis outbreak. J. Med. Entomol. 1983, 20, 686–687. [Google Scholar] [CrossRef]
- Rodríguez-Sosa, M.A.; Vásquez-Bautista, Y.E.; Fimia-Duarte, R.; Guerrero, K.A.; Alarcón-Elbal, P.M.; Rodríguez-Sosa, M.A.; Vásquez-Bautista, Y.E.; Fimia-Duarte, R.; Guerrero, K.A.; Alarcón-Elbal, P.M. Primer registro de Uranotaenia cooki (Diptera: Culicidae) para República Dominicana. Rev. Colomb. Entomol. 2018, 44, 141–144. [Google Scholar] [CrossRef]
- Rueda, J.; Rodríguez-sosa, M.A.; Vásquez-bautista, Y.E.; Guerrero, K.A. Primera cita de Culex (Culex) garciai González Broche, 2000 (Diptera: Culicidae) para La Española. An. Biol. 2018, 40, 95–101. [Google Scholar] [CrossRef]
- Bisset Lazcano, J.A.; Marquetti Fernández, M.C.; Rodriguez Coto, M.M. Contribución de estudios entomológicos sobre Aedes aegypti y Aedes albopictus. Retrospectiva y retos para su control en Cuba. Rev. Cuba. Med. Trop. 2017, 69, 1–20. [Google Scholar]
- Jinbo, U.; Kato, T.; Ito, M. Current progress in DNA barcoding and future implications for entomology. Entomol. Sci. 2011, 14, 107–124. [Google Scholar] [CrossRef]
- Chan, A.; Chiang, L.P.; Hapuarachchi, H.C.; Tan, C.H.; Pang, S.C.; Lee, R.; Lee, K.S.; Ng, L.C.; Lam-Phua, S.G. DNA Barcoding: Complementing morphological identification of mosquito species in Singapore. Parasit. Vectors 2014, 7, 569. [Google Scholar] [CrossRef]
- Buhay, J.E. “COI-like” sequences are becoming problematic in molecular systematic and DNA barcodeng studies. J. Crustac. Biol. 2009, 29, 96–110. [Google Scholar] [CrossRef]
- Weeraratne, T.C.; Surendran, S.N.; Parakrama Karunaratne, S.H.P. DNA barcoding of morphologically characterized mosquitoes belonging to the subfamily Culicinae from Sri Lanka. Parasit. Vectors 2018, 11, 266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCreless, E.E.; Huff, D.D.; Croll, D.A.; Tershy, B.R.; Spatz, D.R.; Holmes, N.D.; Butchart, S.H.M.; Wilcox, C. Past and estimated future impact of invasive alien mammals on insular threatened vertebrate populations. Nat. Commun. 2016, 7, 12488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russell, J.C.; Meyer, J.Y.; Holmes, N.D.; Pagad, S. Invasive alien species on Islands: Impacts, distribution, interactions and management. Environ. Conserv. 2017, 44, 359–370. [Google Scholar] [CrossRef] [Green Version]
- Thurman, E.B. Report of the initial operation of a light trap in northern Thailand. Mosq. News 1955, 15, 215–224. [Google Scholar]
- Walter Reed Biosystematics Unit (WRBU). 2021. Culex salinarius Coquillett. 1904. Available online: https://wrbu.si.edu/vectorspecies/mosquitoes/salinarius (accessed on 10 May 2022).
- Sardelis, M.R.; Turell, M.J.; Dohm, D.J.; O’Guinn, M.L. Vector competence of selected north american Culex and Coquillettidia mosquitoes for West Nile virus. Emerg. Infect. Dis. 2001, 7, 1018–1022. [Google Scholar] [CrossRef]
- Diéguez-Fernández, L.; Rodríguez-Sosa, M.A.; Vásquez-Bautista, Y.E.; Borge-de Prada, M.; Alarcón-Elbal, P.M. Aportes a la bioecología de Culex quinquefasciatus (Diptera: Culicidae), vector de encefalitis, en Jarabacoa, República Dominicana. Folia Entomol. Mex. 2020, 6, 26–32. [Google Scholar]
- Raccurt, C.P.; Rebholtz, C.; Delécolle, J.C. Culicoides haitiensis et Culicoides borinqueni en Haïti. 2. Notes Écologiques et Chorologiques [Dipt. Ceratopogonidae]. Bull. Société Entomol. Fr. 1996, 91, 125–130. [Google Scholar] [CrossRef]
- Delecolle, J.C.; Raccurt, C.P.; Rebholtz, C. Culicoides haitiensis n. sp. et C. borinqueni Fox et Hoffman en Haiti. Etude morphologique (Diptera, Ceratopogonidae). Nouv. Rev. Entomol. 1986, 3, 107–115. [Google Scholar]
- Perez-Gelabert, D.E. Checklist, bibliography and quantitative data of the arthropods of Hispaniola. Zootaxa 2020, 4749, zootaxa.4749.1.1. [Google Scholar] [CrossRef]
- Meiswinkel, R.; Labuschagne, K.; Goffredo, M. Christopher. Columbus and Culicoides: Was C. jamaicensis Edwards, 1922 introduced into the Mediterranean 500 years ago and later re-named C. paolae Boorman 1996? Vet. Ital. 2004, 40, 340–344. [Google Scholar]
- Aguilar-Vega, C.; Rivera, B.; Lucientes, J.; Gutiérrez-Boada, I.; Sánchez-Vizcaíno, J.M. A Study of the composition of the Obsoletus complex and genetic diversity of Culicoides obsoletus populations in Spain. Parasit. Vectors 2021, 14, 1–13. [Google Scholar] [CrossRef]
- Jansen, C.C.; Webb, C.E.; Graham, G.C.; Craig, S.B.; Zborowski, P.; Ritchie, S.A.; Russell, R.C.; Van Den Hurk, A.F. Blood sources of mosquitoes collected from urban and peri-urban environments in Eastern Australia with species-specific molecular analysis of avian blood meals. Am. J. Trop. Med. Hyg. 2009, 81, 849–857. [Google Scholar] [CrossRef] [Green Version]
- Fitzpatrick, D.M.; Hattaway, L.M.; Hsueh, A.N.; Ramos-Niño, M.E.; Cheetham, S.M. PCR-based bloodmeal analysis of Aedes aegypti and Culex quinquefasciatus (Diptera: Culicidae) in St. George Parish, Grenada. J. Med. Entomol. 2009, 56, 1170–1175. [Google Scholar] [CrossRef] [Green Version]
- Cebrián-Camisón, S.; de la Puente, J.M.; Figuerola, J. A literature review of host feeding patterns of invasive Aedes mosquitoes in Europe. Insects 2020, 11, 848. [Google Scholar] [CrossRef]
- Santos, C.S.; Pie, M.R.; da Rocha, T.C.; Navarro-Silva, M.A. Molecular identification of blood meals in mosquitoes (Diptera, Culicidae) in urban and forested habitats in southern Brazil. PLoS ONE 2019, 14, e0212517. [Google Scholar] [CrossRef]
- Martínez-de la Puente, J.; Figuerola, J.; Soriguer, R. Fur or feather? Feeding preferences of species of Culicoides biting midges in Europe. Trends Parasitol. 2015, 31, 16–22. [Google Scholar] [CrossRef] [Green Version]
- McGregor, B.L.; Stenn, T.; Sayler, K.A.; Blosser, E.M.; Blackburn, J.K.; Wisely, S.M.; Burkett-Cadena, N.D. Host use patterns of Culicoides spp. biting midges at a big game preserve in Florida, U.S.A., and implications for the transmission of Orbiviruses. Med. Vet. Entomol. 2019, 33, 110–120. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Saenz, A.; Mccarter, P.; Baylis, M. The influence of host number on the attraction of biting midges, Culicoides spp., to light traps. Med. Vet. Entomol. 2011, 25, 113–115. [Google Scholar] [CrossRef]
- Davies, J.F. The distribution of sandflies (Culicoides spp.) Breeding in a tidal mangrove swamp in Jamaica and the effect of tides on the emergence of C. furens Poey and C. barbosai (Wirth & Blanton). West Indian Med. J. 1967, 16, 39–50. [Google Scholar] [PubMed]
- Mo, C.L.; Thompson, L.H.; Homan, E.J.; Oviedo, M.T.; Greiner, E.C.; González, J.; Sáenz, M.R. Bluetongue virus isolations from vectors and ruminants in Central America and the Caribbean. Interamerican Bluetongue Team. Am. J. Vet. Res. 1994, 55, 211–215. [Google Scholar] [PubMed]
- Tanya, V.N.; Greiner, E.C.; Gibbs, E.P.J. Evaluation of Culicoides insignis (Diptera: Ceratopogonidae) as a vector of Bluetongue virus. Vet. Microbiol. 1992, 32, 1–14. [Google Scholar] [CrossRef]
- Rios, R.R.S.; Santarém, M.C.A.; Ribeiro Júnior, K.A.L.; de Melo, B.A.; da Silva, S.G.M.; da Silva, N.C.; Dos Santos, V.R.V.; Dos Santos, J.M.; Santana, A.E.G.; Fraga, A.B. Culicoides insignis Lutz, 1913 (Diptera: Ceratopogonidae) biting midges in Northeast of Brazil. Insects 2021, 12, 366. [Google Scholar] [CrossRef]
- Bandeira, M.C.A.; Da Penha, A.; Moraes, J.L.P.; Brito, G.A.; Rebêlo, J.M.M. Infestation of brazilian peridomiciliary areas by Culicoides (Diptera: Ceratopogonidae) in humid and semihumid climates. J. Med. Entomol. 2016, 53, 1163–1168. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, L.P.C.; Silva, F.S. Seasonal abundance of livestock-associated Culicoides species in northeastern Brazil. Med. Vet. Entomol. 2014, 28, 228–231. [Google Scholar] [CrossRef]
- Costa, J.C.; Lorosa, E.S.; Moraes, J.L.P.; Rebêlo, J.M.M. Espécies de Culicoides (Diptera; Ceratopogonidae) e hospedeiros potenciais em área de ecoturismo do Parque Nacional dos Lençóis Maranhenses, Brasil. Rev. Pan-Amazônica Saúde 2013, 4, 11–18. [Google Scholar] [CrossRef]
- González, M.A.; Dilger, E.; Ronderos, M.M.; Spinelli, G.R.; Courtenay, O.; Hamilton, J.G.C. Significant reduction in abundance of peridomestic mosquitoes (Culicidae) and Culicoides Midges (Ceratopogonidae) after chemical intervention in Western São Paulo, Brazil. Parasit. Vectors 2020, 13, 549. [Google Scholar] [CrossRef]
- González, M.A.; Ruiz-Arrondo, I.; Rodríguez-Sosa, M.A.; Rueda, J.; Oteo, J.A.; Alarcón-Elbal, P.M. New insights on black Flies (Diptera: Simuliidae) from a touristic destination of Hispaniola. Int. J. Trop. Insect Sci. 2022, 42, 2281–2295. [Google Scholar] [CrossRef]
- Rodríguez-Pérez, M.A.; Adeleke, M.A.; Burkett-Cadena, N.D.; Garza-Hernández, J.A.; Reyes-Villanueva, F.; Cupp, E.W.; Toé, L.; Salinas-Carmona, M.C.; Rodríguez-Ramírez, A.D.; Katholi, C.R.; et al. Development of a novel trap for the collection of black flies of the Simulium ochraceum complex. PLoS ONE 2013, 8, e76814. [Google Scholar] [CrossRef]
- Brito, L.G.; Regitano, L.C.d.A.; Huacca, M.E.F.; Carrilho, E.; Paes, M.J.; Moya-Borja, G.E. Genotype characterization of the Haematobia irritans (Diptera: Muscidae) from Brazil, Dominican Republic and Colombia based on randomly amplified polymorphic DNA (RAPD) analysis. Rev. Bras. Parasitol. Vet. 2008, 17, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Rojas, J.J.; Arque-Chunga, W.; Fernández-Salas, I.; Rebollar-Téllez, E.A. Comparative field evaluation of different traps for collecting adult Phlebotomine sand flies (Diptera: Psychodidae) in an endemic area of cutaneous leishmaniasis in Quintana Roo, Mexico. J. Am. Mosq. Control Assoc. 2016, 32, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.N. Phlebotominae sand Flies (Diptera: Psychodidae) and diffuse cutaneous leishmaniasis in the Dominican Republic. Ph.D. Thesis, University of Florida, Gainesville, FL, USA, 1984. [Google Scholar]
- Shaw, J.; Pratlong, F.; Floeter-Winter, L.; Ishikawa, E.; El Baidouri, F.; Ravel, C.; Dedet, J.P. Characterization of Leishmania (Leishmania) waltoni n. nsp. (Kinetoplastida: Trypanosomatidae), the parasite responsible for diffuse cutaneous leishmaniasis in the Dominican Republic. Am. J. Trop. Med. Hyg. 2015, 93, 552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarado, J.; Oddone, N.; Gil, L. Fortalecimiento de la Cadena de Valor de Turismo en Pedernales, República Dominicana. Publicación de las Naciones Unidas: CEPAL, México, 2017. Available online: https://www.cepal.org/es/publicaciones/41644-fortalecimiento-la-cadena-valor-turismo-pedernales-republica-dominicana (accessed on 22 October 2021).
- Beck, H.E.; Zimmermann, N.E.; McVicar, T.R.; Vergopolan, N.; Berg, A.; Wood, E.F. Present and Future Köppen-Geiger Climate Classification Maps at 1-Km Resolution. Sci. Data 2018, 5, 180214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wirth, W.W.; Dyce, A.L.; Spinelli, G. An atlas of wing photographs, with a summary of the numerical characters of the neotropical species of Culicoides (Diptera: Ceratopogonidae). Contrib. Am. Entomol. Inst. 1998, 22, 4. [Google Scholar]
- González Broche, R. Culícidos de Cuba; Editorial Científico Técnica: La Habana, Cuba, 2006. [Google Scholar]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Bravo-Barriga, D.; Parreira, R.; Maia, C.; Afonso, M.O.; Blanco-Ciudad, J.; Serrano, F.J.; Pérez-Martín, J.E.; Gómez-Gordo, L.; Campino, L.; Reina, D.; et al. Detection of Leishmania DNA and blood meal sources in Phlebotomine sand flies (Diptera: Psychodidae) in western of Spain: Update on distribution and risk factors associated. Acta Trop. 2016, 164, 414–424. [Google Scholar] [CrossRef]
- Barcode of Life Data System (BOLD). Available online: http://www.boldsystems.org/index.php/IDS_OpenIdEngine (accessed on 15 May 2022).
- Benson, D.A.; Cavanaugh, M.; Clark, K.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2013, 41, D36–D42. [Google Scholar] [CrossRef] [Green Version]
- DNA Data Bank of Japan (DDBJ). Available online: https://www.ddbj.nig.ac.jp/index-e.html (accessed on 15 May 2022).
- Siegel, S.; Castellan, N.J., Jr. Nonparametric Statistics for the Behavioral, Sciences, 2nd ed.; McGraw-Hill Humanities: New York, NY, USA, 1988; pp. 262–272. [Google Scholar]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics, Oxford University Press: New York, NY, USA, 2000.
- FigTree Version-v1.4.2. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 15 May 2022).
- Tamura, K. Estimation of the number of nucleotide substitutions when there are strong Transition-Transversion and G+C-Content Biases. Mol. Biol. Evol. 1992, 9, 678–687. [Google Scholar]
- Rozas, J.; Ferrer-Mata, A.; Sanchez-del Barrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sanchez-Gracia, A. DnaSP 6: DNA Sequence polymorphism analysis of large data sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Leigh, J.W.; Bryant, D. POPART: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diptera Species | Number Analysed | Number Amplified (%) 1 | Host |
|---|---|---|---|
| Culex quinquefasciatus | 1 | 1 (100) | Gallus gallus |
| Culex nigripalpus * | 1 | 1 (100) | Gallus gallus |
| Aedes albopictus | 1 | 1 (100) | Sus scrofa |
| Aedes scapularis | 1 | 1 (100) | Sus scrofa |
| Psorophora confinnis | 1 | 1 (100) | Sus scrofa |
| Culicoides insignis | 30 | 22 (73.3) | Bos taurus |
| Culicoides pusillus | 12 | 5 (41.6) | Bos taurus |
| Culicoides foxi * | 1 | 1 (100) | Bos taurus |
| Total | 48 | 33 (68.7) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
González, M.A.; Bravo-Barriga, D.; Rodríguez-Sosa, M.A.; Rueda, J.; Frontera, E.; Alarcón-Elbal, P.M. Species Diversity, Habitat Distribution, and Blood Meal Analysis of Haematophagous Dipterans Collected by CDC-UV Light Traps in the Dominican Republic. Pathogens 2022, 11, 714. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11070714
González MA, Bravo-Barriga D, Rodríguez-Sosa MA, Rueda J, Frontera E, Alarcón-Elbal PM. Species Diversity, Habitat Distribution, and Blood Meal Analysis of Haematophagous Dipterans Collected by CDC-UV Light Traps in the Dominican Republic. Pathogens. 2022; 11(7):714. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11070714
Chicago/Turabian StyleGonzález, Mikel Alexander, Daniel Bravo-Barriga, María Altagracia Rodríguez-Sosa, Juan Rueda, Eva Frontera, and Pedro María Alarcón-Elbal. 2022. "Species Diversity, Habitat Distribution, and Blood Meal Analysis of Haematophagous Dipterans Collected by CDC-UV Light Traps in the Dominican Republic" Pathogens 11, no. 7: 714. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11070714