
Fusarium Yellows of Ginger (Zingiber officinale Roscoe) Caused by Fusarium oxysporum f. sp. zingiberi Is Associated with Cultivar-Specific Expression of Defense-Responsive Genes
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ginger Cultivars
2.2. Plant Growth
2.3. Foz Isolates
2.4. DNA Extraction, PCR, and Phylogenetic Analysis
2.5. Plant Inoculation
2.6. Pathogenicity Trial
2.7. RNA Isolation and cDNA Synthesis
2.8. Selection of Candidate Genes, Primer Design, and Quantitative PCR Analysis
2.9. Primers for Gene Expression Analysis
2.10. Quantitative Real-Time PCR
3. Results
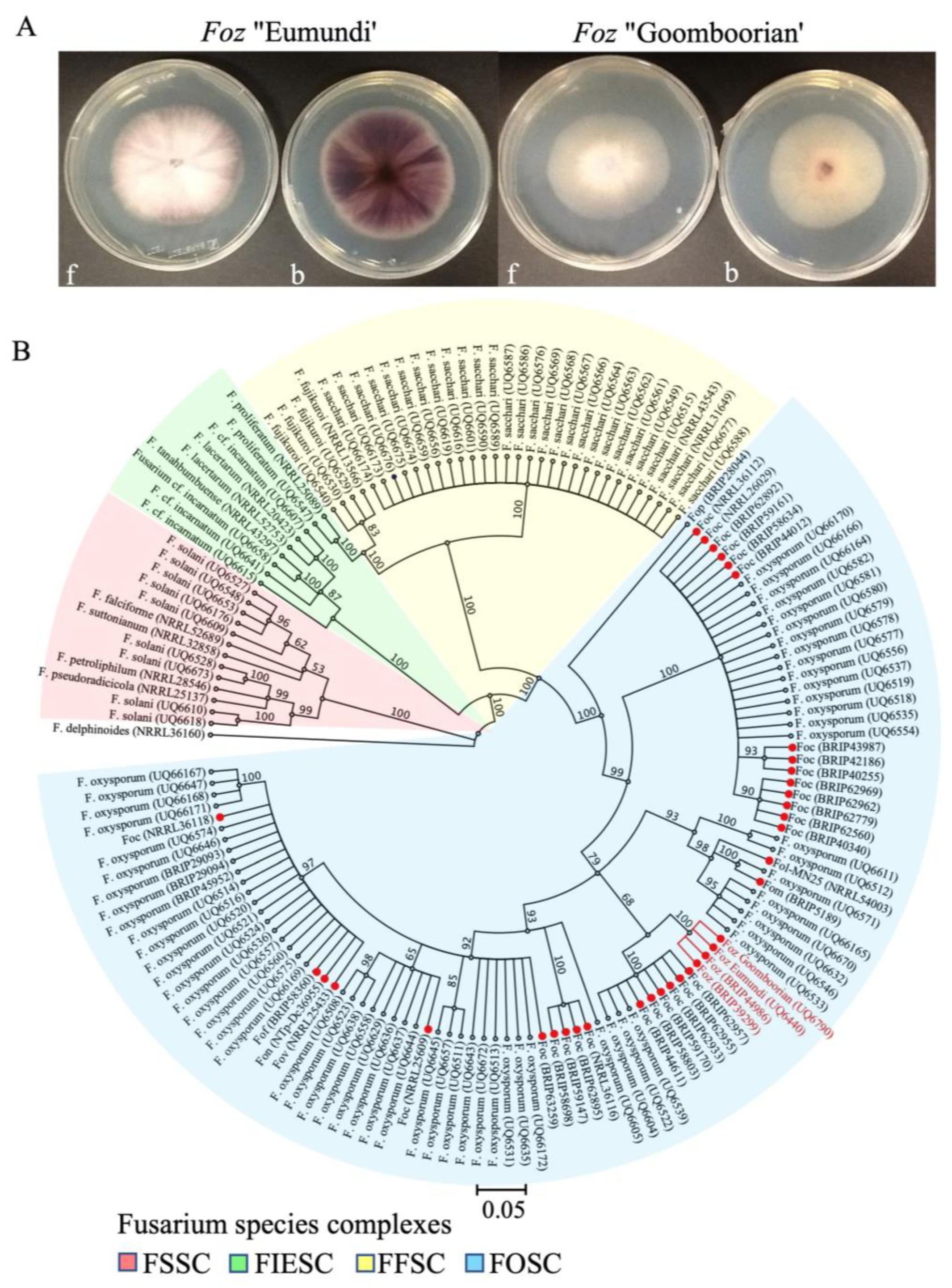
3.1. Morphology and Phylogenetic Analysis of Foz
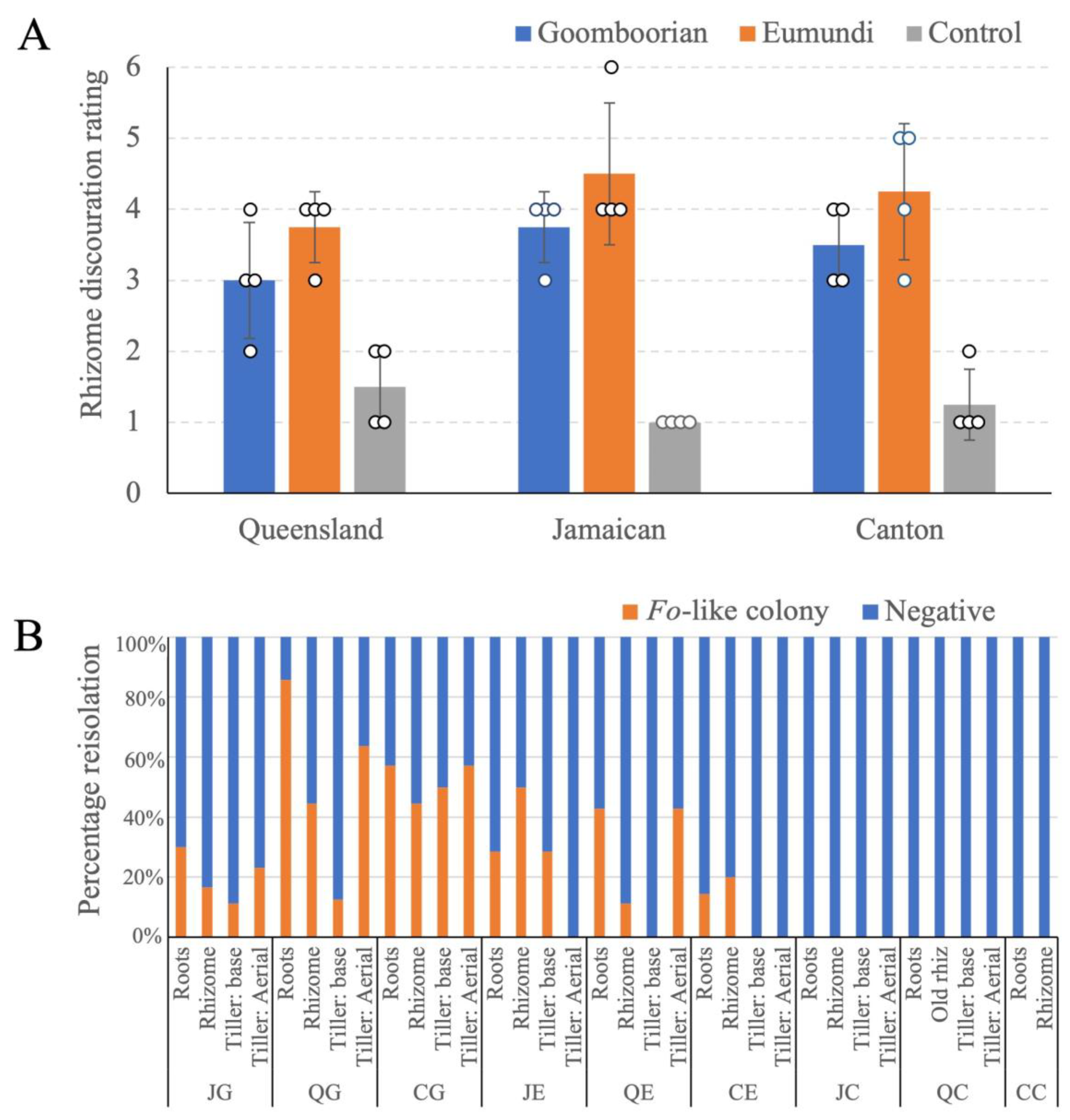
3.2. Pathogenicity Trial
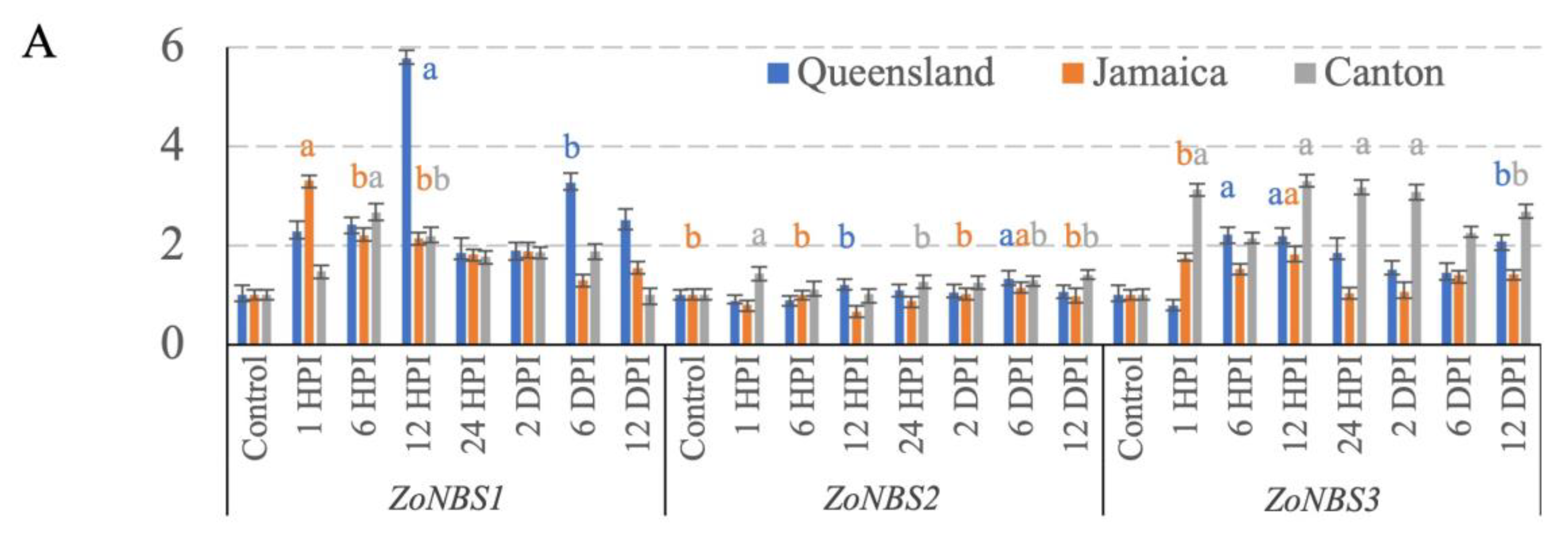
3.3. NBS-LRR Gene Expression
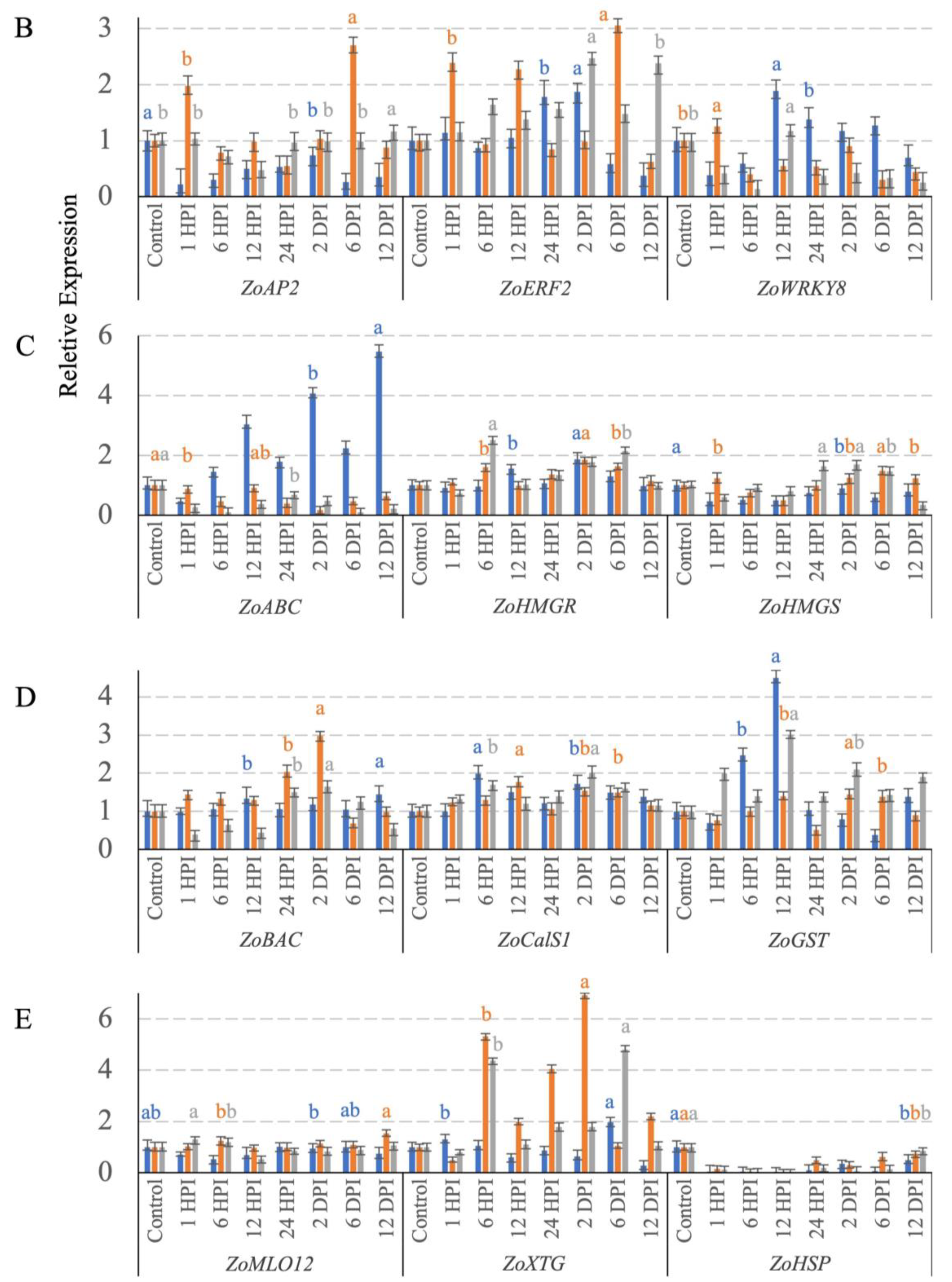
3.4. Transcription Factor Expression
3.5. Terpenoid-Synthesis-Related Gene Expression
3.6. Cell Wall Defense Related Gene Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grant, K.L.; Lutz, R.B. Ginger. Am. J. Health Syst. Pharm. 2000, 57, 945–947. [Google Scholar] [CrossRef] [PubMed]
- Parthasarathy, V.A.; Prasath, D. Cardamom. In Handbook of Herbs and Spices; Woodhead Publishing: Sawston, UK, 2012; pp. 131–170. [Google Scholar]
- Bhandari, U.; Sharma, J.N.; Zafar, R. The protective action of ethanolic ginger (Zingiber officinale) extract in cholesterol fed rabbits. J. Ethnopharmacol. 1998, 61, 167–171. [Google Scholar] [CrossRef] [PubMed]
- Fuhrman, B.; Rosenblat, M.; Hayek, T.; Coleman, R.; Aviram, M. Ginger extract consumption reduces plasma cholesterol, inhibits LDL oxidation and attenuates development of atherosclerosis in atherosclerotic, apolipoprotein E-deficient mice. J. Nutr. 2000, 130, 1124–1131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karunakaran, R.; Sadanandan, S.P. Zingiber officinale: Antiinflammatory actions and potential usage for arthritic conditions. In Bioactive Food as Dietary Interventions for Arthritis and Related Inflammatory Diseases, 2nd ed.; Watson, R.R., Preedy, V.R., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 233–244. [Google Scholar]
- Nicoll, R.; Henein, M.Y. Ginger (Zingiber officinale Roscoe): A hot remedy for cardiovascular disease? Int. J. Cardiol. 2007, 131, 408–409. [Google Scholar] [CrossRef] [PubMed]
- Shukla, Y.; Singh, M. Cancer preventive properties of ginger: A brief review. Food Chem. Toxicol. 2007, 45, 683–690. [Google Scholar] [CrossRef]
- Gaur, M.; Das, A.; Sahoo, R.K.; Kar, B.; Nayak, S.; Subudhi, E. De Novo transcriptome assembly of Zingiber officinale cv. Suruchi of Odisha. Genom. Data 2016, 9, 87–88. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Chi, L.D.; Mao, L.G.; Yan, D.D.; Wu, Z.F.; Ma, T.T.; Guo, M.X.; Wang, Q.X.; Ouyang, C.B.; Cao, A.C. First report of ginger rhizome rot caused by Fusarium oxysporum in China. Plant Dis. 2014, 98, 282. [Google Scholar] [CrossRef]
- Nair, K.P. The Agronomy and Economy of Ginger; Elsevier: Edinburgh, UK, 2013; pp. 225–292. [Google Scholar]
- Wohlmuth, H.; Leach, D.N.; Smith, M.K.; Myers, S.P. Gingerol content of diploid and tetraploid clones of ginger (Zingiber officinale Roscoe). J. Agric. Food Chem. 2005, 53, 5772–5778. [Google Scholar] [CrossRef]
- Kizhakkayil, J.; Sasikumar, B. Diversity, characterization and utilization of ginger: A review. Plant Genet. Resour. 2011, 9, 464–477. [Google Scholar] [CrossRef]
- Adaniya, S.; Shoda, M.; Fujieda, K. Effects of day length on flowering and rhizome swelling in ginger (Zingiber officinale Roscoe). J. Jpn. Soc. Hortic. Sci. 1989, 58, 649–656. [Google Scholar] [CrossRef]
- D’Hont, A.; Denoeud, F.; Aury, J.M.; Baurens, F.C.; Carreel, F.; Garsmeur, O.; Noel, B.; Bocs, S.; Droc, G.; Rouard, M.; et al. The banana (Musa acuminata) genome and the evolution of monocotyledonous plants. Nature 2012, 488, 213–217. [Google Scholar] [CrossRef] [Green Version]
- Mahdi, H.J.; Andayani, R.; Aziz, I. Determination of phylogenetic and molecular characteristics of three Malaysian ginger cultivars (Zingiber officinale Roscoe) using microsatellite DNA. Trop. Life Sci. Res. 2013, 24, 65–76. [Google Scholar]
- Prasath, D.; Karthika, R.; Habeeba, N.T.; Suraby, E.J.; Rosana, O.B.; Shaji, A.; Eapen, S.J.; Deshpande, U.; Anandaraj, M. Comparison of the transcriptomes of ginger (Zingiber officinale Rosc.) and mango ginger (Curcuma amada Roxb.) in response to the bacterial wilt infection. PLoS ONE 2014, 9, e99731. [Google Scholar] [CrossRef] [Green Version]
- Ravindran, P.N.; Babu, K.N.; Shiva, K. Botany and Crop Improvement of Ginger. In Ginger: The Genus Zingiber; Ravindran, P.N., Babu, K.N., Eds.; CRC Press: Boca Raton, FL, USA, 2016; pp. 15–85. [Google Scholar]
- AgriFutures Australia. AgriFutures Ginger Program RD&E Plan 2022–2027. Available online: https://www.agrifutures.com.au/rural-industries/ginger/ (accessed on 12 December 2022).
- Horticulture Innovation Australia Ltd. Australian Horticulture Statistics Handbook 2020/21. 2022. Available online: https://www.horticulture.com.au/growers/help-your-business-grow/research-reports-publications-fact-sheets-and-more/grower-resources/ha18002-assets/australian-horticulture-statistics-handbook/ (accessed on 12 December 2022).
- Australian Ginger Industry Association. Australian Ginger Industry Association. Available online: https://www.australianginger.org.au (accessed on 12 December 2022).
- Whiley, A.W. Effect of plant density on time to first harvest maturity, knob size and yield in two cultivars of ginger (Zingiber officinale Rosc.) grown in southeast Queensland. Aust. J. Exp. Agric. 1981, 58, 245–251. [Google Scholar]
- Keating, J. The Australian Ginger Industry. Available online: https://www.daf.qld.gov.au/data/assets/pdf_file/0006/74895/Australian-ginger-industry-report (accessed on 12 December 2022).
- Smith, M.K.; Hamill, S.D.; Gogel, B.J.; Severn-Ellis, A.A. Ginger (Zingiber officinale) autotetraploids with improved processing quality produced by an in vitro colchicine treatment. Aust. J. Exp. Agric. 2004, 44, 1065–1072. [Google Scholar] [CrossRef] [Green Version]
- Sanewski, G.M. Rhizome and Fibre Development in Early Harvest Ginger (Zingiber officinale Rosc). Ph.D. Thesis, The University of Queensland, Brisbane, Australia, 2002. Available online: https://espace.library.uq.edu.au/view/UQ:223854 (accessed on 12 December 2022).
- Wohlmuth, H.; Smith, M.K.; Brooks, L.O.; Myers, S.P.; Leach, D.N. Essential oil composition of diploid and tetraploid clones of ginger (Zingiber officinale Roscoe) grown in Australia. J. Agric. Food Chem. 2006, 54, 1414–1419. [Google Scholar] [CrossRef]
- Dohroo, N.P.; Kansal, S.; Ahluwalia, N. Studies on eco-farmer-friendly practices for management of soft rot of ginger (Zingiber officinale). Indian Phytopathol. 2015, 68, 93–96. [Google Scholar]
- Islam, M.M.; Khatun, F.; Faruk, M.I.; Rahman, M.M.; Hossain, M.A. Incidence of rhizome rot of ginger in some selected areas of Bangladesh and the causal pathogens associated with the disease. Bangladesh J. Agric. Res. 2019, 44, 569–576. [Google Scholar] [CrossRef]
- Chawla, S.; Rafie, R.; Likins, M.; Ren, S.; Ndegwa, E.; Mersha, Z. First report of Fusarium yellows and rhizome rot caused by Fusarium oxysporum f. sp. zingiberi on ginger in the continental United States. Plant Dis. 2021, 105, 3289. [Google Scholar] [CrossRef]
- Harsha, K.; Nair, A.R.; Tejaswini, G. Identification of Fusarium oxysporum as causative agent of rhizome rot disease in seed ginger rhizomes in Wayanad, Kerala. Indian Phytopathol. 2021, 74, 253–256. [Google Scholar] [CrossRef]
- Okwuowulu, P.A. Ginger in Africa and the Pacific Ocean islands. In Ginger: The Genus Zingiber; Ravindran, P.N., Babu, K.N., Eds.; CRC Press: Boca Raton, FL, USA, 2016; pp. 1–26. [Google Scholar]
- Pegg, K.G.; Moffett, M.L.; Colbran, R.C. Diseases of ginger in Queensland. Qld. Agric. J. 1974, 100, 611–618. [Google Scholar]
- Gordon, T.R. Fusarium oxysporum and the Fusarium wilt syndrome. Annu. Rev. Phytopathol. 2017, 55, 23–39. [Google Scholar] [CrossRef] [PubMed]
- Summerell, B.A.; Leslie, J.F. Fifty years of Fusarium: How could nine species have ever been enough? Fungal Divers. 2011, 50, 135–144. [Google Scholar] [CrossRef]
- Gupta, M.; Jarial, K.; Vikram, A. Morphological, cultural, pathological and molecular variability among Fusarium oxysporum f.sp. zingiberi isolates. Int. J. Bio-Resour. Stress Manag. 2014, 5, 375–380. [Google Scholar] [CrossRef]
- Leslie, J.F.; Summerell, B.A. Morphological Characters. In The Fusarium Laboratory Manual, 1st ed.; Blackwell Pub.: Ames, IA, USA, 2006. [Google Scholar] [CrossRef]
- Stirling, A.M. The causes of poor establishment of ginger (Zingiber officinale) in Queensland, Australia. Australas. Plant Pathol. 2004, 33, 203–210. [Google Scholar] [CrossRef]
- Grover, A.; Gowthaman, R. Strategies for development of fungus-resistant transgenic plants. Curr. Sci. 2003, 84, 330–340. [Google Scholar]
- Hartmann, A.; Berkowitz, O.; Whelan, J.; Narsai, R. Cross-species transcriptomic analyses reveals common and opposite responses in Arabidopsis, rice and barley following oxidative stress and hormone treatment. BMC Plant Biol. 2022, 22, 62. [Google Scholar] [CrossRef]
- Karthika, R.; Prasath, D.; Anandaraj, M. Comparative gene expression studies of candidate genes associated with defence response in ginger and mango ginger post inoculation with Ralstonia solanacearum. Physiol. Mol. Plant Pathol. 2018, 103, 1–7. [Google Scholar] [CrossRef]
- Belkhadir, Y.; Subramaniam, R.; Dangl, J.L. Plant disease resistance protein signaling: NBS–LRR proteins and their partners. Curr. Opin. Plant Biol. 2004, 7, 391–399. [Google Scholar] [CrossRef]
- Nishitani, K.; Vissenberg, K. Roles of the XTH protein family in the expanding cell. In The Expanding Cell; Verbelen, J.P., Vissenberg, K., Eds.; Plant Cell Monographs; Springer: Berlin/Heidelberg, Germany, 2006; Volume 6, pp. 89–116. [Google Scholar] [CrossRef]
- Gullner, G.; Komives, T.; Király, L.; Schröder, P. Glutathione S-transferase enzymes in plant-pathogen interactions. Front. Plant Sci. 2018, 9, 1836. [Google Scholar] [CrossRef] [Green Version]
- Chattopadhyay, A.; Purohit, J.; Tiwari, K.K.; Deshmukh, R. Targeting transcription factors for plant disease resistance: Shifting paradigm. Curr. Sci. 2019, 117, 1598–1607. [Google Scholar] [CrossRef]
- Ferreira, A.; Glas, V.B.; Louise, N. PCR from fungal spores after microwave treatment. Fungal Genet. Rep. 1996, 43, 25–26. [Google Scholar] [CrossRef] [Green Version]
- O’Donnell, K.; Kistler, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple evolutionary origins of the fungus causing Panama disease of banana: Concordant evidence from nuclear and mitochondrial gene genealogies. Proc. Natl. Acad. Sci. USA 1998, 95, 2044–2049. [Google Scholar] [CrossRef]
- Czislowski, E.; Fraser-Smith, S.; Zander, M.; O’Neill, W.T.; Meldrum, R.A.; Tran-Nguyen, L.T.T.; Batley, J.; Aitken, E.A.B. Investigation of the diversity of effector genes in the banana pathogen, Fusarium oxysporum f. sp. cubense, reveals evidence of horizontal gene transfer. Mol. Plant Pathol. 2018, 19, 1155–1171. [Google Scholar] [CrossRef] [Green Version]
- Czislowski, E.; Zeil-Rolfe, I.; Aitken, E.A.B. Effector profiles of endophytic Fusarium associated with asymptomatic banana (Musa sp.) hosts. Int. J. Mol. Sci. 2021, 22, 2508. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leslie, J.F.; Summerell, B.A. Techniques for recovering Fusarium. In The Fusarium Laboratory Manual; Blackwell Pub: Ames, IA, USA, 2006; pp. 15–20. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Staskawicz, B.J.; Ausubel, F.M.; Baker, B.J.; Ellis, J.G.; Jones, J.D. Molecular genetics of plant disease resistance. Science 1995, 268, 661–667. [Google Scholar] [CrossRef] [Green Version]
- Schwessinger, B.; Ronald, P.C. Plant innate immunity: Perception of conserved microbial signatures. Annu. Rev. Plant Biol. 2012, 63, 451–482. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Wang, W.; Zhang, T.; Gong, Z.; Zhao, H.; Han, G.-Z. Out of Water: The Origin and early diversification of plant R-genes. Plant Physiol. 2018, 177, 82–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.X.; Yano, M.; Yamanouchi, U.; Iwamoto, M.; Monna, L.; Hayasaka, H.; Katayose, Y.; Sasaki, T. The Pib gene for rice blast resistance belongs to the nucleotide binding and leucine-rich repeat class of plant disease resistance genes. Plant J. 1999, 19, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Radwan, O.; Mouzeyar, S.; Nicolas, P.; Bouzidi, M.F. Induction of a sunflower CC-NBS-LRR resistance gene analogue during incompatible interaction with Plasmopara halstedii. J. Exp. Bot. 2005, 56, 567–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, H.; Zhao, Z.; Malik, A.A.; Qian, C.; Chen, J. Identification and characterization of potential NBS-encoding resistance genes and induction kinetics of a putative candidate gene associated with downy mildew resistance in Cucumis. BMC Plant Biol. 2010, 10, 186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, S.; Ellwood, S.; Calis, O.; Patrick, E.; Li, T.; Coleman, M.; Turner, J.G. Broad-spectrum mildew resistance in Arabidopsis thaliana mediated by RPW8. Science 2001, 291, 118–120. [Google Scholar] [CrossRef] [PubMed]
- Karthika, R.; Prasath, D.; Anandaraj, M. Transcriptome-wide identification and characterization of resistant gene analogs (RGAs) of ginger (Zingiber officinale Rosc.) and mango ginger (Curcuma amada Roxb.) under stress induced by pathogen. Sci. Hortic. 2019, 248, 81–88. [Google Scholar] [CrossRef]
- Lorang, J.M.; Sweat, T.A.; Wolpert, T.J. Plant disease susceptibility conferred by a “resistance” gene. Proc. Natl. Acad. Sci. USA 2007, 104, 14861–14866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutterson, N.; Reuber, T.L. Regulation of disease resistance pathways by AP2/ERF transcription factors. Curr. Opin. Plant Biol. 2004, 7, 465–471. [Google Scholar] [CrossRef] [PubMed]
- Hawku, M.D.; Goher, F.; Islam, M.A.; Guo, J.; He, F.; Bai, X.; Yuan, P.; Kang, Z.; Guo, J. TaAP2-15, An AP2/ERF transcription factor, is positively involved in wheat resistance to Puccinia striiformis f. sp. tritici. Int. J. Mol. Sci. 2021, 22, 2080. [Google Scholar] [CrossRef]
- Gu, Y.Q.; Wildermuth, M.C.; Chakravarthy, S.; Loh, Y.T.; Yang, C.; He, X.; Han, Y.; Martin, G.B. Tomato transcription factors pti4, pti5, and pti6 activate defense responses when expressed in Arabidopsis. Plant Cell. 2002, 14, 817–831. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Chai, C.; Ai, G.; Jia, Y.; Liu, W.; Zhang, X.; Bai, T.; Dou, D. A Nicotiana benthamiana AP2/ERF transcription factor confers resistance to Phytophthora parasitica. Phytopathol. Res. 2020, 2, 4. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Zhang, D.; Chen, J.; Yang, Y.; Huang, Z.; Huang, D.; Wang, X.-C.; Huang, R. Tomato stress-responsive factor TSRF1 interacts with ethylene responsive element GCC box and regulates pathogen resistance to Ralstonia solanacearum. Plant Mol. Biol. 2004, 55, 825–834. [Google Scholar] [CrossRef]
- Zhou, J.; Tang, X.; Martin, G.B. The Pto kinase conferring resistance to tomato bacterial speck disease interacts with proteins that bind a cis-element of pathogenesis-related genes. Embo J. 1997, 16, 3207–3218. [Google Scholar] [CrossRef] [Green Version]
- Eulgem, T.; Somssich, I.E. Networks of WRKY transcription factors in defense signaling. Curr. Opin. Plant Biol. 2007, 10, 366–371. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Deyholos, M.K. Functional characterization of Arabidopsis NaCl-inducible WRKY25 and WRKY33 transcription factors in abiotic stresses. Plant Mol. Biol. 2009, 69, 91–105. [Google Scholar] [CrossRef]
- Pandey, S.P.; Somssich, I.E. The role of WRKY transcription factors in plant immunity. Plant Physiol. 2009, 150, 1648–1655. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.F.; Liu, J.K.; Yang, F.M.; Zhang, G.Y.; Wang, D.; Zhang, L.; Ou, Y.B.; Yao, Y.A. The WRKY transcription factor WRKY8 promotes resistance to pathogen infection and mediates drought and salt stress tolerance in Solanum lycopersicum. Physiol. Plant 2020, 168, 98–117. [Google Scholar] [CrossRef]
- Fu, Z.Q.; Dong, X. Systemic acquired resistance: Turning local infection into global defense. Annu. Rev. Plant Biol. 2013, 64, 839–863. [Google Scholar] [CrossRef] [Green Version]
- Pieterse, C.M.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Van Wees, S.C.; Bakker, P.A. Induced systemic resistance by beneficial microbes. Annu. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef] [Green Version]
- Ahuja, I.; Kissen, R.; Bones, A.M. Phytoalexins in defense against pathogens. Trends Plant Sci. 2012, 17, 73–90. [Google Scholar] [CrossRef]
- Li, R.; Tee, C.S.; Jiang, Y.L.; Jiang, X.Y.; Venkatesh, P.N.; Sarojam, R.; Ye, J. A terpenoid phytoalexin plays a role in basal defense of Nicotiana benthamiana against Potato virus X. Sci. Rep. 2015, 5, 9682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ninkuu, V.; Zhang, L.; Yan, J.; Fu, Z.; Yang, T.; Zeng, H. Biochemistry of terpenes and recent advances in plant protection. Int. J. Mol. Sci. 2021, 22, 5710. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Vallet, A.; Ramos, B.; Bednarek, P.; López, G.; Piślewska-Bednarek, M.; Schulze-Lefert, P.; Molina, A. Tryptophan-derived secondary metabolites in Arabidopsis thaliana confer non-host resistance to necrotrophic Plectosphaerella cucumerina fungi. Plant J. 2010, 63, 115–127. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Concepción, M. Early steps in isoprenoid biosynthesis: Multilevel regulation of the supply of common precursors in plant cells. Phytochem. Rev. 2006, 5, 1–15. [Google Scholar] [CrossRef]
- Nagegowda, D.A.; Bach, T.J.; Chye, M.L. Brassica juncea 3-hydroxy-3-methylglutaryl (HMG)-CoA synthase 1: Expression and characterization of recombinant wild-type and mutant enzymes. Biochem. J. 2004, 383, 517–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, E.J.; Schenk, P.M.; Kazan, K.; Penninckx, I.A.; Anderson, J.P.; Maclean, D.J.; Cammue, B.P.; Ebert, P.R.; Manners, J.M. Pathogen-responsive expression of a putative ATP-binding cassette transporter gene conferring resistance to the diterpenoid sclareol is regulated by multiple defense signaling pathways in Arabidopsis. Plant Physiol. 2003, 133, 1272–1284. [Google Scholar] [CrossRef] [Green Version]
- Pierman, B.; Toussaint, F.; Bertin, A.; Lévy, D.; Smargiasso, N.; De Pauw, E.; Boutry, M. Activity of the purified plant ABC transporter NtPDR1 is stimulated by diterpenes and sesquiterpenes involved in constitutive and induced defenses. J. Biol. Chem. 2017, 292, 19491–19502. [Google Scholar] [CrossRef] [Green Version]
- Underwood, W. The plant cell wall: A dynamic barrier against pathogen invasion. Front. Plant Sci. 2012, 3, 85. [Google Scholar] [CrossRef] [Green Version]
- Bani, M.; Cimmino, A.; Evidente, A.; Rubiales, D.; Rispail, N. Pisatin involvement in the variation of inhibition of Fusarium oxysporum f. sp. pisi spore germination by root exudates of Pisum spp. germplasm. Plant Pathol. 2018, 67, 1046–1054. [Google Scholar] [CrossRef]
- Jacobs, A.K.; Lipka, V.; Burton, R.A.; Panstruga, R.; Strizhov, N.; Schulze-Lefert, P.; Fincher, G.B. An Arabidopsis callose synthase, GSL5, is required for wound and papillary callose formation. Plant Cell. 2003, 15, 2503–2513. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, M.T.; Stein, M.; Hou, B.H.; Vogel, J.P.; Edwards, H.; Som erville, S.C. Loss of a callose synthase results in salicylic acid-dependent disease resistance. Science 2003, 301, 969–972. [Google Scholar] [CrossRef]
- Vierling, E. The roles of heat shock proteins in plants. Annu. Rev. Plant Physiol Plant Mol. Biol. 1991, 42, 579–620. [Google Scholar] [CrossRef]
- Lee, J.H.; Yun, H.S.; Kwon, C. Molecular communications between plant heat shock responses and disease resistance. Mol. Cells 2012, 34, 109–116. [Google Scholar] [CrossRef] [Green Version]
- Acevedo-Garcia, J.; Gruner, K.; Reinstädler, A.; Kemen, A.; Kemen, E.; Cao, L.; Takken, F.L.W.; Reitz, M.U.; Schäfer, P.; O’Connell, R.J.; et al. The powdery mildew-resistant Arabidopsis mlo2 mlo6 mlo12 triple mutant displays altered infection phenotypes with diverse types of phytopathogens. Sci. Rep. 2017, 7, 9319. [Google Scholar] [CrossRef] [Green Version]
- Birch, P.R.J.; Cooke, D.E.L. The early days of late blight. eLife 2013, 2, e00954. [Google Scholar] [CrossRef]
- Fry, W. Phytophthora infestans: The plant (and R gene) destroyer. Mol. Plant Pathol. 2008, 9, 385–402. [Google Scholar] [CrossRef]
- Mokyr, J.; GrÁDa, C.Ó. What do people die of during famines: The great Irish famine in comparative perspective. Eur. Rev. Econ. Hist. 2002, 6, 339–363. [Google Scholar] [CrossRef] [Green Version]
- Ploetz, R.C. Management of Fusarium wilt of banana: A review with special reference to tropical race 4. Crop. Prot. 2015, 73, 7–15. [Google Scholar] [CrossRef]
- Parker, E.D. Ecological implications of clonal diversity in parthenogenetic morphospecies. Am. Zool 1979, 19, 753–762. [Google Scholar] [CrossRef]
- Hamill, S.D.; Moisander, J.A.; Smith, M.K. Micropropagation of vegetatively propagated crops: Accelerating release of new cultivars and providing an important source of clean planting material. Acta Hortic. 2009, 829, 213–218. [Google Scholar] [CrossRef]
- Dake, G.N. Diseases of ginger (Zingiber officinale Rosc.) and their management. J. Spices Aromat. Crops 1995, 4, 40–48. [Google Scholar]
- O’Neill, W.T.; Pattison, A.B.; Daniells, J.W.; Hermanto, C.; Molina, A.B. Vegetative compatibility group analysis of Indonesian Fusarium oxysporum f. sp. cubense isolates. Acta Hortic. 2011, 897, 345–351. [Google Scholar] [CrossRef]
- Puhalla, J.E. Classification of strains of Fusarium oxysporum on the basis of vegetative compatibility. Can. J. Bot. 1985, 63, 179–183. [Google Scholar] [CrossRef]
- Pappalardo, L.; Smith, M.K.; Hamill, S.D.; Stirling, A.M.; McKay, D. DNA amplification fingerprinting analysis of genetic variation within Fusarium oxysporum f.sp. zingiberi. Australas. Plant Pathol 2009, 38, 51–54. [Google Scholar] [CrossRef]
- Teakle, D. Fusarium rhizome rot of ginger in Queensland. Queensl. J. Agric. Anim. Sci. 1965, 22, 265–272. [Google Scholar]
- Warman, N.M.; Aitken, E.A. The movement of Fusarium oxysporum f. sp. cubense (sub-tropical race 4) in susceptible cultivars of banana. Front. Plant Sci. 2018, 9, 1748. [Google Scholar]
- Baayen, R.; De Maat, A. Passive transport of microconidia of Fusarium oxysporum f. sp. dianthi in carnation after root inoculation. Neth. J. Plant Pathol. 1987, 93, 3–13. [Google Scholar] [CrossRef]
- Cobon, J.A.; Pattison, A.B.; Penrose, L.D.J.; Chandra, K.A.; O’Neill, W.T.; Smith, M.K. Comparison of the reproduction and pathogenicity of isolates of Radopholus similis (burrowing nematode) from Australia and Fiji on ginger (Zingiber officinale) and banana (Musa spp.). Australas. Pant Pathol. 2019, 48, 529–539. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5′ to 3′) |
|---|---|
| ZoNBS1-F | TTGGATTCCGGAGGACAAAC |
| ZoNBS1-R | CATCCTAAGAGTGGCCAAGAAG |
| ZoNBS2-F | ACGTGTGGAGCGATGATAAG |
| ZoNBS2-R | ATGTCCCGAGTTGTAACAATGA |
| ZoNBS3-F | GGAAGATGGGCTGGCTTAAT |
| ZoNBS3-R | GCTGGCTTCTCTTTCCTCTAAT |
| ZoWRKY8-F | TGTTTCCATCTCCTACGTCTG |
| ZoWRKY8-R | GTTGGACTTGACCTCATCCT |
| ZoAP2-F | ATGTCAAGGAGCGGCAATAC |
| ZoAP2-R | CATATCCTCGACCGCTTGTTC |
| ZoERF2-F | TCTGAATCCTCCTCGATCAT |
| ZoERF2-R | AGGAGGTTAAGGTCGAAATTG |
| ZoABC-F | TCTGGGACACCTACTGATATTG |
| ZoABC-R | GAATTGCTGATGAAGGGACAG |
| ZoHMGR-F | TGGTCTGTGAAGCAATTATCA |
| ZoHMGR-R | ACCCAGCAAGGTTCTTAATC |
| ZoHMGS-F | TAGATACGGAGCCAAGGATT |
| ZoHMGS-R | GCATAATGTCGACGGTACAT |
| ZoMLO12-F | GTGTATTGCCGCCTTATCTT |
| ZoMLO12-R | CTTTGTAGTTCGATCCCATCTG |
| ZoCalS1-F | GTCCTGAAACCTCTTTCTAGTG |
| ZoCalS1-R | GCAAGGAAGAGAGGCAATAC |
| ZoXTG-F | CAGCAGTTCTACCTCTGGTTC |
| ZoXTG-R | CGTCCCGTCCACATAAATCA |
| ZoGST-F | TGCCGTCTTTGGAGCATAAC |
| ZoGST-R | CGGGATCATCAGGCAGTAATTT |
| ZoBAC-F | GCATCACTGACAGAGAGAAGA |
| ZoBAC-R | GAGCACAAACTGAGGGAGTA |
| ZoHSP-F | AAAGAGGACCAGCTGGAATAC |
| ZoHSP-R | GGTGGTCTTCTCTGTCCATAAA |
| ZoActin-F | TAGGTGCCCAGAGGTTCTATT |
| ZoActin-R | ACCGCTAAGCACCACATTAC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prasath, D.; Matthews, A.; O’Neill, W.T.; Aitken, E.A.B.; Chen, A. Fusarium Yellows of Ginger (Zingiber officinale Roscoe) Caused by Fusarium oxysporum f. sp. zingiberi Is Associated with Cultivar-Specific Expression of Defense-Responsive Genes. Pathogens 2023, 12, 141. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens12010141
Prasath D, Matthews A, O’Neill WT, Aitken EAB, Chen A. Fusarium Yellows of Ginger (Zingiber officinale Roscoe) Caused by Fusarium oxysporum f. sp. zingiberi Is Associated with Cultivar-Specific Expression of Defense-Responsive Genes. Pathogens. 2023; 12(1):141. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens12010141
Chicago/Turabian StylePrasath, Duraisamy, Andrea Matthews, Wayne T. O’Neill, Elizabeth A. B. Aitken, and Andrew Chen. 2023. "Fusarium Yellows of Ginger (Zingiber officinale Roscoe) Caused by Fusarium oxysporum f. sp. zingiberi Is Associated with Cultivar-Specific Expression of Defense-Responsive Genes" Pathogens 12, no. 1: 141. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens12010141