
Antimicrobial and Biofilm-Preventing Activity of l-Borneol Possessing 2(5H)-Furanone Derivative F131 against S. aureus—C. albicans Mixed Cultures
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
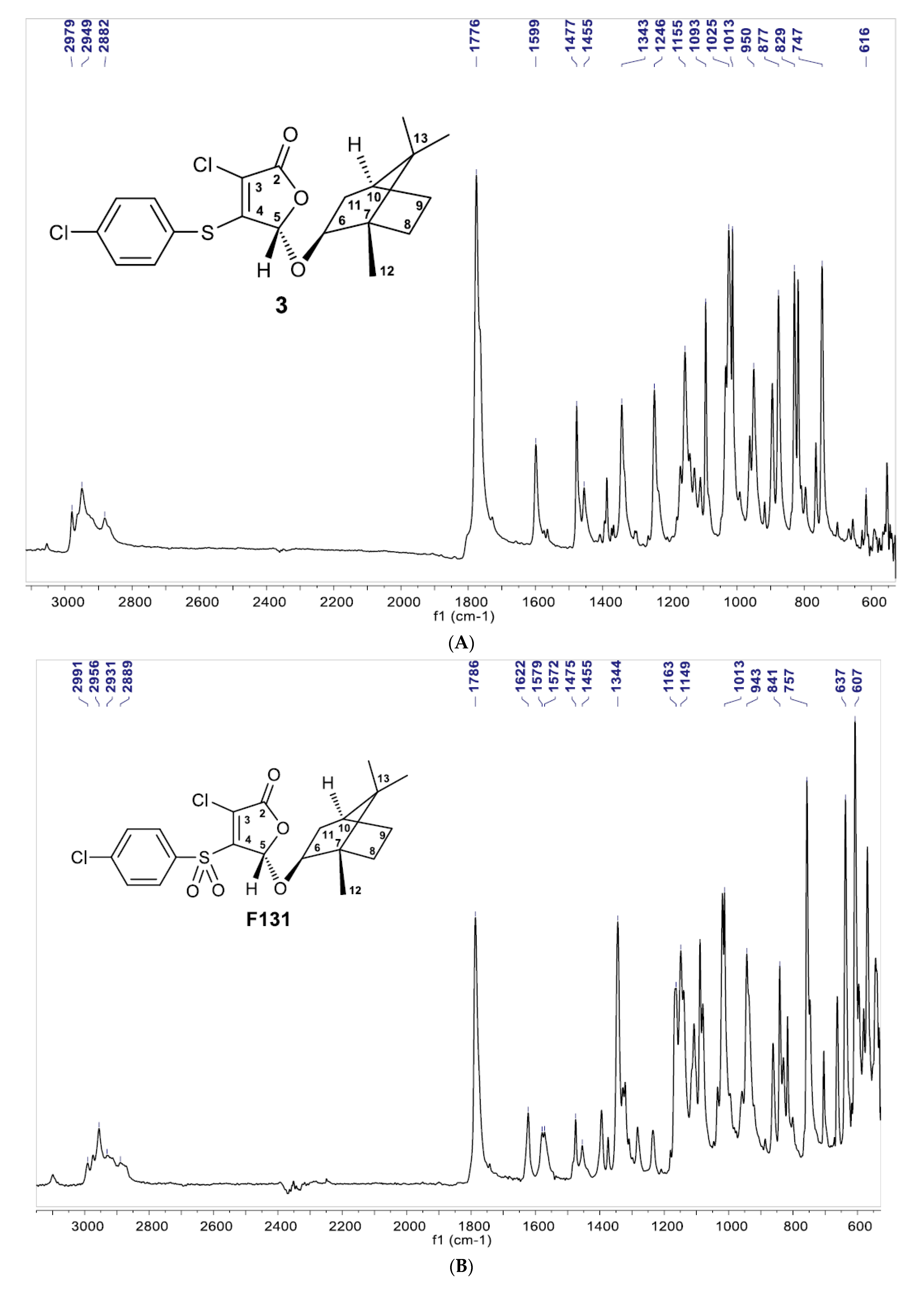
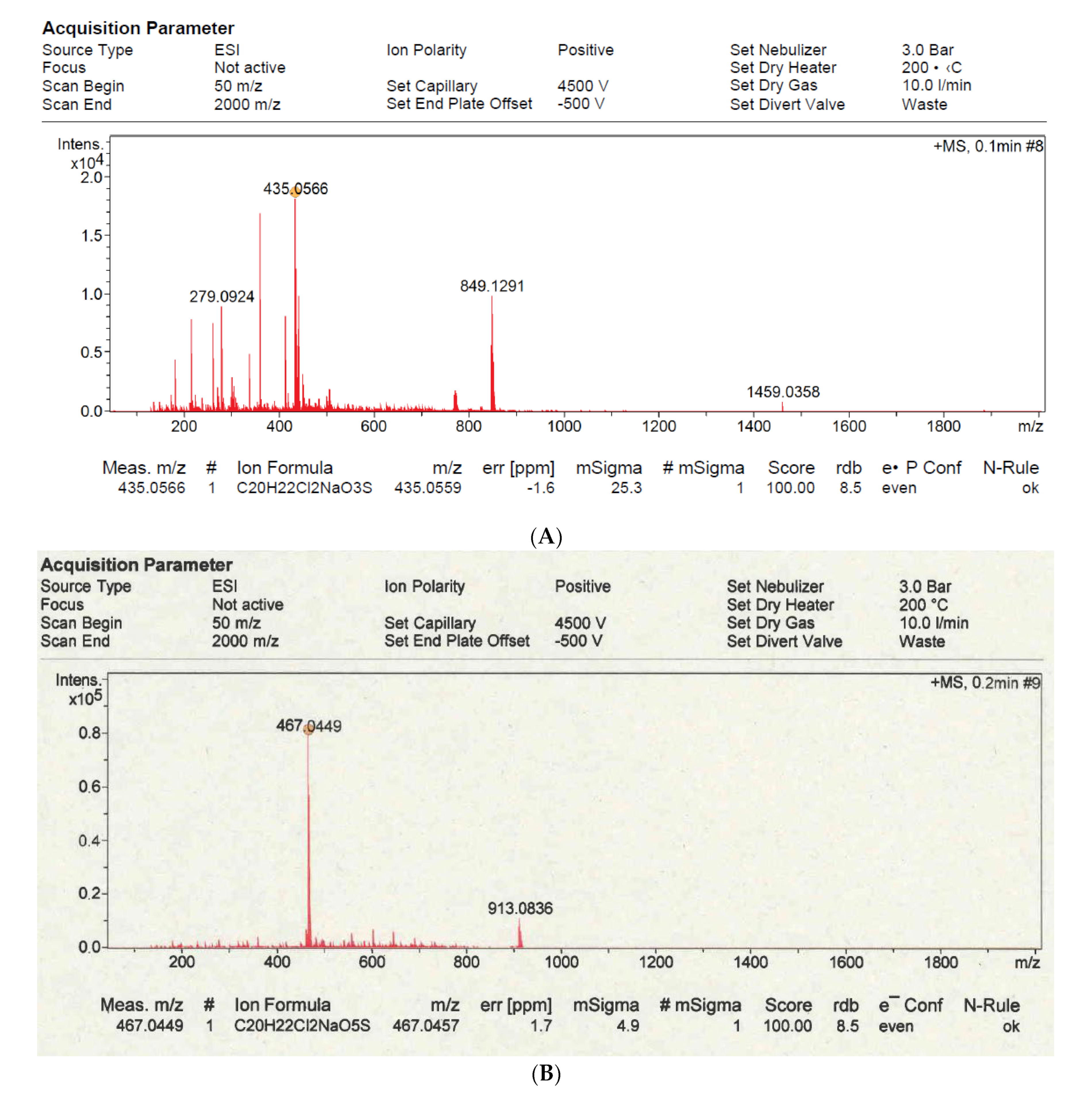
2.1. Chemistry (General Information)
2.2. Synthesis
2.3. Antimicrobials
2.4. Strains and Growth Conditions
2.5. Determination of the Minimal Inhibitory (MIC) and the Minimal Bactericidal/Fungicidal Concentrations (MBC/MFC) of F131 and Reference Antimicrobials
2.6. Analysis of the Biofilm–Prevention Concentration (BPC)
2.7. Assessment of Synergy between F131 and Conventional Antimicrobials
2.8. Quantification of Viable Cells
2.9. Confocal laser Scanning Microscopy (CLSM)
2.10. RNA Isolation and Real-Time One-Step qRT-PCR
2.11. Data Analysis
3. Results
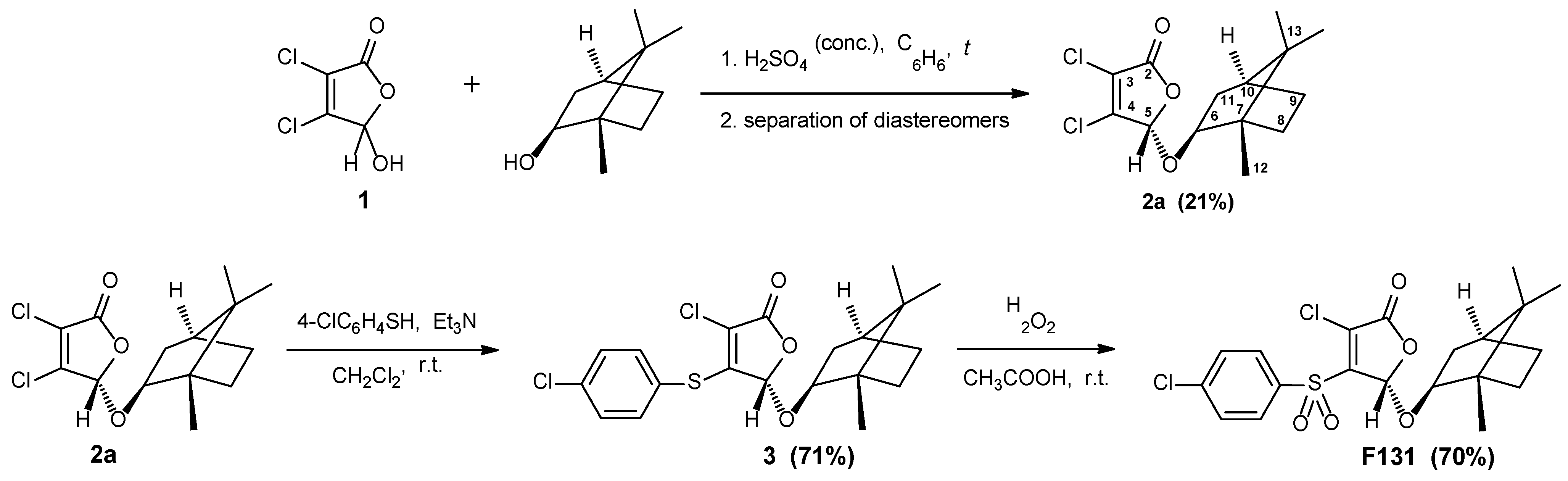
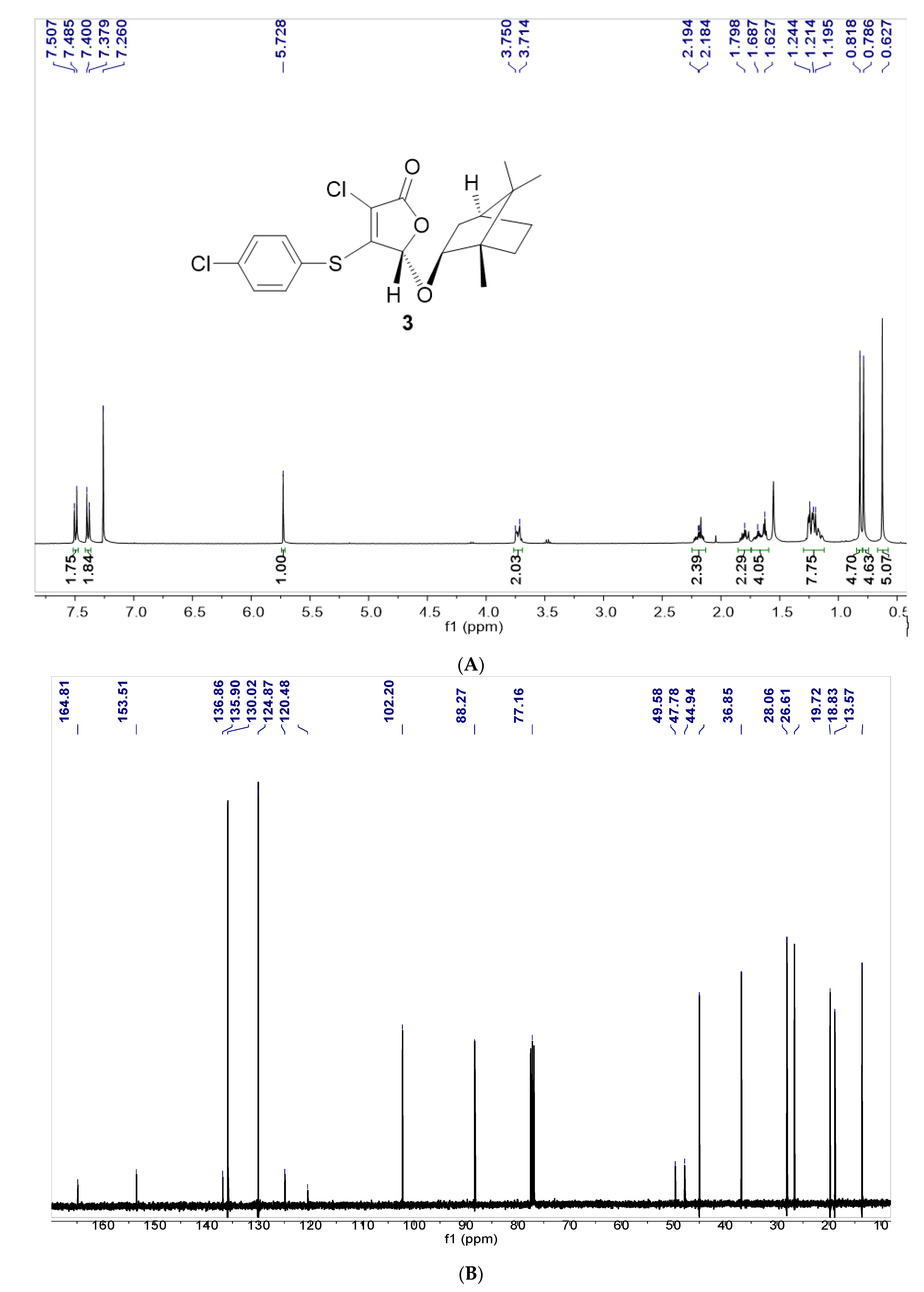
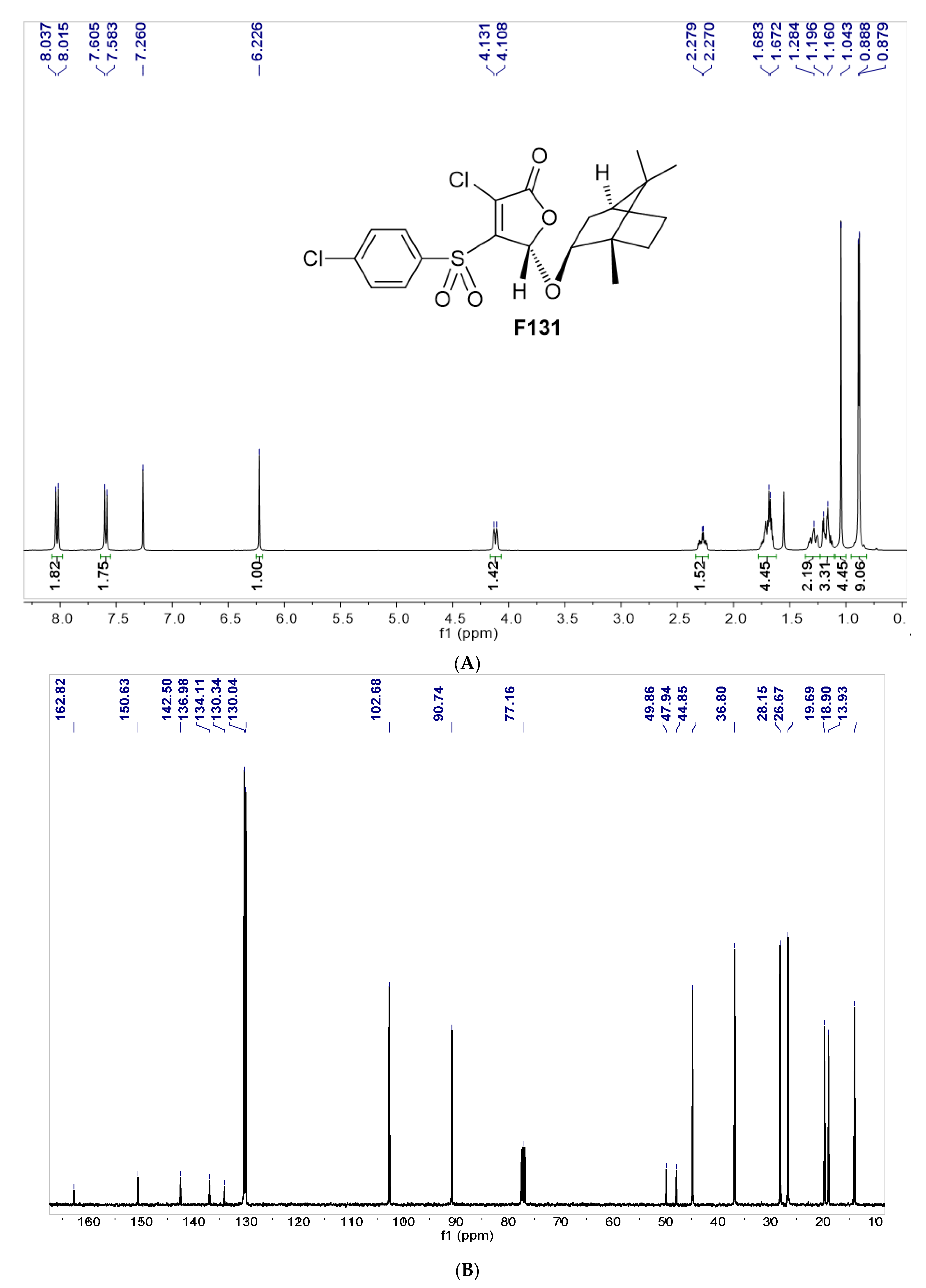
3.1. Synthesis of Furanone F131
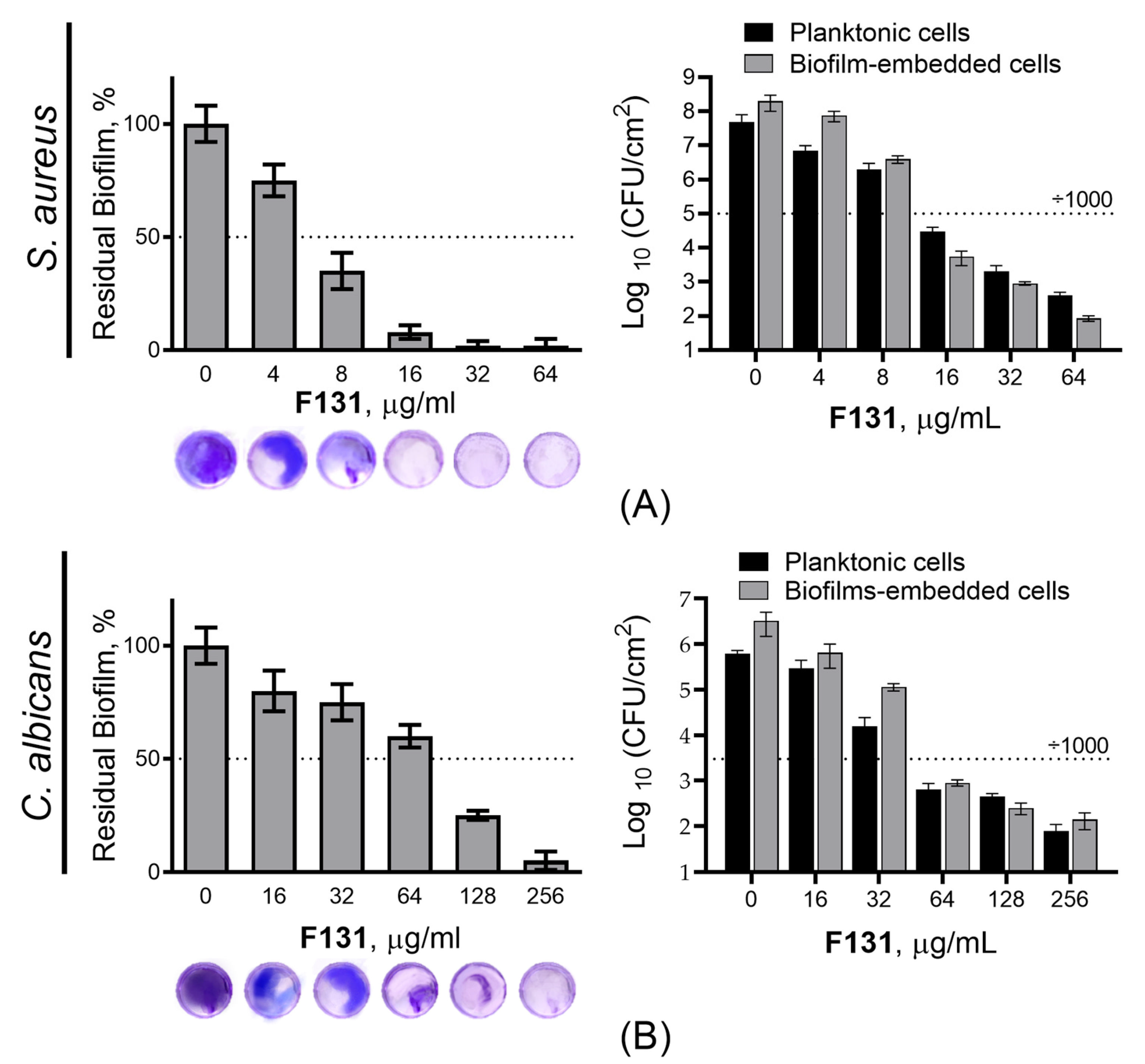
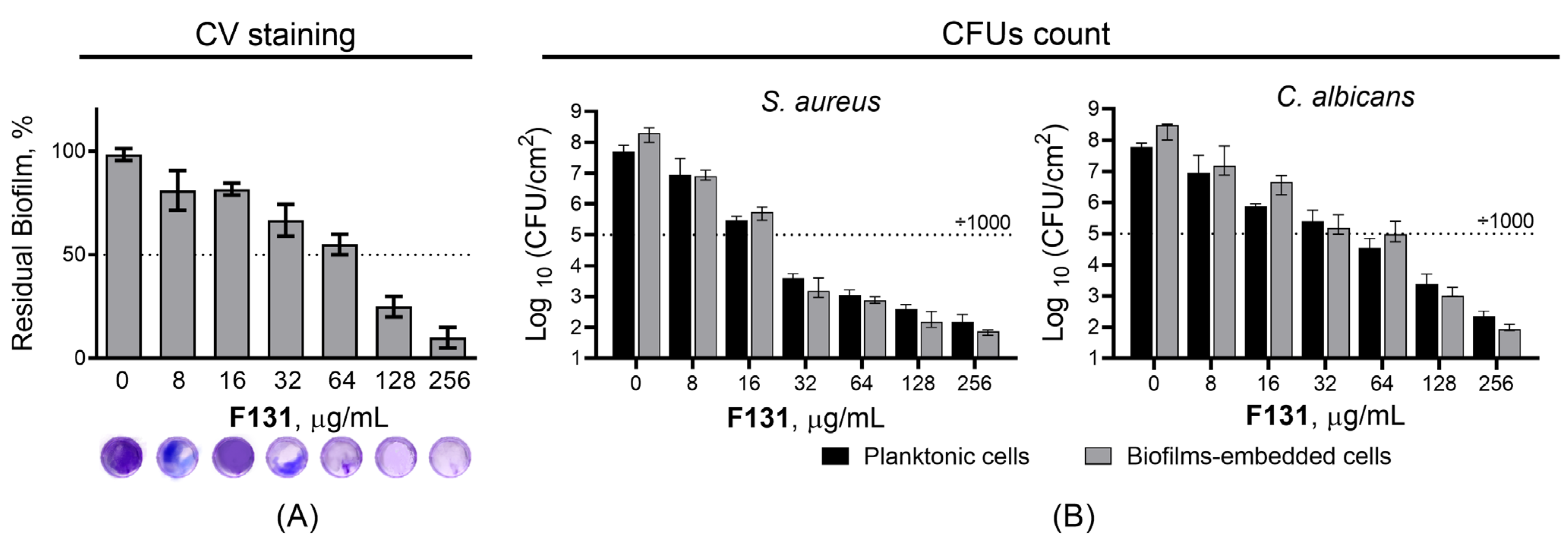
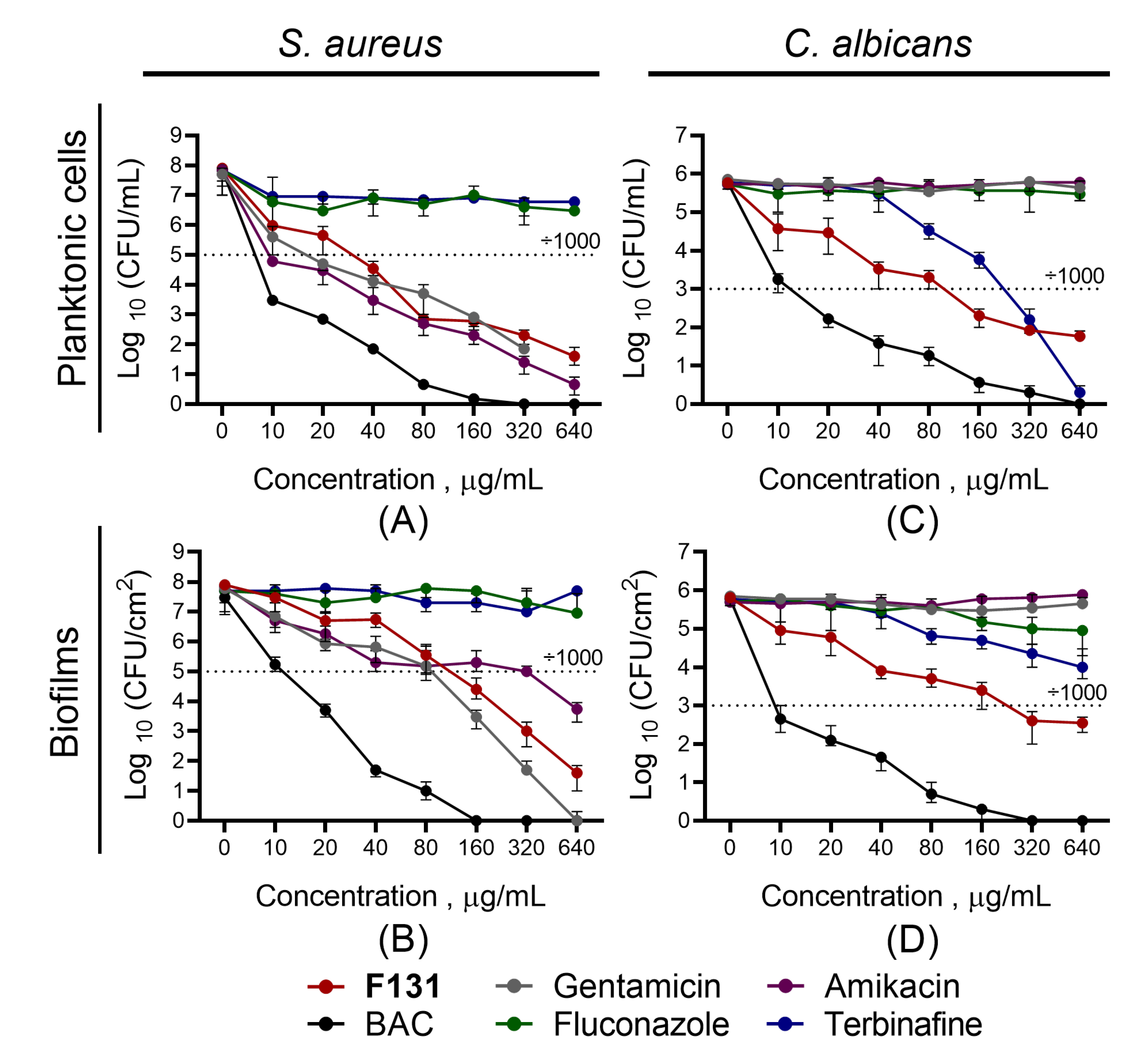
3.2. Antimicrobial and Biofilm Preventing Activity of F131
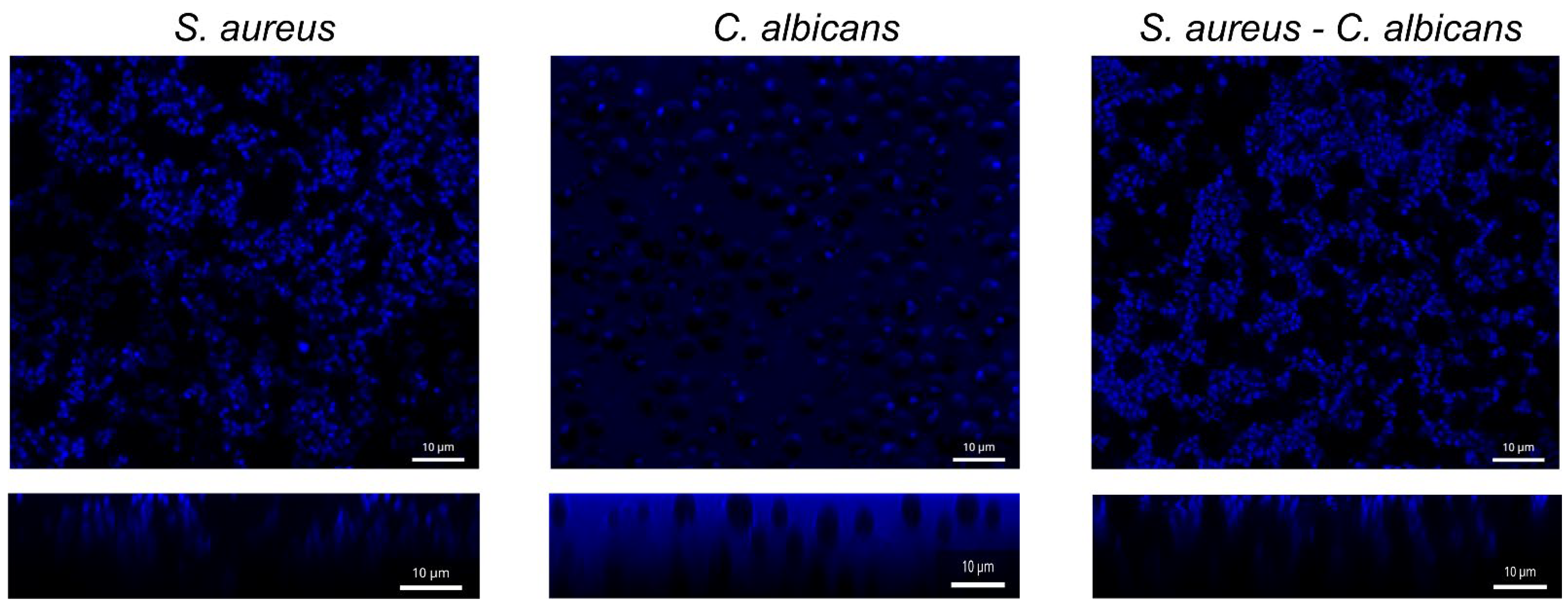
3.3. Antibiofilm Activity of F131 on S. aureus and C. albicans Mixed Biofilms
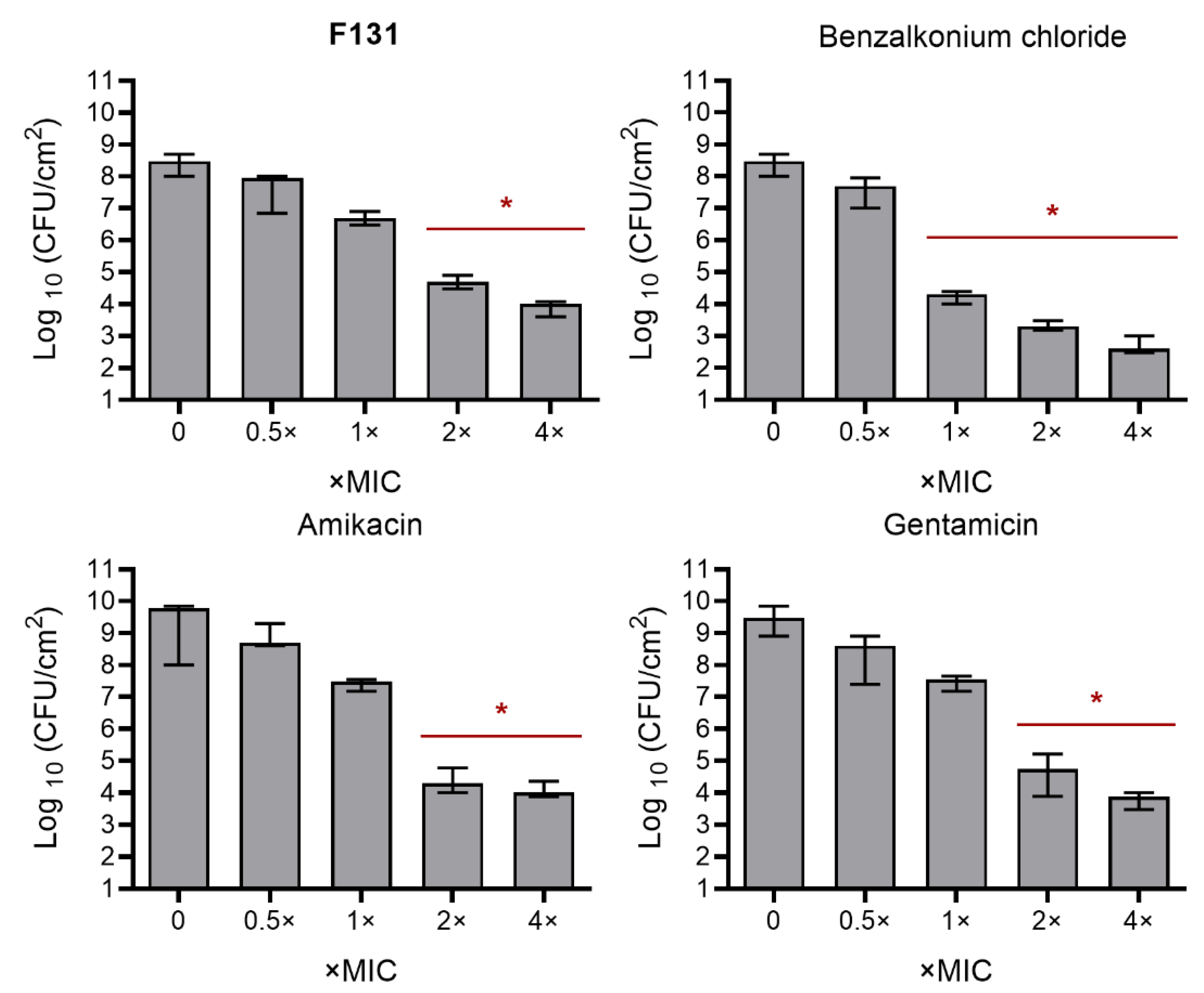
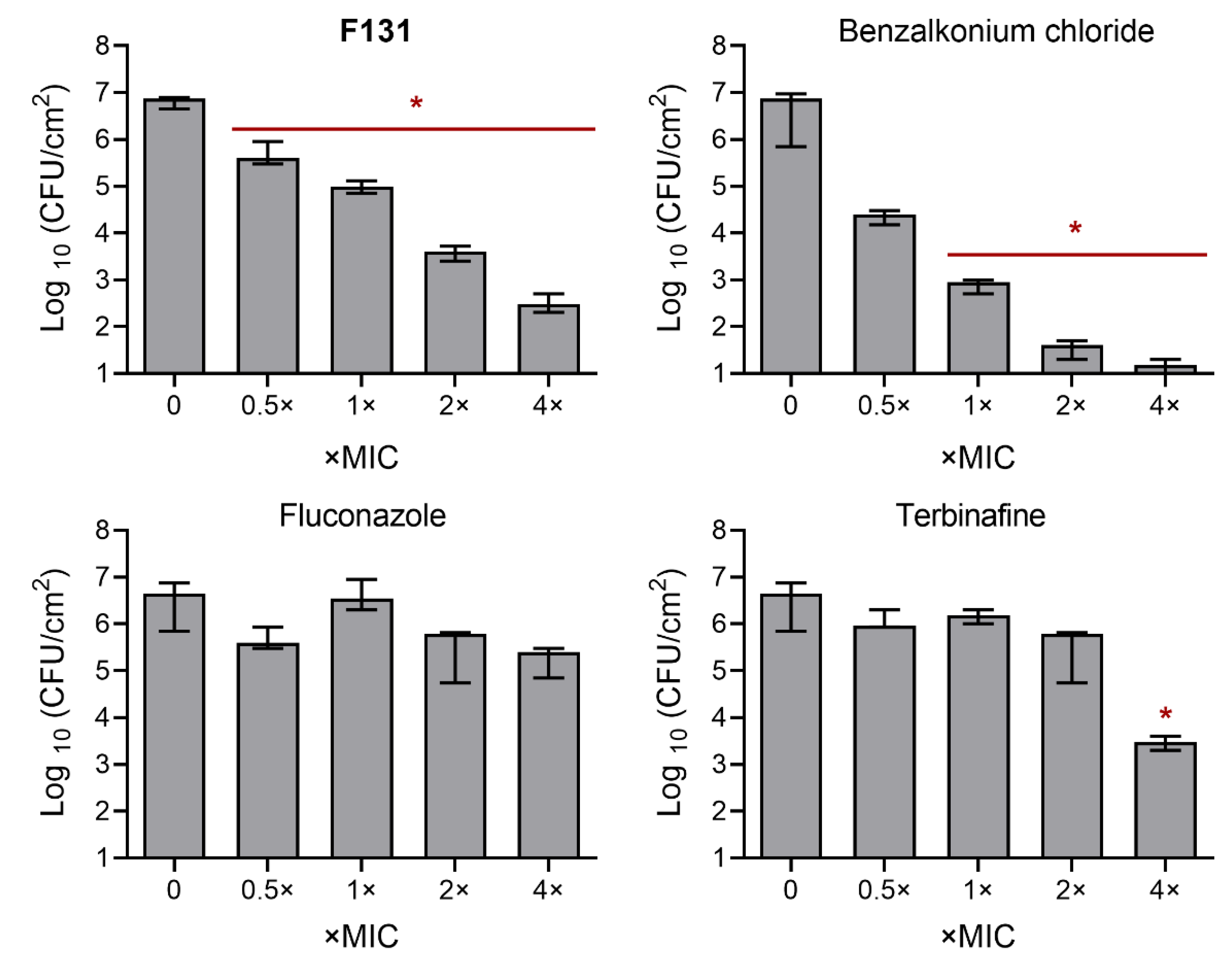
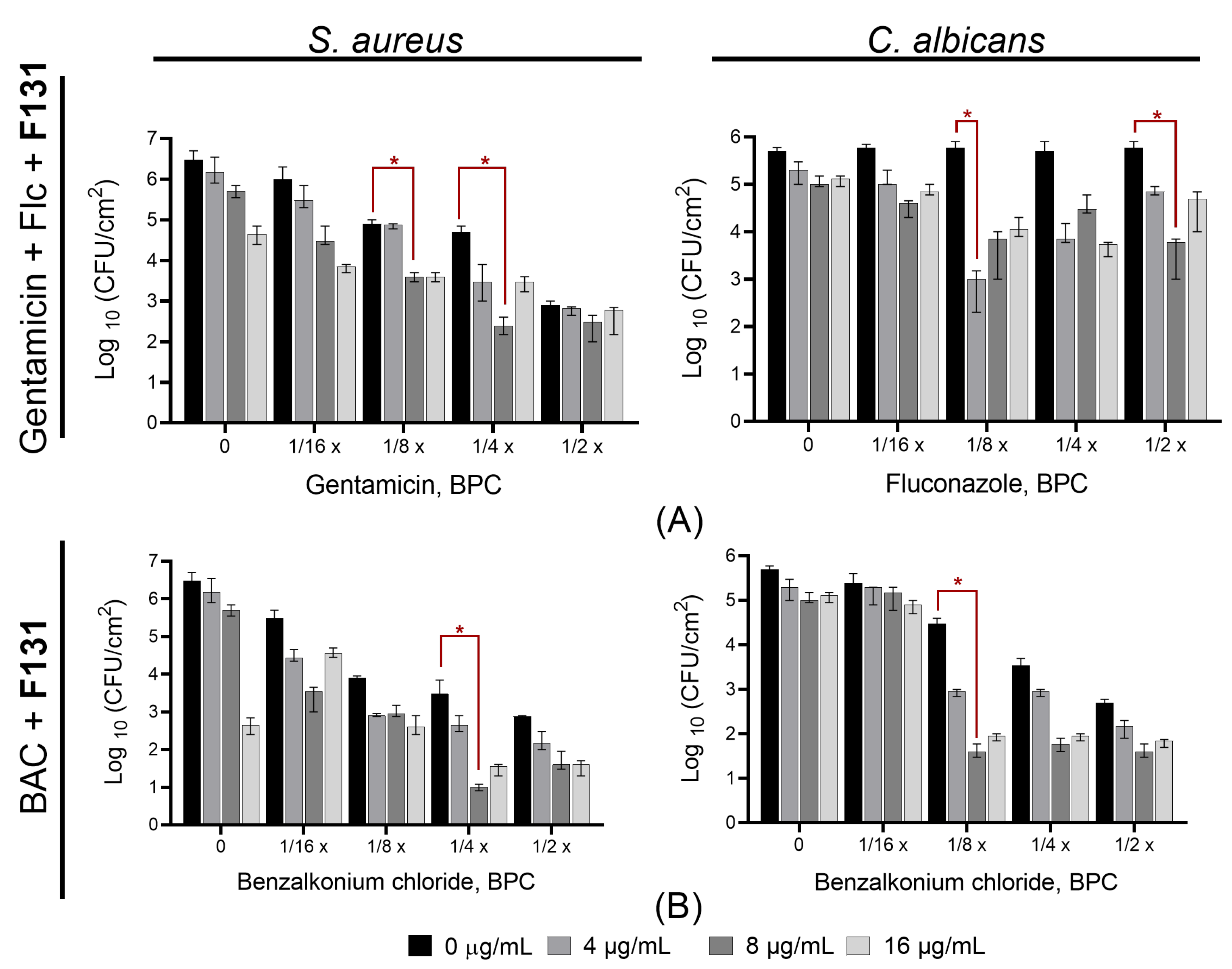
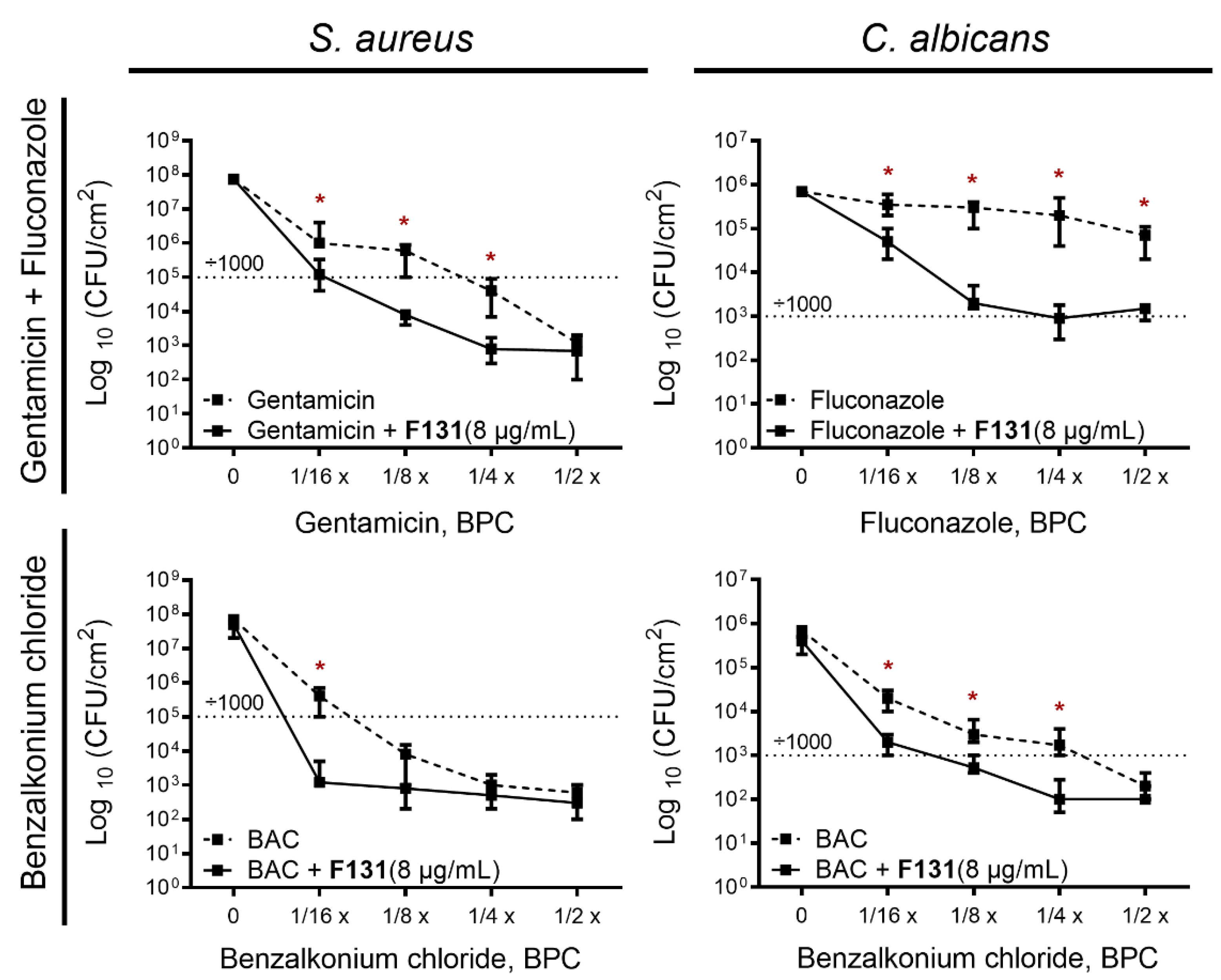
3.4. Synergistic Effects of F131 with Other Antimicrobials on S. aureus–C. albicans Mixed Biofilms
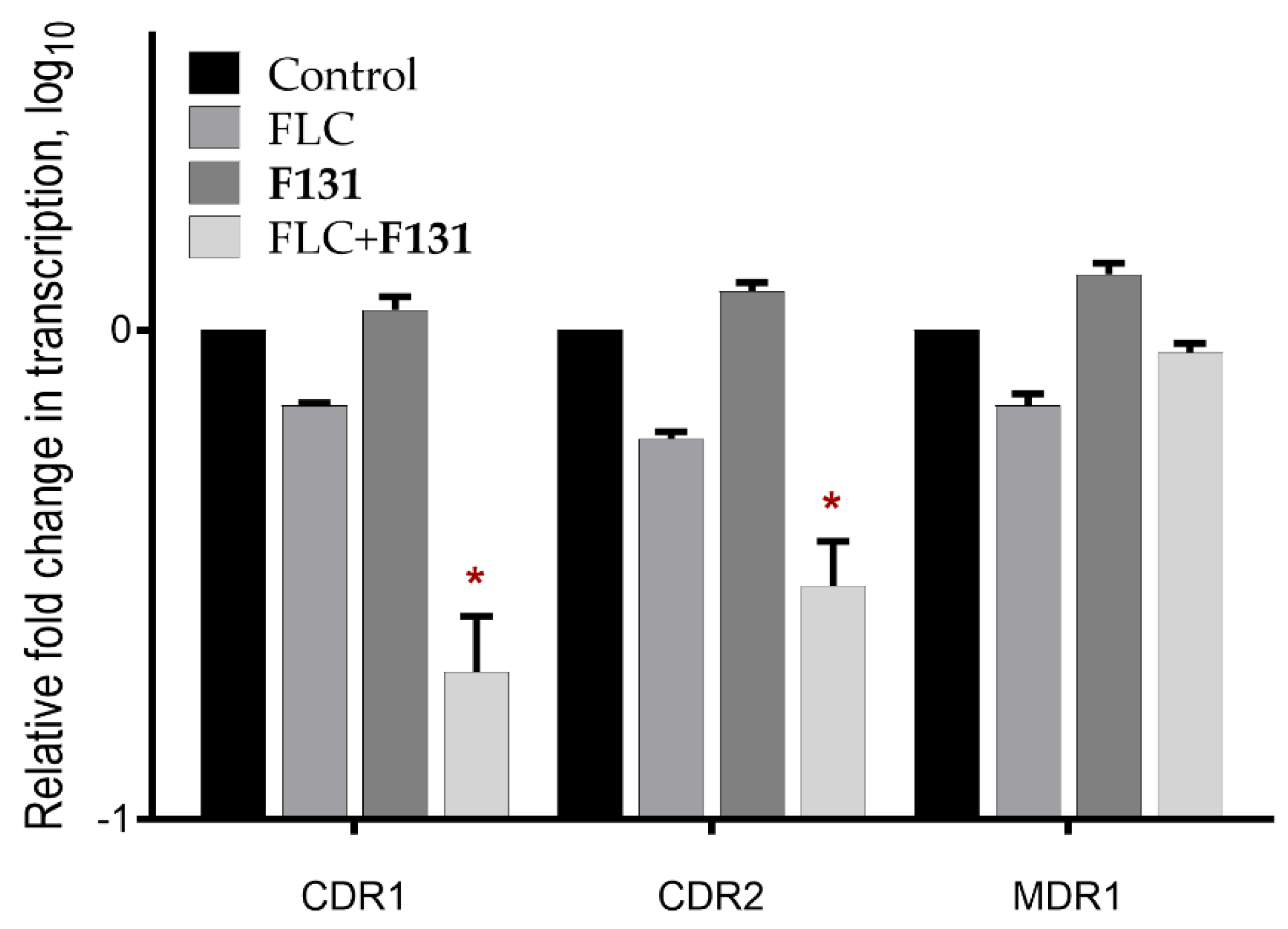
3.5. The Combination of F131 with Fluconazole Significantly Decreased the Expression Level of CDR1 and CDR2 Genes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A




References
- Muthukrishnan, G.; Masters, E.A.; Daiss, J.L.; Schwarz, E.M. Mechanisms of Immune Evasion and Bone Tissue Colonization That Make Staphylococcus aureus the Primary Pathogen in Osteomyelitis. Curr. Osteoporos. Rep. 2019, 17, 395–404. [Google Scholar] [CrossRef] [PubMed]
- Budzyńska, A.; Skowron, K.; Kaczmarek, A.; Wietlicka-Piszcz, M.; Gospodarek-Komkowska, E. Virulence Factor Genes and Antimicrobial Susceptibility of Staphylococcus aureus Strains Isolated from Blood and Chronic Wounds. Toxins 2021, 13, 491. [Google Scholar] [CrossRef] [PubMed]
- Pichereau, S.; Rose, W.E. Invasive community-associated MRSA infections: Epidemiology and antimicrobial management. Expert Opin. Pharmacother. 2010, 11, 3009–3025. [Google Scholar] [CrossRef] [PubMed]
- Sharma, N.K.; Garg, R.; Baliga, S.; Bhat, K.G. Nosocomial Infections and Drug Susceptibility Patterns in Methicillin Sensitive and Methicillin Resistant Staphylococcus aureus. J. Clin. Diagn. Res. 2013, 7, 2178–2180. [Google Scholar] [CrossRef]
- Turner, N.A.; Sharma-Kuinkel, B.K.; Maskarinec, S.A.; Eichenberger, E.M.; Shah, P.P.; Carugati, M.; Holland, T.L.; Fowler, V.G., Jr. Methicillin-resistant Staphylococcus aureus: An overview of basic and clinical research. Nat. Rev. Genet. 2019, 17, 203–218. [Google Scholar] [CrossRef]
- Cheung, G.Y.C.; Bae, J.S.; Otto, M. Pathogenicity and virulence of Staphylococcus aureus. Virulence 2021, 12, 547–569. [Google Scholar] [CrossRef]
- Miao, J.; Chen, L.; Wang, J.; Wang, W.; Chen, D.; Li, L.; Li, B.; Deng, Y.; Xu, Z. Current methodologies on genotyping for nosocomial pathogen methicillin-resistant Staphylococcus aureus (MRSA). Microb. Pathog. 2017, 107, 17–28. [Google Scholar] [CrossRef]
- Khan, A.; Wilson, B.; Gould, I.M. Current and future treatment options for community-associated MRSA infection. Expert Opin. Pharmacother. 2018, 19, 457–470. [Google Scholar] [CrossRef]
- Nataraj, B.H.; Mallappa, R.H. Antibiotic Resistance Crisis: An Update on Antagonistic Interactions between Probiotics and Methicillin-Resistant Staphylococcus aureus (MRSA). Curr. Microbiol. 2021, 78, 2194–2211. [Google Scholar] [CrossRef]
- Archer, N.K.; Mazaitis, M.J.; Costerton, J.W.; Leid, J.G.; Powers, M.E.; Shirtliff, M.E. Staphylococcus aureus biofilms: Properties, Regulation, and Roles in Human Disease. Virulence 2011, 2, 445–459. [Google Scholar] [CrossRef]
- Kaplan, J.B.; Mlynek, K.D.; Hettiarachchi, H.; Alamneh, Y.A.; Biggemann, L.; Zurawski, D.V.; Black, C.C.; Bane, C.E.; Kim, R.K.; Granick, M.S. Extracellular polymeric substance (EPS)-degrading enzymes reduce staphylococcal surface attachment and biocide resistance on pig skin in vivo. PLoS ONE 2018, 13, e0205526. [Google Scholar] [CrossRef]
- Lister, J.L.; Horswill, A.R. Staphylococcus aureus biofilms: Recent developments in biofilm dispersal. Front. Cell. Infect. Microbiol. 2014, 4, 178. [Google Scholar] [CrossRef] [Green Version]
- Schilcher, K.; Horswill, A.R. Staphylococcal Biofilm Development: Structure, Regulation, and Treatment Strategies. Microbiol. Mol. Biol. Rev. 2020, 84, e00026-19. [Google Scholar] [CrossRef]
- Idrees, M.; Sawant, S.; Karodia, N.; Rahman, A. Staphylococcus aureus Biofilm: Morphology, Genetics, Pathogenesis and Treatment Strategies. Int. J. Environ. Res. Public Health 2021, 18, 7602. [Google Scholar] [CrossRef]
- Kranjec, C.; Angeles, D.M.; Mårli, M.T.; Fernández, L.; García, P.; Kjos, M.; Diep, D. Staphylococcal Biofilms: Challenges and Novel Therapeutic Perspectives. Antibiotics 2021, 10, 131. [Google Scholar] [CrossRef]
- Cassat, J.E.; Thomsen, I. Staphylococcus aureus infections in children. Curr. Opin. Infect. Dis. 2021, 34, 510–518. [Google Scholar] [CrossRef]
- Liu, T.; Chen, Y.; Bi, G.; Luo, J.; Du, Z.; Kong, J.; Chen, Y. Generation of Methicillin-Resistant Staphylococcus Aureus Biofilm Infection in an Immunosuppressed Rat Model. J. Pharmacol. Exp. Ther. 2017, 23, 5803–5811. [Google Scholar] [CrossRef] [Green Version]
- Arciola, C.R.; Campoccia, D.; Montanaro, L. Implant infections: Adhesion, biofilm formation and immune evasion. Nat. Rev. Microbiol. 2018, 16, 397–409. [Google Scholar] [CrossRef]
- Balasubramanian, D.; Harper, L.; Shopsin, B.; Torres, V.J. Staphylococcus aureus pathogenesis in diverse host environments. Pathog. Dis. 2017, 75, ftx005. [Google Scholar] [CrossRef] [Green Version]
- Carolus, H.; Van Dyck, K.; Van Dijck, P. Candida albicans and Staphylococcus Species: A Threatening Twosome. Front. Microbiol. 2019, 10, 2162. [Google Scholar] [CrossRef]
- Vila, T.; Kong, E.F.; Montelongo-Jauregui, D.; Van Dijck, P.; Shetty, A.C.; McCracken, C.; Bruno, V.M.; Jabra-Rizk, M.A. Therapeutic implications of C. albicans-S. aureus mixed biofilm in a murine subcutaneous catheter model of polymicrobial infection. Virulence 2021, 12, 835–851. [Google Scholar] [CrossRef] [PubMed]
- Fox, E.P.; Cowley, E.S.; Nobile, C.J.; Hartooni, N.; Newman, D.K.; Johnson, A.D. Anaerobic Bacteria Grow within Candida albicans Biofilms and Induce Biofilm Formation in Suspension Cultures. Curr. Biol. 2014, 24, 2411–2416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, M.E.; Gomes, F.; Rodrigues, C.F. Candida spp./Bacteria Mixed Biofilms. J. Fungi 2019, 6, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayer, F.L.; Wilson, D.; Hube, B. Candida albicanspathogenicity mechanisms. Virulence 2013, 4, 119–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suchodolski, J.; Krasowska, A. Fructose Induces Fluconazole Resistance in Candida albicans through Activation of Mdr1 and Cdr1 Transporters. Int. J. Mol. Sci. 2021, 22, 2127. [Google Scholar] [CrossRef]
- Cavalheiro, M.; Pais, P.; Galocha, M.; Teixeira, M.C. Host-Pathogen Interactions Mediated by MDR Transporters in Fungi: As Pleiotropic as it Gets! Genes 2018, 9, 332. [Google Scholar] [CrossRef] [Green Version]
- Nami, S.; Mohammadi, R.; Vakili, M.; Khezripour, K.; Mirzaei, H.; Morovati, H. Fungal vaccines, mechanism of actions and immunology: A comprehensive review. Biomed. Pharmacother. 2018, 109, 333–344. [Google Scholar] [CrossRef]
- Achkar, J.M.; Fries, B.C. Candida Infections of the Genitourinary Tract. Clin. Microbiol. Rev. 2010, 23, 253–273. [Google Scholar] [CrossRef] [Green Version]
- Ganguly, S.; Mitchell, A.P. Mucosal biofilms of Candida albicans. Curr. Opin. Microbiol. 2011, 14, 380–385. [Google Scholar] [CrossRef] [Green Version]
- Nobile, C.J.; Johnson, A.D. Candida albicans Biofilms and Human Disease. Annu. Rev. Microbiol. 2015, 69, 71–92. [Google Scholar] [CrossRef]
- D’Enfert, C.; Kaune, A.-K.; Alaban, L.-R.; Chakraborty, S.; Cole, N.; Delavy, M.; Kosmala, D.; Marsaux, B.; Fróis-Martins, R.; Morelli, M.; et al. The impact of the Fungus-Host-Microbiota interplay upon Candida albicans infections: Current knowledge and new perspectives. FEMS Microbiol. Rev. 2020, 45, fuaa060. [Google Scholar] [CrossRef]
- Lopes, J.P.; Lionakis, M.S. Pathogenesis and virulence of Candida albicans. Virulence 2021, 13, 89–121. [Google Scholar] [CrossRef]
- Tumbarello, M.; Fiori, B.; Trecarichi, E.M.; Posteraro, P.; Losito, A.R.; De Luca, A.; Sanguinetti, M.; Fadda, G.; Cauda, R.; Posteraro, B. Risk Factors and Outcomes of Candidemia Caused by Biofilm-Forming Isolates in a Tertiary Care Hospital. PLoS ONE 2012, 7, e33705. [Google Scholar] [CrossRef] [Green Version]
- Rajendran, R.; Sherry, L.; Nile, C.J.; Sherriff, A.; Johnson, E.M.; Hanson, M.F.; Williams, C.; Munro, C.A.; Jones, B.J.; Ramage, G. Biofilm formation is a risk factor for mortality in patients with Candida albicans bloodstream infection—Scotland, 2012–2013. Clin. Microbiol. Infect. 2016, 22, 87–93. [Google Scholar] [CrossRef] [Green Version]
- Pfaller, M.; Neofytos, D.; Diekema, D.; Azie, N.; Meier-Kriesche, H.-U.; Quan, S.-P.; Horn, D. Epidemiology and outcomes of candidemia in 3648 patients: Data from the Prospective Antifungal Therapy (PATH Alliance®) registry, 2004–2008. Diagn. Microbiol. Infect. Dis. 2012, 74, 323–331. [Google Scholar] [CrossRef]
- Harriott, M.M.; Noverr, M.C. Candida albicans and Staphylococcus aureus Form Polymicrobial Biofilms: Effects on Antimicrobial Resistance. Antimicrob. Agents Chemother. 2009, 53, 3914–3922. [Google Scholar] [CrossRef] [Green Version]
- Peters, B.M.; Ovchinnikova, E.S.; Krom, B.P.; Schlecht, L.M.; Zhou, H.; Hoyer, L.L.; Busscher, H.J.; van der Mei, H.C.; Jabra-Rizk, M.A.; Shirtliff, M.E. Staphylococcus aureus adherence to Candida albicans hyphae is mediated by the hyphal adhesin Als3p. Microbiology 2012, 158, 2975–2986. [Google Scholar] [CrossRef] [Green Version]
- Kong, E.F.; Tsui, C.; Kucharíková, S.; Andes, D.; Van Dijck, P.; Jabra-Rizk, M.A. Commensal Protection of Staphylococcus aureus against Antimicrobials by Candida albicans Biofilm Matrix. Mbio 2016, 7, e01365-16. [Google Scholar] [CrossRef] [Green Version]
- Kong, E.F.; Kucharíková, S.; Van Dijck, P.; Peters, B.M.; Shirtliff, M.E.; Jabra-Rizk, M.A. Clinical Implications of Oral Candidiasis: Host Tissue Damage and Disseminated Bacterial Disease. Infect. Immun. 2015, 83, 604–613. [Google Scholar] [CrossRef] [Green Version]
- Schlecht, L.M.; Peters, B.M.; Krom, B.P.; Freiberg, J.A.; Hänsch, G.M.; Filler, S.G.; Jabra-Rizk, M.A.; Shirtliff, M.E. Systemic Staphylococcus aureus infection mediated by Candida albicans hyphal invasion of mucosal tissue. Microbiology 2015, 161, 168–181. [Google Scholar] [CrossRef]
- Sharafutdinov, I.S.; Trizna, E.Y.; Baidamshina, D.R.; Ryzhikova, M.N.; Sibgatullina, R.R.; Khabibrakhmanova, A.M.; Latypova, L.Z.; Kurbangalieva, A.R.; Rozhina, E.V.; Klinger-Strobel, M.; et al. Antimicrobial Effects of Sulfonyl Derivative of 2(5H)-Furanone against Planktonic and Biofilm Associated Methicillin-Resistant and -Susceptible Staphylococcus aureus. Front. Microbiol. 2017, 8, 2246. [Google Scholar] [CrossRef] [PubMed]
- Janssens, J.C.A.; Steenackers, H.; Robijns, S.; Gellens, E.; Levin, J.; Zhao, H.; Hermans, K.; De Coster, D.; Verhoeven, T.L.; Marchal, K.; et al. Brominated Furanones Inhibit Biofilm Formation by Salmonella enterica Serovar Typhimurium. Appl. Environ. Microbiol. 2008, 74, 6639–6648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Proctor, C.R.; McCarron, P.A.; Ternan, N.G. Furanone quorum-sensing inhibitors with potential as novel therapeutics against Pseudomonas aeruginosa. J. Med. Microbiol. 2020, 69, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Ren, D.; Sims, J.J.; Wood, T.K. Inhibition of biofilm formation and swarming of Escherichia coli by (5Z)-4-bromo-5-(bromomethylene)-3-butyl-2(5H)-furanone. Environ. Microbiol. 2001, 3, 731–736. [Google Scholar] [CrossRef] [PubMed]
- Ren, D.; Bedzyk, L.A.; Ye, R.W.; Thomas, S.M.; Wood, T.K. Differential gene expression shows natural brominated furanones interfere with the autoinducer-2 bacterial signaling system of Escherichia coli. Biotechnol. Bioeng. 2004, 88, 630–642. [Google Scholar] [CrossRef]
- Ren, D.; Bedzyk, L.A.; Setlow, P.; England, D.F.; Kjelleberg, S.; Thomas, S.M.; Ye, R.W.; Wood, T.K. Differential Gene Expression to Investigate the Effect of (5Z)-4-Bromo- 5-(Bromomethylene)-3-Butyl-2(5H)-Furanone on Bacillus subtilis. Appl. Environ. Microbiol. 2004, 70, 4941–4949. [Google Scholar] [CrossRef] [Green Version]
- Ngwane, A.H.; Panayides, J.-L.; Chouteau, F.; Macingwana, L.; Viljoen, A.; Baker, B.; Madikane, E.; de Kock, C.; Wiesner, L.; Chibale, K.; et al. Design, synthesis, and in vitro antituberculosis activity of 2(5H)-Furanone derivatives. IUBMB Life 2016, 68, 612–620. [Google Scholar] [CrossRef] [Green Version]
- Yujie, L.; Geng, X.; Huang, Y.-C.; Li, Y.; Yang, K.; Ye, L.; Chen, X.; Zhao, G.; Yin, C. The Effect of Brominated Furanones on the Formation of Staphylococcus aureus Biofilm on PVC. Cell Biophys. 2013, 67, 1501–1505. [Google Scholar] [CrossRef]
- Gondela, E.; Walczak, K.Z. Synthesis and preliminary bioactivity assays of 3,4-dichloro-5-(ω-hydroxyalkylamino)-2(5H)-furanones. Eur. J. Med. Chem. 2010, 45, 3993–3997. [Google Scholar] [CrossRef]
- Hentzer, M.; Riedel, K.; Rasmussen, T.B.; Heydorn, A.; Andersen, J.B.; Parsek, M.R.; Rice, S.A.; Eberl, L.; Molin, S.; Høiby, N.; et al. Inhibition of quorum sensing in Pseudomonas aeruginosa biofilm bacteria by a halogenated furanone compound. Microbiology 2002, 148, 87–102. [Google Scholar] [CrossRef]
- Hentzer, M.; Wu, H.; Andersen, J.B.; Riedel, K.; Rasmussen, T.B.; Bagge, N.; Kumar, N.; Schembri, M.; Song, Z.; Kristoffersen, P.; et al. Attenuation of Pseudomonas aeruginosa virulence by quorum sensing inhibitors. EMBO J. 2003, 22, 3803–3815. [Google Scholar] [CrossRef]
- Sharafutdinov, I.S.; Pavlova, A.S.; Khabibrakhmanova, A.M.; Faizova, G.M.; Kurbangalieva, A.R.; Tanaka, K.; Trizna, E.Y.; Baidamshina, D.R.; Bogachev, M.I.; Kayumov, A.R. Targeting Bacillus cereus Cells: Increasing Efficiency of Antimicrobials by the Bornyl-Possessing 2(5H)-furanone Derivative. New Microbiol. 2019, 42, 29–36. [Google Scholar]
- Sharafutdinov, I.S.; Ozhegov, G.D.; Sabirova, A.E.; Novikova, V.V.; Lisovskaya, S.A.; Khabibrakhmanova, A.M.; Kurbangalieva, A.R.; Bogachev, M.I.; Kayumov, A.R. Increasing Susceptibility of Drug-Resistant Candida albicans to Fluconazole and Terbinafine by 2(5H)-Furanone Derivative. Molecules 2020, 25, 642. [Google Scholar] [CrossRef] [Green Version]
- Kayumov, A.; Khakimullina, E.N.; Sharafutdinov, I.; Trizna, E.Y.; Latypova, L.Z.; Lien, H.T.; Margulis, A.B.; Bogachev, M.; Kurbangalieva, A.R. Inhibition of biofilm formation in Bacillus subtilis by new halogenated furanones. J. Antibiot. 2014, 68, 297–301. [Google Scholar] [CrossRef] [Green Version]
- Buchta, V.; Pour, M.; Kubanová, P.; Silva, L.; Votruba, I.; Vopršálová, M.; Schiller, R.; Fáková, H.; Špulák, M. In Vitro Activities of 3-(Halogenated Phenyl)-5-Acyloxymethyl- 2,5-Dihydrofuran-2-ones against Common and Emerging Yeasts and Molds. Antimicrob. Agents Chemother. 2004, 48, 873–878. [Google Scholar] [CrossRef] [Green Version]
- Trizna, E.; Latypova, L.; Kurbangalieva, A.; Bogachev, M.I.; Kayumov, A. 2(5H)-Furanone Derivatives as Inhibitors of Staphylococcal Biofilms. BioNanoScience 2016, 6, 423–426. [Google Scholar] [CrossRef]
- Lonn-Stensrud, J.; Landin, M.A.; Benneche, T.; Petersen, F.C.; Scheie, A.A. Furanones, potential agents for preventing Staphylococcus epidermidis biofilm infections? J. Antimicrob. Chemother. 2008, 63, 309–316. [Google Scholar] [CrossRef] [Green Version]
- Pan, J.; Bahar, A.A.; Syed, H.; Ren, D. Reverting Antibiotic Tolerance of Pseudomonas aeruginosa PAO1 Persister Cells by (Z)-4-bromo-5-(bromomethylene)-3-methylfuran-2(5H)-one. PLoS ONE 2012, 7, e45778. [Google Scholar] [CrossRef] [Green Version]
- Pan, J.; Ren, D. Structural effects on persister control by brominated furanones. Bioorganic Med. Chem. Lett. 2013, 23, 6559–6562. [Google Scholar] [CrossRef]
- Chen, Q.; Huang, B. A Novel Chiral 5((-)-bornyloxy)-3,4-dichloro-2(5 H)-furanone the Efficient Optically Pure Synthesis and Stereospecific Tandem Michael Addition Elimination Reaction. Chin. Chem. Lett. 1993, 4, 675–678. [Google Scholar]
- Leclercq, R.; Cantón, R.; Brown, D.F.J.; Giske, C.G.; Heisig, P.; MacGowan, A.P.; Mouton, J.W.; Nordmann, P.; Rodloff, A.C.; Rossolini, G.M.; et al. EUCAST expert rules in antimicrobial susceptibility testing. Clin. Microbiol. Infect. 2013, 19, 141–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merritt, J.H.; Kadouri, D.E.; O’Toole, G.A. Growing and Analyzing Static Biofilms. Curr. Protoc. Microbiol. 2005, 22, 1B-1. [Google Scholar] [CrossRef]
- Odds, F.C. Synergy, antagonism, and what the chequerboard puts between them. J. Antimicrob. Chemother. 2003, 52, 1. [Google Scholar] [CrossRef]
- Herigstad, B.; Hamilton, M.; Heersink, J. How to optimize the drop plate method for enumerating bacteria. J. Microbiol. Methods 2001, 44, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Kayumov, A.R.; Nureeva, A.A.; Trizna, E.Y.; Gazizova, G.R.; Bogachev, M.; Shtyrlin, N.; Pugachev, M.; Sapozhnikov, S.V.; Shtyrlin, Y. New Derivatives of Pyridoxine Exhibit High Antibacterial Activity against Biofilm-Embedded Staphylococcus Cells. BioMed Res. Int. 2015, 2015, 890968. [Google Scholar] [CrossRef]
- Watamoto, T.; Samaranayake, L.P.; Egusa, H.; Yatani, H.; Seneviratne, C.J. Transcriptional regulation of drug-resistance genes in Candida albicans biofilms in response to antifungals. J. Med. Microbiol. 2011, 60, 1241–1247. [Google Scholar] [CrossRef]
- Jia, W.; Zhang, H.; Li, C.; Li, G.; Liu, X.; Wei, J. The calcineruin inhibitor cyclosporine a synergistically enhances the susceptibility of Candida albicans biofilms to fluconazole by multiple mechanisms. BMC Microbiol. 2016, 16, 113. [Google Scholar] [CrossRef] [Green Version]
- Zheng, L.; Duan, Z.; Tang, D.; He, Y.; Chen, X.; Chen, Q.; Li, M. GP IIb/IIIa-Mediated Platelet Activation and Its Modulation of the Immune Response of Monocytes against Candida albicans. Front. Cell. Infect. Microbiol. 2021, 11, 783085. [Google Scholar] [CrossRef]
- Kurbangalieva, A.R.; Devyatova, N.F.; Bogdanov, A.V.; Berdnikov, E.A.; Mannafov, T.G.; Krivolapov, D.B.; Litvinov, I.A.; Chmutova, G.A. Synthesis of Novel Arylthio Derivatives of Mucochloric Acid. Phosphorus Sulfur Silicon Relat. Elem. 2007, 182, 607–630. [Google Scholar] [CrossRef]
- Latypova, L.Z.; Saigitbatalova, E.S.; Chulakova, D.R.; Lodochnikova, O.A.; Kurbangalieva, A.; Berdnikov, E.A.; Chmutova, G.A. Sulfides, sulfones, and sulfoxides of the furan-2(5H)-one series. synthesis and structure. Russ. J. Org. Chem. 2014, 50, 521–534. [Google Scholar] [CrossRef]
- Khabibrakhmanova, A.M.; Rabbanieva, E.S.; Gerasimova, D.P.; Islamov, D.R.; Latypova, L.Z.; Lodochnikova, O.A.; Kurbangalieva, A.R. Optically Active Bisthioethers and Disulfones Derived from Furan-2(5H)-one and Dithiols: Synthesis and Structure. Russ. J. Org. Chem. 2022, 58, 1160–1169. [Google Scholar] [CrossRef]
- Sharafutdinov, I.S.; Pavlova, A.S.; Akhatova, F.S.; Khabibrakhmanova, A.M.; Rozhina, E.V.; Romanova, Y.J.; Fakhrullin, R.; Lodochnikova, O.A.; Kurbangalieva, A.R.; Bogachev, M.I.; et al. Unraveling the Molecular Mechanism of Selective Antimicrobial Activity of 2(5H)-Furanone Derivative against Staphylococcus aureus. Int. J. Mol. Sci. 2019, 20, 694. [Google Scholar] [CrossRef]
- Huang, D.; Zheng, S.; Zhang, T.; Cheng, Y.-X. Design, synthesis, fungicidal activities and structure–activity relationship studies of (−)-borneol derivatives containing 2-aryl-thiazole scaffold. Bioorganic Med. Chem. Lett. 2021, 45, 128006. [Google Scholar] [CrossRef]
- Zhang, X.; Fang, J.; Li, C.; Zhang, J.; Yang, S.; Deng, B.; Tu, S. Design, Synthesis, and Fungicidal Activities of Indole-Modified Cinnamamide Derivatives. Chem. Biodivers. 2022, 19, e2022009. [Google Scholar] [CrossRef]
- Yang, W.; Chen, X.; Xiuling, Y.; Guo, S.; Wang, Z.; Yu, X. Advances in Pharmacological Activities of Terpenoids. Nat. Prod. Commun. 2020, 15, 1934578X20903555. [Google Scholar] [CrossRef] [Green Version]
- Silva, A.T.M.E.; Pereira, V.V.; Takahashi, J.A.; Silva, R.R.; Duarte, L.P. Microwave-assisted synthesis of borneol esters and their antimicrobial activity. Nat. Prod. Res. 2017, 32, 1714–1720. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence |
|---|---|
| q18s for | 5’ GGATTTACTGAAGACTAACTACTG 3’ |
| q18s rev | 5’ GAACAACAACCGATCCCTAGT 3’ |
| qGAPDH for | 5’ GTCTCCTCTGACTTCAACAGCG 3’ |
| qGAPDH rev | 5’ ACCACCCTGTTGCTGTAGCCAA 3’ |
| qCDR1 for | 5’ GTACTATCCATCAACCATCAGCACTT 3’ |
| qCDR1 rev | 5’ GCCGTTCTTCCACCTTTTTGTA 3’ |
| qCDR1 for | 5’ TGCTGAACCGACAGACTCAGTT 3’ |
| qCDR2 rev | 5’ AAGAGATTGCCAATTGTCCCATA 3’ |
| qMDR1 for | 5’ TCAGTCCGATGTCAGAAAATGC 3’ |
| qMDR2 rev | 5’ GCAGTGGGAATTTGTAGTATGACAA 3’ |
| S. aureus Strains and Isolates | MIC, µg/mL | MBC, µg/mL | ||||||
|---|---|---|---|---|---|---|---|---|
| F131 | Gen (** Ecoff = 2) | Ami (** Ecoff = 16) | BAC * | F131 | Gen | Ami | BAC * | |
| ATCC 29213 (MSSA) | 8 | 4 | 8 | 0.5 | 32 | 16 | 32 | 2 |
| 18 (MSSA) | 8 | 4 | 8 | 0.5 | 32 | 16 | 32 | 2 |
| 25 (MSSA) | 8 | 8 | 8 | 0.5 | 64 | 32 | 32 | 2 |
| 26 (MSSA) | 16 | 8 | 8 | 0.5 | 32 | 32 | 32 | 4 |
| 1053 (MRSA) | 8 | 16 | 32 | 0.5 | 64 | 32 | 32 | 4 |
| 1065 (MRSA) | 8 | 4 | 32 | 0.5 | 32 | 32 | 128 | 2 |
| 1130 (MRSA) | 8 | 8 | 32 | 0.25 | 64 | 16 | 256 | 4 |
| 1145 (MRSA) | 16 | 4 | 64 | 0.25 | 32 | 32 | 256 | 4 |
| 1167 (MRSA) | 8 | 32 | 32 | 0.5 | 32 | 64 | 256 | 2 |
| 1168 (MRSA) | 16 | 16 | 32 | 0.25 | 128 | 16 | 128 | 4 |
| C. albicans Isolates | MIC | MFC | ||||||
|---|---|---|---|---|---|---|---|---|
| F131 | FLC | TRB | BAC | F131 | FLC | TRB | BAC | |
| K4940 FR | 64 | >512 | 128 | 4 | >512 | >512 | 256 | 4 |
| K5050 FR | 64 | >512 | 128 | 4 | >512 | >512 | 256 | 4 |
| K4074 FR | 64 | >512 | 256 | 4 | 256 | >512 | 512 | 4 |
| K3957 FR | 64 | >512 | 128 | 4 | 256 | >512 | 512 | 4 |
| 661FR | 32 | >512 | 256 | 2 | 256 | >512 | 512 | 4 |
| 688 FR | 64 | >512 | 128 | 2 | 128 | >512 | 256 | 4 |
| 701 | 128 | 16 | 128 | 2 | 128 | >512 | 512 | 4 |
| 703 | 32 | 16 | 128 | 4 | 256 | >512 | 256 | 8 |
| 761 | 32 | 64 | 256 | 2 | 256 | >512 | 512 | 8 |
| 762 | 64 | 32 | 128 | 2 | 256 | >512 | 256 | 4 |
| 722 | 32 | 128 | 64 | 4 | 128 | >512 | 256 | 4 |
| 748 | 64 | 128 | 128 | 2 | 128 | >512 | 256 | 4 |
| BPC | BPC (F131) | FIC | FIC (F131) | FICImin | |
|---|---|---|---|---|---|
| Gentamicin | 16 | 16 | 1 | 2 | 0.1875 |
| Amikacin | 16 | 16 | 8 | 4 | 0.75 |
| Benzalkonium chloride | 0.5 | 16 | 0.0625 | 2 | 0.25 |
| BPC | BPC (F131) | FIC | FIC (F131) | FICImin | |
|---|---|---|---|---|---|
| Fluconazole | 512< | 128 | 128 | 16 | 0.375 |
| Terbinafine | 512 | 128 | 256 | 64 | 1 |
| Benzalkonium chloride | 4 | 128 | 0.5 | 8 | 0.1875 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sulaiman, R.; Trizna, E.; Kolesnikova, A.; Khabibrakhmanova, A.; Kurbangalieva, A.; Bogachev, M.; Kayumov, A. Antimicrobial and Biofilm-Preventing Activity of l-Borneol Possessing 2(5H)-Furanone Derivative F131 against S. aureus—C. albicans Mixed Cultures. Pathogens 2023, 12, 26. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens12010026
Sulaiman R, Trizna E, Kolesnikova A, Khabibrakhmanova A, Kurbangalieva A, Bogachev M, Kayumov A. Antimicrobial and Biofilm-Preventing Activity of l-Borneol Possessing 2(5H)-Furanone Derivative F131 against S. aureus—C. albicans Mixed Cultures. Pathogens. 2023; 12(1):26. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens12010026
Chicago/Turabian StyleSulaiman, Rand, Elena Trizna, Alena Kolesnikova, Alsu Khabibrakhmanova, Almira Kurbangalieva, Mikhail Bogachev, and Airat Kayumov. 2023. "Antimicrobial and Biofilm-Preventing Activity of l-Borneol Possessing 2(5H)-Furanone Derivative F131 against S. aureus—C. albicans Mixed Cultures" Pathogens 12, no. 1: 26. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens12010026