
Low Transmission of Chikungunya Virus by Aedes aegypti from Vientiane Capital, Lao PDR
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Virus Strains
2.3. Mosquito Collections
2.4. Mosquito Oral Infections
2.5. Infection, Dissemination, and Transmission Analysis
2.6. Statistical Analysis
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Coffey, L.L.; Beeharry, Y.; Bordería, A.V.; Blanc, H.; Vignuzzi, M. Arbovirus high fidelity variant loses fitness in mosquitoes and mice. Proc. Natl. Acad. Sci. USA 2011, 108, 16038–16043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kraemer, M.U.G.; Sinka, M.E.; Duda, K.A.; Mylne, A.Q.N.; Shearer, F.M.; Barker, C.M.; Moore, C.G.; Carvalho, R.G.; Coelho, G.E.; Van Bortel, W.; et al. The global distribution of the arbovirus vectors Aedes aegypti and Aedes albopictus. eLife 2015, 4, e08347. [Google Scholar] [CrossRef] [PubMed]
- Paupy, C.; Delatte, H.; Bagny, L.; Corbel, V.; Fontenille, D. Aedes albopictus, an arbovirus vector: From the darkness to the light. Microbes Infect. 2009, 11, 1177–1185. [Google Scholar] [CrossRef] [PubMed]
- Kramer, L.D.; Ebel, G.D. Dynamics of flavivirus infection in mosquitoes. Adv. Virus Res. 2003, 60, 187–232. [Google Scholar]
- Rudolph, K.E.; Lessler, J.; Moloney, R.M.; Kmush, B.; Cummings, D.A. Incubation periods of mosquito-borne viral infections: A systematic review. Am. J. Trop. Med. Hyg. 2014, 90, 882–891. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, O.; Albert, M.L. Biology and pathogenesis of chikungunya virus. Nat. Rev. Microbiol. 2010, 8, 491–500. [Google Scholar] [CrossRef]
- Strauss, J.H.; Strauss, E.G. The alphaviruses: Gene expression, replication, and evolution. Microbiol. Rev. 1994, 58, 491–562. [Google Scholar] [CrossRef]
- Weaver, S.C. Arrival of Chikungunya Virus in the New World: Prospects for Spread and Impact on Public Health. PLoS Negl. Trop. Dis. 2014, 8, e2921. [Google Scholar] [CrossRef] [Green Version]
- Volk, S.M.; Chen, R.; Tsetsarkin, K.A.; Adams, A.P.; Garcia, T.I.; Sall, A.A.; Nasar, F.; Schuh, A.J.; Holmes, E.C.; Higgs, S.; et al. Genome-Scale Phylogenetic Analyses of Chikungunya Virus Reveal Independent Emergences of Recent Epidemics and Various Evolutionary Rates. J. Virol. 2010, 84, 6497–6504. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.C. An epidemic of virus disease in Southern Province, Tanganyika territory, in 1952–1953. Trans. R. Soc. Trop. Med. Hyg. 1955, 49, 28–32. [Google Scholar] [CrossRef]
- Lumsden, W.H.R. An epidemic of virus disease in Southern Province, Tanganyika territory, in 1952–1953 II. General description and epidemiology. Trans. R. Soc. Trop. Med. Hyg. 1955, 49, 33–57. [Google Scholar] [CrossRef] [PubMed]
- Amraoui, F.; Failloux, A.B. Chikungunya: An unexpected emergence in Europe. Curr. Opin. Virol. 2016, 21, 146–150. [Google Scholar] [CrossRef] [PubMed]
- Yactayo, S.; Staples, J.E.; Millot, V.; Cibrelus, L.; Ramon-Pardo, P. Epidemiology of Chikungunya in the Americas. J. Infect. Dis. 2016, 214 (Suppl. 5), S441–S445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staples, J.E.; Breiman, R.F.; Powers, A.M. Chikungunya Fever: An Epidemiological Review of a Re-Emerging Infectious Disease. Clin. Infect. Dis. 2009, 49, 942–948. [Google Scholar] [CrossRef] [PubMed]
- Wimalasiri-Yapa, B.M.C.R.; Stassen, L.; Huang, X.; Hafner, L.M.; Hu, W.; Devine, G.; Yakob, L.; Jansen, C.C.; Faddy, H.M.; Viennet, E.; et al. Chikungunya virus in Asia–Pacific: A systematic review. Emerg. Microbes Infect. 2019, 8, 70–79. [Google Scholar] [CrossRef] [Green Version]
- Le, B.C.T.; Ekalaksananan, T.; Thaewnongiew, K.; Phanthanawiboon, S.; Aromseree, S.; Phanitchat, T.; Chuerduangphui, J.; Suwannatrai, A.T.; Alexander, N.; Overgaard, H.J.; et al. Interepidemic Detection of Chikungunya Virus Infection and Transmission in Northeastern Thailand. Am. J. Trop. Med. Hyg. 2020, 103, 1660–1669. [Google Scholar] [CrossRef]
- Díaz-Menéndez, M.; Esteban, E.T.; Ujiie, M.; Calleri, G.; Rothe, C.; Malvy, D.; Nicastri, E.; Bissinger, A.L.; Grandadam, M.; Alpern, J.D.; et al. Travel-associated chikungunya acquired in Myanmar in 2019. Eurosurveillance 2020, 25, 1900721. [Google Scholar] [CrossRef]
- Arif, M.; Tauran, P.; Kosasih, H.; Pelupessy, N.M.; Sennang, N.; Mubin, R.H.; Sudarmono, P.; Tjitra, E.; Murniati, D.; Alam, A.; et al. Chikungunya in Indonesia: Epidemiology and diagnostic challenges. PLoS Negl. Trop. Dis. 2020, 14, e0008355. [Google Scholar] [CrossRef]
- Sharif, N.; Sarkar, M.K.; Ferdous, R.N.; Ahmed, S.N.; Billah, B.; Talukder, A.A.; Zhang, M.; Dey, S.K. Molecular Epidemiology, Evolution and Reemergence of Chikungunya Virus in South Asia. Front. Microbiol. 2021, 12, 1411. [Google Scholar] [CrossRef]
- Calvez, E.; Pommelet, V.; Somlor, S.; Pompon, J.; Viengphouthong, S.; Bounmany, P.; Chindavong, T.A.; Xaybounsou, T.; Prasayasith, P.; Keosenhom, S.; et al. Trends of the Dengue Serotype-4 Circulation with Epidemiological, Phylogenetic, and Entomological Insights in Lao PDR between 2015 and 2019. Pathogens 2020, 9, 728. [Google Scholar] [CrossRef]
- Castonguay-Vanier, J.; Klitting, R.; Sengvilaipaseuth, O.; Piorkowski, G.; Baronti, C.; Sibounheuang, B.; Vongsouvath, M.; Chanthongthip, A.; Thongpaseuth, S.; Mayxay, M.; et al. Molecular epidemiology of dengue viruses in three provinces of Lao PDR, 2006–2010. PLoS Negl. Trop. Dis. 2018, 12, e0006203. [Google Scholar] [CrossRef] [PubMed]
- Lao, M.; Caro, V.; Thiberge, J.-M.; Bounmany, P.; Vongpayloth, K.; Buchy, P.; Duong, V.; Vanhlasy, C.; Hospied, J.-M.; Thongsna, M.; et al. Co-Circulation of Dengue Virus Type 3 Genotypes in Vientiane Capital, Lao PDR. PLoS ONE 2014, 9, e115569. [Google Scholar] [CrossRef] [PubMed]
- Somlor, S.; Vongpayloth, K.; Diancourt, L.; Buchy, P.; Duong, V.; Phonekeo, D.; Ketmayoon, P.; Vongphrachanh, P.; Brey, P.T.; Caro, V.; et al. Chikungunya virus emergence in the Lao PDR, 2012–2013. PLoS ONE 2017, 12, e0189879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcombe, S.; Fustec, B.; Cattel, J.; Chonephetsarath, S.; Thammavong, P.; Phommavanh, N.; David, J.-P.; Corbel, V.; Sutherland, I.W.; Hertz, J.C.; et al. Distribution of insecticide resistance and mechanisms involved in the arbovirus vector Aedes aegypti in Laos and implication for vector control. PLoS Negl. Trop. Dis. 2019, 13, e0007852. [Google Scholar] [CrossRef] [Green Version]
- Tangena, J.-A.A.; Marcombe, S.; Thammavong, P.; Chonephetsarath, S.; Somphong, B.; Sayteng, K.; Grandadam, M.; Sutherland, I.W.; Lindsay, S.W.; Brey, P.T. Bionomics and insecticide resistance of the arboviral vector Aedes albopictus in northern Lao PDR. PLoS ONE 2018, 13, e0206387. [Google Scholar] [CrossRef] [Green Version]
- Calvez, E.; Vetsaphong, P.; Somlor, S.; Xaybounsou, T.; Viengphouthong, S.; Dupont-Rouzeyrol, M.; Pommelet, V.; Brey, P.T. First probable case of congenital Zika syndrome in Lao People’s Democratic Republic. Int. J. Infect. Dis. 2021, 105, 595–597. [Google Scholar] [CrossRef]
- Chanthavy, S.; Phouthone, S.; Khonesavanh, P.; Darouny, P.; Sonesavanh, P.; Khamphaphongphane, B.; Viengsavanh, K.; Lewis, H.C.; Soulaphy, C.; Souliphone, P.; et al. Emergence of chikungunya in Moonlapamok and Khong Districts, Champassak Province, the Lao People’s Democratic Republic, May to September 2012. West. Pac. Surveill. Response J. 2013, 4, 46–50. [Google Scholar] [CrossRef]
- Phommanivong, V.; Kanda, S.; Shimono, T.; Lamaningao, P.; Darcy, A.W.; Mishima, N.; Phaytanavanh, B.; Nishiyama, T. Co-circulation of the dengue with chikungunya virus during the 2013 outbreak in the southern part of Lao PDR. Trop. Med. Health 2016, 44, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Kanamitsu, M.; Taniguchi, K.; Urasawa, S.; Ogata, T.; Wada, Y.; Saroso, J.S. Geographic Distribution of Arbovirus Antibodies in Indigenous Human Populations in the Indo-Australian Archipelago. Am. J. Trop. Med. Hyg. 1979, 28, 351–363. [Google Scholar] [CrossRef] [Green Version]
- Calvez, E.; Bounmany, P.; Somlor, S.; Xaybounsou, T.; Viengphouthong, S.; Keosenhom, S.; Brey, P.T.; Lacoste, V.; Grandadam, M. Multiple chikungunya virus introductions in Lao PDR from 2014 to 2020. PLoS ONE 2022, 17, e0271439. [Google Scholar] [CrossRef]
- Vazeille, M.; Moutailler, S.; Coudrier, D.; Rousseaux, C.; Khun, H.; Huerre, M.; Thiria, J.; Dehecq, J.-S.; Fontenille, D.; Schuffenecker, I.; et al. Two Chikungunya Isolates from the Outbreak of La Reunion (Indian Ocean) Exhibit Different Patterns of Infection in the Mosquito, Aedes albopictus. PLoS ONE 2007, 2, e1168. [Google Scholar] [CrossRef] [PubMed]
- Galun, R.; Avi-Dor, Y.; Bar-Zeev, M. Feeding Response in Aedes aegypti: Stimulation by Adenosine Triphosphate. Science 1963, 142, 1674–1675. [Google Scholar] [CrossRef]
- Pastorino, B.; Bessaud, M.; Grandadam, M.; Murri, S.; Tolou, H.J.; Peyrefitte, C.N. Development of a TaqMan® RT-PCR assay without RNA extraction step for the detection and quantification of African Chikungunya viruses. J. Virol. Methods 2005, 124, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression. Thousand Oaks; Sage: New York, NY, USA, 2011. [Google Scholar]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous Inference in General Parametric Models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Foundation for Statistical Computing V Austria. R Core Team. R, A Language and Environment for Statistical Computing. 2016. Available online: https://www.R-project.org/ (accessed on 21 November 2021).
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis Internet; Springer: New York, NY, USA, 2016; Available online: https://ggplot2.tidyverse.org (accessed on 21 November 2021).
- Wickham, H. The Split-Apply-Combine Strategy for Data Analysis. J. Stat. Softw. 2011, 40, 1–29. [Google Scholar] [CrossRef] [Green Version]
- Dubot-Pérès, A.; Vongphrachanh, P.; Denny, J.; Phetsouvanh, R.; Linthavong, S.; Sengkeopraseuth, B.; Khasing, A.; Xaythideth, V.; Moore, C.; Vongsouvath, M.; et al. An Epidemic of Dengue-1 in a Remote Village in Rural Laos. PLoS Negl. Trop. Dis. 2013, 7, e2360. [Google Scholar] [CrossRef] [Green Version]
- Souza-Neto, J.A.; Powell, J.R.; Bonizzoni, M. Aedes aegypti vector competence studies: A review. Infect. Genet. Evol. 2019, 67, 191–209. [Google Scholar] [CrossRef]
- Franz, A.W.E.; Kantor, A.M.; Passarelli, A.L.; Clem, R.J. Tissue Barriers to Arbovirus Infection in Mosquitoes. Viruses 2015, 7, 3741–3767. [Google Scholar] [CrossRef] [Green Version]
- Weaver, S.C.; Forrester, N.L.; Liu, J.; Vasilakis, N. Population bottlenecks and founder effects: Implications for mosquito-borne arboviral emergence. Nat. Rev. Microbiol. 2021, 19, 184–195. [Google Scholar] [CrossRef]
- De Guilhem de Lataillade, L.; Vazeille, M.; Obadia, T.; Madec, Y.; Mousson, L.; Kamgang, B.; Chen, C.-H.; Failloux, A.-B.; Yen, P.-S. Risk of yellow fever virus transmission in the Asia-Pacific region. Nat. Commun. 2020, 11, 5801. [Google Scholar] [CrossRef]
- Miot, E.F.; Calvez, E.; Aubry, F.; Dabo, S.; Grandadam, M.; Marcombe, S.; Oke, C.; Logan, J.G.; Brey, P.T.; Lambrechts, L. Risk of arbovirus emergence via bridge vectors: Case study of the sylvatic mosquito Aedes malayensis in the Nakai district, Laos. Sci. Rep. 2020, 10, 7750. [Google Scholar] [CrossRef] [PubMed]
- Ko, H.-Y.; Salem, G.M.; Chang, G.-J.J.; Chao, D.-Y. Application of Next-Generation Sequencing to Reveal How Evolutionary Dynamics of Viral Population Shape Dengue Epidemiology. Front. Microbiol. 2020, 11, 1371. [Google Scholar] [CrossRef] [PubMed]
- Lambrechts, L.; Chevillon, C.; Albright, R.G.; Thaisomboonsuk, B.; Richardson, J.H.; Jarman, R.G.; Scott, T.W. Genetic specificity and potential for local adaptation between dengue viruses and mosquito vectors. BMC Evol. Biol. 2009, 9, 160. [Google Scholar] [CrossRef] [Green Version]
- Vogels, C.B.F.; Rückert, C.; Cavany, S.M.; Perkins, T.A.; Ebel, G.D.; Grubaugh, N.D. Arbovirus coinfection and co-transmission: A neglected public health concern? PLoS Biol. 2019, 17, e3000130. [Google Scholar] [CrossRef] [Green Version]
- Le Coupanec, A.; Tchankouo-Nguetcheu, S.; Roux, P.; Khun, H.; Huerre, M.; Morales-Vargas, R.; Enguehard, M.; Lavillette, D.; Missé, D.; Choumet, V. Co-Infection of Mosquitoes with Chikungunya and Dengue Viruses Reveals Modulation of the Replication of Both Viruses in Midguts and Salivary Glands of Aedes aegypti Mosquitoes. Int. J. Mol. Sci. 2017, 18, 1708. [Google Scholar] [CrossRef] [Green Version]
- Rückert, C.; Weger-Lucarelli, J.; Garcia-Luna, S.M.; Young, M.C.; Byas, A.; Murrieta, R.A.; Fauver, J.R.; Ebel, G.D. Impact of simultaneous exposure to arboviruses on infection and transmission by Aedes aegypti mosquitoes. Nat. Commun. 2017, 8, 15412. [Google Scholar] [CrossRef] [Green Version]
- Vega-Rúa, A.; Zouache, K.; Girod, R.; Failloux, A.-B.; Lourenço-De-Oliveira, R. High Level of Vector Competence of Aedes aegypti and Aedes albopictus from Ten American Countries as a Crucial Factor in the Spread of Chikungunya Virus. J. Virol. 2014, 88, 6294–6306. [Google Scholar] [CrossRef] [Green Version]
- Vega-Rúa, A.; Lourenço-De-Oliveira, R.; Mousson, L.; Vazeille, M.; Fuchs, S.; Yébakima, A.; Gustave, J.; Girod, R.; Dusfour, I.; Leparc-Goffart, I.; et al. Chikungunya Virus Transmission Potential by Local Aedes Mosquitoes in the Americas and Europe. PLoS Negl. Trop. Dis. 2015, 9, e0003780. [Google Scholar] [CrossRef] [Green Version]
- Dupont-Rouzeyrol, M.; Caro, V.; Guillaumot, L.; Vazeille, M.; D’Ortenzio, E.; Thiberge, J.-M.; Baroux, N.; Gourinat, A.-C.; Grandadam, M.; Failloux, A.-B. Chikungunya Virus and the Mosquito Vector Aedes aegypti in New Caledonia (South Pacific Region). Vector-Borne Zoonotic Dis. 2012, 12, 1036–1041. [Google Scholar] [CrossRef] [Green Version]
- Kramer, L.D.; Ciota, A.T. Dissecting vectorial capacity for mosquito-borne viruses. Curr. Opin. Virol. 2015, 15, 112–118. [Google Scholar] [CrossRef] [Green Version]
- Garrett-Jones, C.; Grab, B. The Assessment of Insecticidal Impact on the Malaria Mosquito’s Vectorial Capacity, from Data on the Proportion of Parous Females. Bull. World Health Organ. 1964, 31, 71–86. [Google Scholar] [PubMed]
- Lequime, S.; Paul, R.E.; Lambrechts, L.; Paul, R. Determinants of Arbovirus Vertical Transmission in Mosquitoes. PLoS Pathog. 2016, 12, e1005548. [Google Scholar] [CrossRef] [PubMed]
- Mendenhall, I.H.; Manuel, M.; Moorthy, M.; Lee, T.T.M.; Low, D.H.W.; Missé, D.; Gubler, D.J.; Ellis, B.R.; Ooi, E.E.; Pompon, J. Peridomestic Aedes malayensis and Aedes albopictus are capable vectors of arboviruses in cities. PLoS Negl. Trop. Dis. 2017, 11, e0005667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vega-Rúa, A.; Marconcini, M.; Madec, Y.; Manni, M.; Carraretto, D.; Gomulski, L.M.; Gasperi, G.; Failloux, A.-B.; Malacrida, A.R. Vector competence of Aedes albopictus populations for chikungunya virus is shaped by their demographic history. Commun. Biol. 2020, 3, 326. [Google Scholar] [CrossRef]
- Grandadam, M.; Caro, V.; Plumet, S.; Thiberge, J.M.; Souarès, Y.; Failloux, A.B.; Tolou, H.J.; Budelot, M.; Cosserat, D.; Leparc-Goffart, I.; et al. Chikungunya Virus, Southeastern France. Emerg. Infect. Dis. 2011, 17, 910–913. [Google Scholar] [CrossRef]
- Khongwichit, S.; Chansaenroj, J.; Thongmee, T.; Benjamanukul, S.; Wanlapakorn, N.; Chirathaworn, C.; Poovorawan, Y. Large-scale outbreak of Chikungunya virus infection in Thailand, 2018–2019. PLoS ONE 2021, 16, e0247314. [Google Scholar] [CrossRef]
- Puntasecca, C.J.; King, C.H.; LaBeaud, A.D. Measuring the global burden of chikungunya and Zika viruses: A systematic review. PLoS Negl. Trop. Dis. 2021, 15, e0009055. [Google Scholar] [CrossRef]
- Kounnavong, S.; Gopinath, D.; Hongvanthong, B.; Khamkong, C.; Sichanthongthip, O. Malaria elimination in Lao PDR: The challenges associated with population mobility. Infect. Dis. Poverty 2017, 6, 81. [Google Scholar] [CrossRef] [Green Version]
- Pongvongsa, T.; Nonaka, D.; Iwagami, M.; Soundala, P.; Khattignavong, P.; Xangsayarath, P.; Nishimoto, F.; Kobayashi, J.; Hongvanthon, B.; Brey, P.T.; et al. Malaria among foreign migrant workers in Savannakhet Province, Lao People’s Democratic Republic. Trop. Med. Health 2019, 47, 10. [Google Scholar] [CrossRef]
- Coker, R.J.; Hunter, B.; Rudge, J.W.; Liverani, M.; Hanvoravongchai, P. Emerging infectious diseases in southeast Asia: Regional challenges to control. Lancet 2011, 377, 599–609. [Google Scholar] [CrossRef]
- Gutiérrez-Bugallo, G.; Boullis, A.; Martinez, Y.; Hery, L.; Rodríguez, M.; Bisset, J.A.; Vega-Rúa, A. Vector competence of Aedes aegypti from Havana, Cuba, for dengue virus type 1, chikungunya, and Zika viruses. PLoS Negl. Trop. Dis. 2020, 14, e0008941. [Google Scholar] [CrossRef] [PubMed]
- Richard, V.; Paoaafaite, T.; Cao-Lormeau, V.-M. Vector Competence of Aedes aegypti and Aedes polynesiensis Populations from French Polynesia for Chikungunya Virus. PLoS Negl. Trop. Dis. 2016, 10, e0004694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agha, S.B.; Chepkorir, E.; Mulwa, F.; Tigoi, C.; Arum, S.; Guarido, M.M.; Ambala, P.; Chelangat, B.; Lutomiah, J.; Tchouassi, D.P.; et al. Vector competence of populations of Aedes aegypti from three distinct cities in Kenya for chikungunya virus. PLoS Negl. Trop. Dis. 2017, 11, e0005860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viglietta, M.; Bellone, R.; Blisnick, A.A.; Failloux, A.-B. Vector Specificity of Arbovirus Transmission. Front. Microbiol. 2021, 12, 773211. [Google Scholar] [CrossRef] [PubMed]


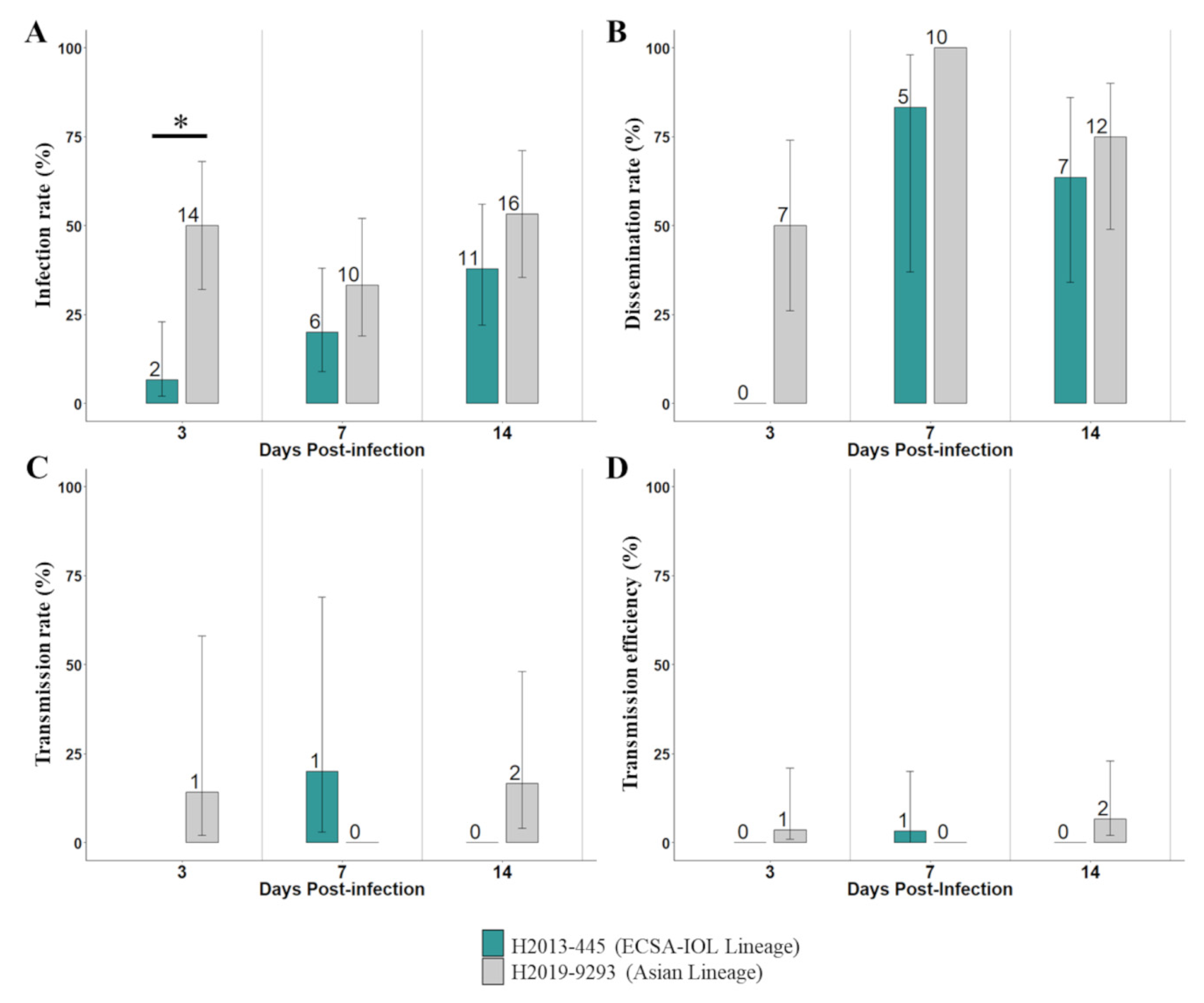{kind=link}
| Infection Rate | Dissemination Rate | |||||
| LR χ2 | Df | p Value | LR χ2 | Df | p Value | |
| Virus | 14.81 | 1 | 0.0001 | 2.522 | 1 | 0.1123 |
| dpi | 9.083 | 2 | 0.0107 | 5.181 | 2 | 0.075 |
| Virus × dpi | 5.674 | 2 | 0.0586 | 2.488 | 2 | 0.2882 |
| Transmission rate | Transmission efficiency | |||||
| LR χ2 | Df | p Value | LR χ2 | Df | p Value | |
| Virus | 1.475 | 1 | 0.1258 | 1.475 | 1 | 0.2245 |
| dpi | 2.197 | 1 | 0.1703 | 2.197 | 2 | 0.3333 |
| Virus × dpi | 4.578 | 1 | 0.0379 | 4.578 | 2 | 0.1014 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Calvez, E.; Miot, E.F.; Keosenhom, S.; Vungkyly, V.; Viengphouthong, S.; Bounmany, P.; Brey, P.T.; Marcombe, S.; Grandadam, M. Low Transmission of Chikungunya Virus by Aedes aegypti from Vientiane Capital, Lao PDR. Pathogens 2023, 12, 31. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens12010031
Calvez E, Miot EF, Keosenhom S, Vungkyly V, Viengphouthong S, Bounmany P, Brey PT, Marcombe S, Grandadam M. Low Transmission of Chikungunya Virus by Aedes aegypti from Vientiane Capital, Lao PDR. Pathogens. 2023; 12(1):31. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens12010031
Chicago/Turabian StyleCalvez, Elodie, Elliott F. Miot, Sitsana Keosenhom, Vaekey Vungkyly, Souksakhone Viengphouthong, Phaithong Bounmany, Paul T. Brey, Sébastien Marcombe, and Marc Grandadam. 2023. "Low Transmission of Chikungunya Virus by Aedes aegypti from Vientiane Capital, Lao PDR" Pathogens 12, no. 1: 31. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens12010031