
Seroprevalence of Tick-Borne Encephalitis (TBE) Virus Antibodies in Wild Rodents from Two Natural TBE Foci in Bavaria, Germany
 , ,
, , 
Abstract
:1. Introduction
2. Results
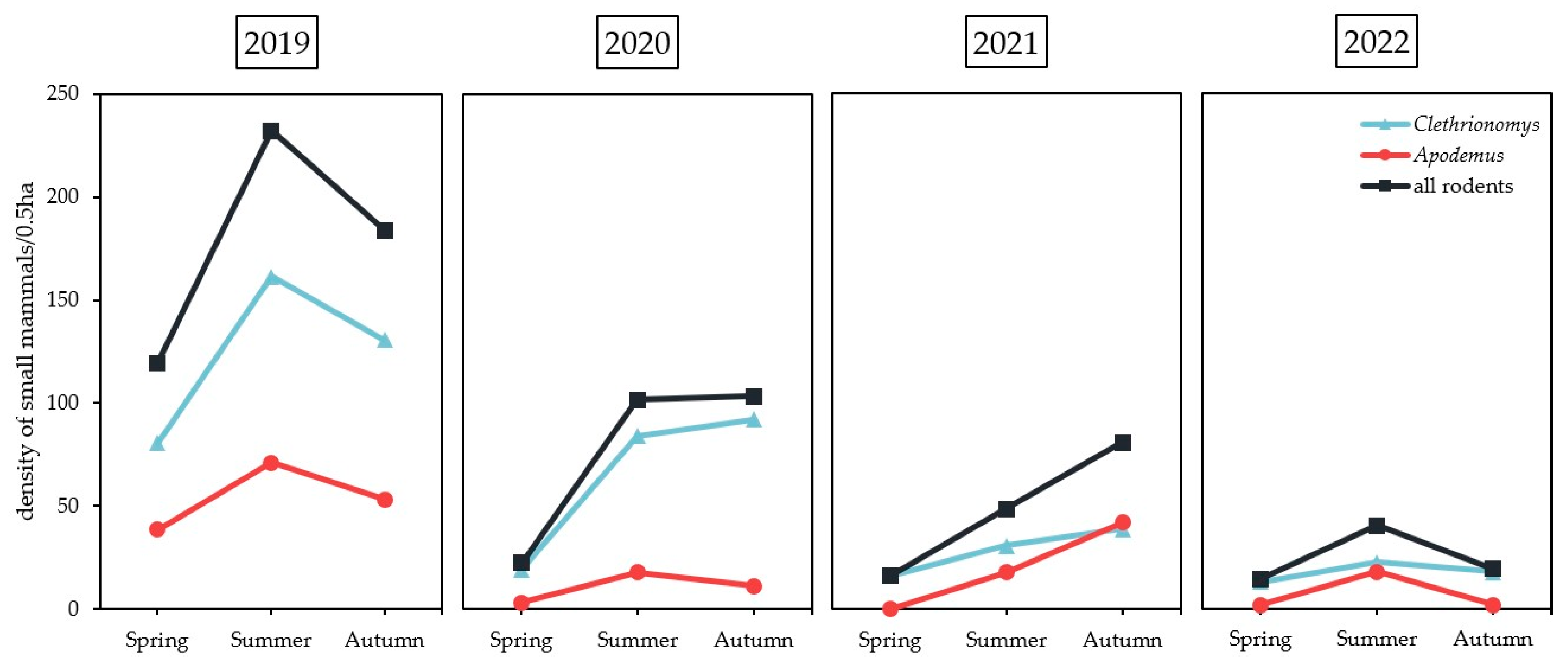
2.1. Small Mammal Trapping
2.2. TBEV Seroprevalence in Small Mammals
2.3. Movement Profile of the Recaptured Small Mammals
3. Discussion
4. Materials and Methods
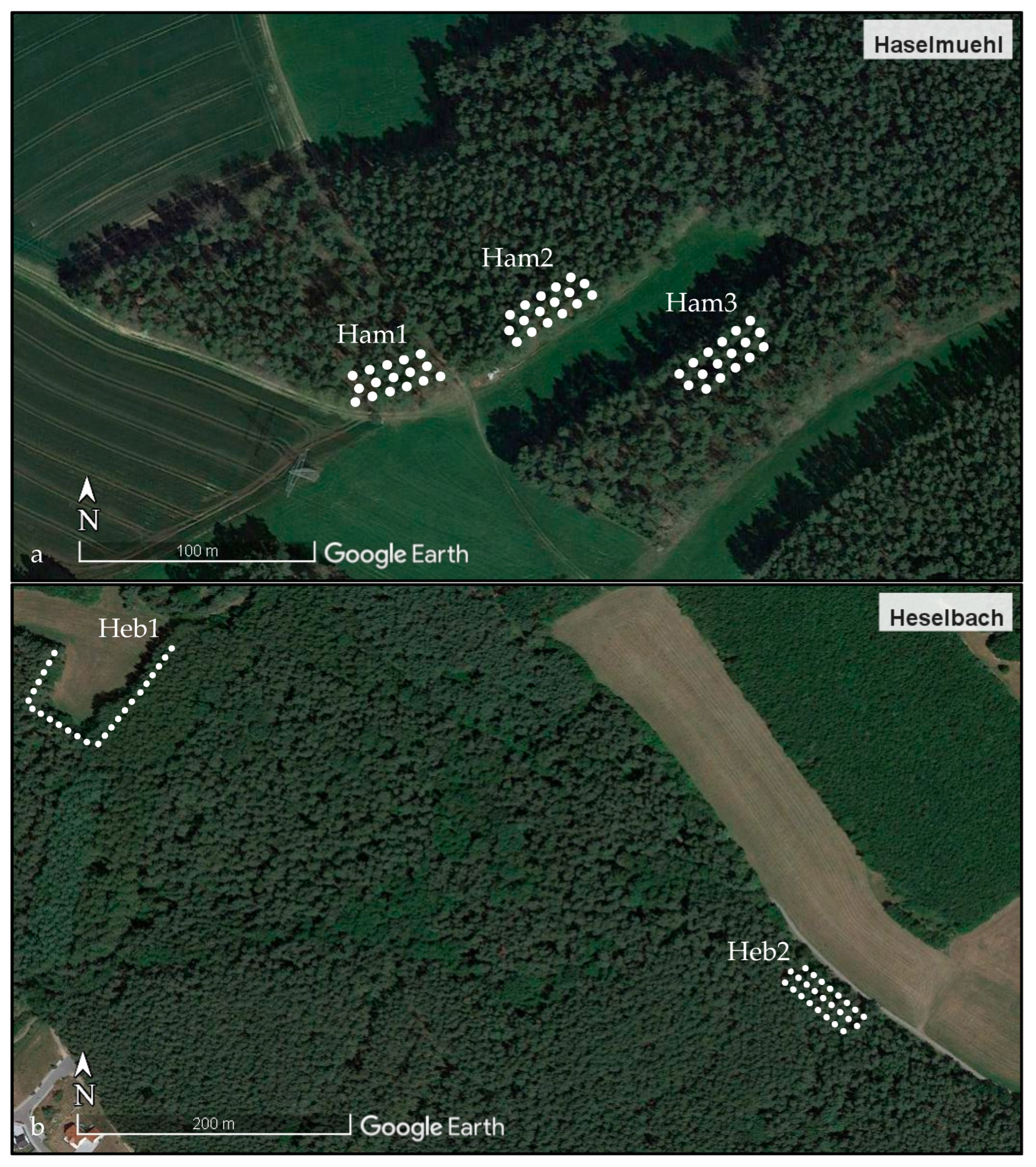
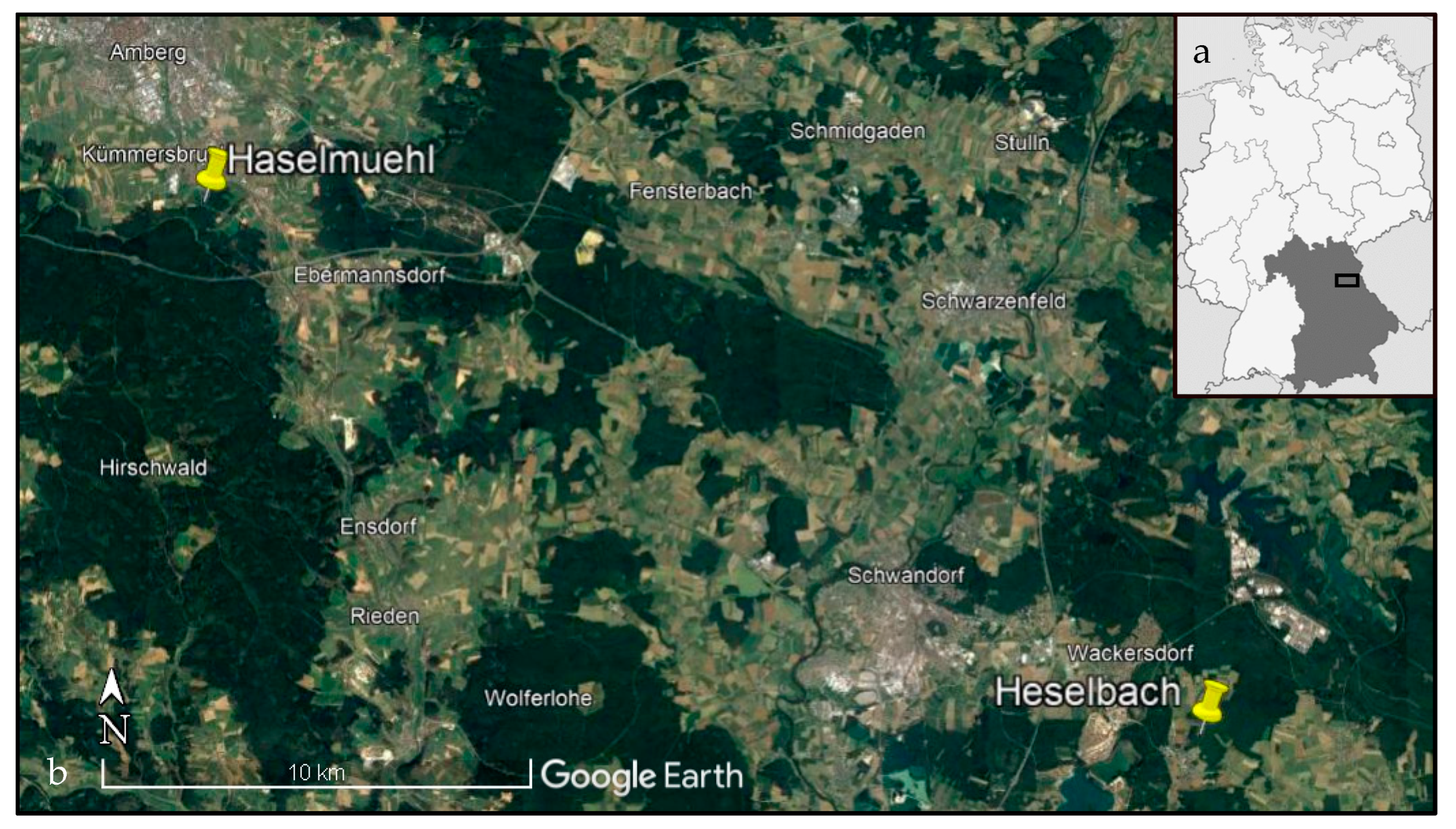
4.1. Study Area
4.2. Small Mammal Trapping and Sampling
4.3. Ethical Statement
4.4. Serological Analysis
4.4.1. Indirect Immunofluorescence Assay (IIFA)
4.4.2. Serum Neutralization Test (SNT)
4.5. Statistical Analysis
4.5.1. Definition
4.5.2. Statistical Analyses of TBEV Seroprevalence in Small Mammals
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lindquist, L.; Vapalahti, O. Tick-borne encephalitis. Lancet 2008, 371, 1861–1871. [Google Scholar] [CrossRef]
- Mansfield, K.L.; Johnson, N.; Phipps, L.P.; Stephenson, J.R.; Fooks, A.R.; Solomon, T. Tick-borne encephalitis virus—A review of an emerging zoonosis. J. Gen. Virol. 2009, 90, 1781–1794. [Google Scholar] [CrossRef]
- Dobler, G.; Gniel, D.; Petermann, R.; Pfeffer, M. Epidemiology and distribution of tick-borne encephalitis. Wien. Med. Wochenschr. 2012, 162, 230–238. [Google Scholar] [CrossRef] [PubMed]
- Amato-Gauci, A.J.; Zeller, H. Tick-borne encephalitis joins the diseases under surveillance in the European Union. Eurosurveillance 2012, 17, 20299. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control. Tick-borne encephalitis—Annual Epidemiological Report for 2020. Available online: https://www.ecdc.europa.eu/en/publications-data/tick-borne-encephalitis-annual-epidemiological-report-2020 (accessed on 3 November 2022).
- Robert Koch-Institut. Epidemiologisches Bulletin; Robert-Koch-Institut: Berlin/Wernigerode, Germany, 2022; Volume 9, pp. 3–19.
- Deviatkin, A.A.; Karganova, G.G.; Vakulenko, Y.A.; Lukashev, A.N. TBEV Subtyping in Terms of Genetic Distance. Viruses 2020, 12, 1240. [Google Scholar] [CrossRef]
- Ruzek, D.; Salát, J.; Palus, M.; Gritsun, T.S.; Gould, E.A.; Dyková, I.; Skallová, A.; Jelínek, J.; Kopecký, J.; Grubhoffer, L. CD8+ T-cells mediate immunopathology in tick-borne encephalitis. Virology 2009, 384, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Valarcher, J.F.; Hägglund, S.; Juremalm, M.; Blomqvist, G.; Chirico, J. Tick-borne encephalitis. Rev. Sci. Tech. 2015, 34, 453–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bogovic, P.; Strle, F. Tick-borne encephalitis: A review of epidemiology, clinical characteristics, and management. WJCC 2015, 3, 430–441. [Google Scholar] [CrossRef]
- Borde, J.P.; Glaser, R.; Braun, K.; Riach, N.; Hologa, R.; Kaier, K.; Chitimia-Dobler, L.; Dobler, G. Decoding the Geography of Natural TBEV Microfoci in Germany: A Geostatistical Approach Based on Land-Use Patterns and Climatological Conditions. Int. J. Environ. Res. Public Health 2022, 19, 11830. [Google Scholar] [CrossRef]
- Labuda, M.; Randolph, S.E. Survival strategy of tick-borne encephalitis virus: Cellular basis and environmental determinants. Zent. Für Bakteriol. 1999, 289, 513–524. [Google Scholar] [CrossRef]
- Randolph, S.E. Transmission of tick-borne pathogens between co-feeding ticks: Milan Labuda’s enduring paradigm. Ticks Tick-Borne Dis. 2011, 2, 179–182. [Google Scholar] [CrossRef] [PubMed]
- Karbowiak, G.; Biernat, B. The role of particular tick developmental stages in the circulation of tick-borne pathogens affecting humans in Central Europe. 2. Tick-borne encephalitis virus. Ann. Parasitol. 2016, 62, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Tonteri, E.; Jääskeläinen, A.E.; Tikkakoski, T.; Voutilainen, L.; Niemimaa, J.; Henttonen, H.; Vaheri, A.; Vapalahti, O. Tick-borne encephalitis virus in wild rodents in winter, Finland, 2008–2009. Emerg. Infect. Dis. 2011, 17, 72–75. [Google Scholar] [CrossRef] [PubMed]
- Bakhvalova, V.N.; Dobrotvorsky, A.K.; Panov, V.V.; Matveeva, V.A.; Tkachev, S.E.; Morozova, O.V. Natural tick-borne encephalitis virus infection among wild small mammals in the southeastern part of western Siberia, Russia. Vector Borne Zoonotic Dis. 2006, 6, 32–41. [Google Scholar] [CrossRef]
- Kerlik, J.; Avdičová, M.; Musilová, M.; Bérešová, J.; Mezencev, R. Breast Milk as Route of Tick-Borne Encephalitis Virus Transmission from Mother to Infant. Emerg. Infect. Dis. 2022, 28, 1060–1061. [Google Scholar] [CrossRef]
- Achazi, K.; Růžek, D.; Donoso-Mantke, O.; Schlegel, M.; Ali, H.S.; Wenk, M.; Schmidt-Chanasit, J.; Ohlmeyer, L.; Rühe, F.; Vor, T.; et al. Rodents as sentinels for the prevalence of tick-borne encephalitis virus. Vector Borne Zoonotic Dis. 2011, 11, 641–647. [Google Scholar] [CrossRef] [Green Version]
- Lipowski, D.; Popiel, M.; Perlejewski, K.; Nakamura, S.; Bukowska-Osko, I.; Rzadkiewicz, E.; Dzieciatkowski, T.; Milecka, A.; Wenski, W.; Ciszek, M.; et al. A Cluster of Fatal Tick-borne Encephalitis Virus Infection in Organ Transplant Setting. J. Infect. Dis. 2017, 215, 896–901. [Google Scholar] [CrossRef] [Green Version]
- Weidmann, M.; Schmidt, P.; Hufert, F.T.; Krivanec, K.; Meyer, H. Tick-borne encephalitis virus in Clethrionomys glareolus in the Czech Republic. Vector Borne Zoonotic Dis. 2006, 6, 379–381. [Google Scholar] [CrossRef]
- Knap, N.; Korva, M.; Dolinšek, V.; Sekirnik, M.; Trilar, T.; Avšič-Županc, T. Patterns of tick-borne encephalitis virus infection in rodents in Slovenia. Vector Borne Zoonotic Dis. 2012, 12, 236–242. [Google Scholar] [CrossRef]
- Zöldi, V.; Papp, T.; Reiczigel, J.; Egyed, L. Bank voles show high seropositivity rates in a natural TBEV focus in Hungary. Infect. Dis. 2015, 47, 178–181. [Google Scholar] [CrossRef]
- Michelitsch, A.; Fast, C.; Sick, F.; Tews, B.A.; Stiasny, K.; Bestehorn-Willmann, M.; Dobler, G.; Beer, M.; Wernike, K. Long-term presence of tick-borne encephalitis virus in experimentally infected bank voles (Myodes glareolus). Ticks Tick-Borne Dis. 2021, 12, 101693. [Google Scholar] [CrossRef] [PubMed]
- Michelitsch, A.; Tews, B.A.; Klaus, C.; Bestehorn-Willmann, M.; Dobler, G.; Beer, M.; Wernike, K. In Vivo Characterization of Tick-Borne Encephalitis Virus in Bank Voles (Myodes glareolus). Viruses 2019, 11, 1069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grzybek, M.; Alsarraf, M.; Tołkacz, K.; Behnke-Borowczyk, J.; Biernat, B.; Stańczak, J.; Strachecka, A.; Guz, L.; Szczepaniak, K.; Paleolog, J.; et al. Seroprevalence of TBEV in bank voles from Poland-a long-term approach. Emerg. Microbes Infect. 2018, 7, 145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bournez, L.; Umhang, G.; Moinet, M.; Richomme, C.; Demerson, J.-M.; Caillot, C.; Devillers, E.; Boucher, J.-M.; Hansmann, Y.; Boué, F.; et al. Tick-Borne Encephalitis Virus: Seasonal and Annual Variation of Epidemiological Parameters Related to Nymph-to-Larva Transmission and Exposure of Small Mammals. Pathogens 2020, 9, 518. [Google Scholar] [CrossRef]
- Borde, J.P.; Kaier, K.; Hehn, P.; Matzarakis, A.; Frey, S.; Bestehorn, M.; Dobler, G.; Chitimia-Dobler, L. The complex interplay of climate, TBEV vector dynamics and TBEV infection rates in ticks-Monitoring a natural TBEV focus in Germany, 2009–2018. PLoS ONE 2021, 16, e0244668. [Google Scholar] [CrossRef]
- Weidmann, M.; Frey, S.; Freire, C.C.M.; Essbauer, S.; Růžek, D.; Klempa, B.; Zubrikova, D.; Vögerl, M.; Pfeffer, M.; Hufert, F.T.; et al. Molecular phylogeography of tick-borne encephalitis virus in central Europe. J. Gen. Virol. 2013, 94, 2129–2139. [Google Scholar] [CrossRef]
- Robert Koch-Institut. SurvStat@RKI 2.0. Available online: https://survstat.rki.de/Content/Query/Create.aspx (accessed on 4 November 2022).
- Zöldi, V.; Papp, T.; Rigó, K.; Farkas, J.; Egyed, L. A 4-year study of a natural tick-borne encephalitis virus focus in Hungary, 2010–2013. EcoHealth 2015, 12, 174–182. [Google Scholar] [CrossRef]
- Viro, P.; Niethammer, J. Clethrionomys glareolus (Schreber, 1780)—Rötelmaus. In Handbuch der Säugetiere Europas; Niethammer, J., Krapp, F., Eds.; Akademische Verlagsgesellschaft: Wiesbaden, Germany, 1982. [Google Scholar]
- Torre, I.; Arrizabalaga, A. Habitat preferences of the bank vole Myodes glareolus in a Mediterranean mountain range. Acta Theriol. 2008, 53, 241–250. [Google Scholar] [CrossRef]
- Litzba, N.; Zelená, H.; Kreil, T.R.; Niklasson, B.; Kühlmann-Rabens, I.; Remoli, M.E.; Niedrig, M. Evaluation of different serological diagnostic methods for tick-borne encephalitis virus: Enzyme-linked immunosorbent, immunofluorescence, and neutralization assay. Vector Borne Zoonotic Dis. 2014, 14, 149–159. [Google Scholar] [CrossRef]
- Girl, P.; Haut, M.; Riederer, S.; Pfeffer, M.; Dobler, G. Comparison of Three Serological Methods for the Epidemiological Investigation of TBE in Dogs. Microorganisms 2021, 9, 399. [Google Scholar] [CrossRef]
- Gilbert, L.; Jones, L.D.; Hudson, P.J.; Gould, E.A.; Reid, H.W. Role of small mammals in the persistence of Louping-ill virus: Field survey and tick co-feeding studies. Med. Vet. Entomol. 2000, 14, 277–282. [Google Scholar] [CrossRef] [PubMed]
- Burri, C.; Korva, M.; Bastic, V.; Knap, N.; Avsic-Zupanc, T.; Gern, L. Serological evidence of tick-borne encephalitis virus infection in rodents captured at four sites in Switzerland. J. Med. Entomol. 2012, 49, 436–439. [Google Scholar] [CrossRef] [Green Version]
- Zeman, P.; Januška, J. Epizootiologic background of dissimilar distribution of human cases of Lyme borreliosis and tick-borne encephalitis in a joint endemic area. Comp. Immunol. Microbiol. Infect. Dis. 1999, 22, 247–260. [Google Scholar] [CrossRef] [PubMed]
- Kozuch, O.; Gresíková, M.; Nosek, J.; Lichard, M.; Sekeyová, M. The role of small rodents and hedgehogs in a natural focus of tick-borne encephalitis. Bull. World Health Organ. 1967, 36, 61–66. [Google Scholar] [PubMed]
- Kozuch, O.; Labuda, M.; Lysý, J.; Weismann, P.; Krippel, E. Longitudinal study of natural foci of Central European encephalitis virus in West Slovakia. Acta Virol. 1990, 34, 537–544. [Google Scholar] [PubMed]
- Kocianová, E.; Kozuch, O.; Bakoss, P.; Rehácek, J.; Kovácová, E. The prevalence of small terrestrial mammals infected with tick-borne encephalitis virus and leptospirae in the foothills of the southern Bavarian forest, Germany. Appl. Parasitol. 1993, 34, 283–290. [Google Scholar]
- Dizij, A.; Kurtenbach, K. Clethrionomys glareolus, but not Apodemus flavicollis, acquires resistance to Ixodes ricinus L., the main European vector of Borrelia burgdorferi. Parasite Immunol. 1995, 17, 177–183. [Google Scholar] [CrossRef]
- Heigl, Z.; von Zeipel, G. Experimental Infection with Tick-Borne Encephalitis Virus in Clethrionomys glareolus, Apodemus flavicollis, Apodemus sylvaticur and Mus musculus. I. Virological Studies. Acta Pathol. Microbiol. Scand. 1966, 66, 489–509. [Google Scholar] [CrossRef]
- Von Zeipel, G.; Heigl, Z. Experimental infection with tick-borne virus in Clethrionomys glareolus, Apodemus flavicollis, Apodemus sylvaticus and Mus musculus. 2. Serological studies. Acta Pathol. Microbiol. Scand. 1966, 68, 211–229. [Google Scholar] [CrossRef]
- Labuda, M.; Nuttall, P.A.; Kozuch, O.; Elecková, E.; Williams, T.; Zuffová, E.; Sabó, A. Non-viraemic transmission of tick-borne encephalitis virus: A mechanism for arbovirus survival in nature. Experientia 1993, 49, 802–805. [Google Scholar] [CrossRef]
- Radda, A.; LOEW, J.; Pretzmann, G. Investigations in a Natural Focus of Early Summer Meningoencephalitis in Lower Austria. 2. Attempt to isolate Virus from Arthropods and Small Mammals. Zent. Fur Bakteriol. Parasitenkd. Infekt. Und Hyg. 1963, 190, 281–298. [Google Scholar]
- Hughes, V.L.; Randolph, S.E. Testosterone increases the transmission potential of tick-borne parasites. Parasitology 2001, 123, 365–371. [Google Scholar] [CrossRef]
- Grzybek, M.; Bajer, A.; Bednarska, M.; Al-Sarraf, M.; Behnke-Borowczyk, J.; Harris, P.D.; Price, S.J.; Brown, G.S.; Osborne, S.-J.; Siński, E.; et al. Long-term spatiotemporal stability and dynamic changes in helminth infracommunities of bank voles (Myodes glareolus) in NE Poland. Parasitology 2015, 142, 1722–1743. [Google Scholar] [CrossRef]
- Labuda, M.; Kozuch, O.; Zuffová, E.; Elecková, E.; Hails, R.S.; Nuttall, P.A. Tick-borne encephalitis virus transmission between ticks cofeeding on specific immune natural rodent hosts. Virology 1997, 235, 138–143. [Google Scholar] [CrossRef] [Green Version]
- Stenseth, N.C.; VIljugrein, H.; Jędrzejewski, W.; Mysterud, A.; Pucek, Z. Population dynamics of Clethrionomys glareolus and Apodemus flavicollis: Seasonal components of density dependence and density independence. Acta Theriol. 2002, 47, 39–67. [Google Scholar] [CrossRef]
- Vukićević-Radić, O.; Mati, R.; Kataranovski, D.; Stamenković, S. Spatial organization and home range of Apodemus flavicollis and A. agrarius on Mt. Avala, Serbia. Acta Zool. Acad. Sci. Hung. 2006, 52, 81–96. [Google Scholar]
- Bujalska, G.; Grüm, L. Social organization of the bank vole (Clethrionomys glareolus, Schreber 1780) and its demographic consequences: A model. Oecologia 1989, 80, 70–81. [Google Scholar] [CrossRef] [PubMed]
- Bajer, A.; Bednarska, M.; Pawełczyk, A.; Behnke, J.M.; Gilbert, F.S.; Sinski, E. Prevalence and abundance of Cryptosporidium parvum and giardia spp. in wild rural rodents from the Mazury Lake District region of Poland. Parasitology 2002, 125, 21–34. [Google Scholar] [CrossRef] [PubMed]
- Cullinane, A.A.; Garvey, M. A review of diagnostic tests recommended by the World Organisation for Animal Health Manual of Diagnostic Tests and Vaccines for Terrestrial Animals. Rev. Sci. Tech. 2021, 40, 75–89. [Google Scholar] [CrossRef] [PubMed]
- Coroian, M.; Mihalca, A.D.; Dobler, G.; Euringer, K.; Girl, P.; Borșan, S.-D.; Kalmár, Z.; Tincuța Briciu, V.; Flonta, M.; Topan, A.; et al. Seroprevalence Rates against West Nile, Usutu, and Tick-Borne Encephalitis Viruses in Blood-Donors from North-Western Romania. Int. J. Environ. Res. Public Health 2022, 19, 8182. [Google Scholar] [CrossRef] [PubMed]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Soft. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Length, R.; Singmann, H.; Love, J.; Buerkner, P.; Herve, M. Emmeans: Estimated marginal means, aka least-squares means. R Package Version 2018, 1, 3. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Haselmuehl | Heselbach | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Clethrionomys glareolus | Apodemus flavicollis | Clethrionomys glareolus | Apodemus flavicollis | ||||||||||||||
| Year | Season | NTR | Seropos. (Seropos./ Sampled Rodents | RF (%) | NTR | Seropos. (Seropos./ Sampled Rodents | RF (%) | NTR | Seropos. (Seropos./ Sampled Rodents | RF (%) | NTR | Seropos. (Seropos./ Sampled Rodents | RF (%) | ||||
| Y | S | Y | S | Y | S | Y | S | ||||||||||
| 2019 | Spring | 34 | 7/30 | 21.3 | 23.3 | 11 | 0/10 | 0 | 0 | 28 | 7/26 | 18.0 | 26.9 | 13 | 4/10 | 22.6 | 40.0 |
| Summer | 72 | 14/59 | 23.7 | 12 | 0/11 | 0 | 49 | 5/32 | 15.6 | 38 | 8/33 | 24.2 | |||||
| Autumn | 52 | 9/52 | 17.3 | 17 | 0/17 | 0 | 42 | 6/42 | 14.3 | 21 | 2/19 | 10.5 | |||||
| 2020 | Spring | 0 | 0/0 | 10.8 | 0 | 0 | 0/0 | 0 | 0 | 7 | 3/7 | 23.9 | 42.9 | 0 | 0/0 | 20.0 | 0 |
| Summer | 19 | 2/19 | 10.5 | 5 | 0/5 | 0 | 49 | 14/44 | 31.8 | 11 | 3/11 | 27.3 | |||||
| Autumn | 19 | 2/18 | 11.1 | 3 | 0/2 | 0 | 41 | 5/41 | 12.2 | 4 | 0/4 | 0 | |||||
| 2021 | Spring | 2 | 0/2 | 17.6 | 0 | 0 | 0/0 | 5.6 | 0 | 11 | 4/11 | 18.8 | 36.4 | 0 | 0/0 | 0 | 0 |
| Summer | 2 | 1/2 | 50.0 | 1 | 0/1 | 0 | 22 | 3/22 | 13.6 | 11 | 0/11 | 0 | |||||
| Autumn | 13 | 2/13 | 15.4 | 17 | 1/17 | 5.9 | 15 | 2/15 | 13.3 | 16 | 0/16 | 0 | |||||
| 2022 | Spring | 6 | 0/6 | 33.3 | 0 | 0 | 0/0 | 0 | 0 | 3 | 0/3 | 0 | 0 | 1 | 0/1 | 7.7 | 0 |
| Summer | 9 | 5/9 | 55.6 | 1 | 0/1 | 0 | 6 | 0/6 | 0 | 11 | 1/11 | 9.1 | |||||
| Autumn | 0 | 0/0 | 0 | 0 | 0/0 | 0 | 11 | 0/11 | 0 | 1 | 0/1 | 0 | |||||
| Total | 228 | 42/210 | 20.0 | 67 | 1/64 | 1.6 | 284 | 49/260 | 18.8 | 127 | 18/117 | 15.4 | |||||
| Factor | Estimate | Std. Error | z-Value | p-Value |
|---|---|---|---|---|
| Total | ||||
| Intercept | −2.9372 | 0.3989 | −7.363 | 1.8 × 10-13 *** |
| A. flavicollis v. Cl. glareolus | 1.0603 | 0.3037 | 3.491 | 0.000481 *** |
| Autumn v. spring | 0.6671 | 0.3517 | 1.897 | 0.057849 . |
| Autumn v. summer | 0.6463 | 0.2745 | 2.355 | 0.018545 * |
| Haselmuehl v. Heselbach | 0.2883 | 0.2356 | 1.224 | 0.221094 |
| 2019 v. 2020 | −0.1045 | 0.2798 | −0.374 | 0.708743 |
| 2019 v. 2021 | −0.5708 | 0.3552 | −1.607 | 0.108080 |
| 2019 v. 2022 | −0.7571 | 0.4762 | −1.590 | 0.111845 |
| Adult v. juvenile | −0.6716 | 0.3426 | −1.960 | 0.049969 * |
| Female v. male | 0.8102 | 0.2509 | 3.229 | 0.001240 ** |
| Haselmuehl | ||||
| Intercept | −5.88806 | 1.16148 | −5.069 | 3.99 × 10−7 *** |
| Autumn v. spring | 0.04609 | 0.58515 | 0.079 | 0.93722 |
| Autumn v. summer | 0.54246 | 0.43445 | 1.249 | 0.21181 |
| A. flavicollis v. Cl. glareolus | 3.44187 | 1.05740 | 3.255 | 0.00113 ** |
| 2019 v. 2020 | −0.65451 | 0.61951 | −1.056 | 0.29074 |
| 2019 v. 2021 | 0.51385 | 0.66947 | 0.768 | 0.44276 |
| 2019 v. 2022 | 0.09561 | 0.68152 | 0.140 | 0.88843 |
| Adult v. juvenile | 0.41223 | 0.51703 | 0.797 | 0.42528 |
| Female v. male | 2.11164 | 0.47819 | 4.416 | 1.01 × 10−5 *** |
| Heselbach | ||||
| Intercept | −1.73161 | 0.49816 | −3.476 | 0.000509 *** |
| Autumn v. spring | 1.30624 | 0.44224 | 2.954 | 0.003140 ** |
| Autumn v. summer | 0.81817 | 0.37229 | 2.198 | 0.027974 * |
| A. flavicollis v. Cl. glareolus | 0.28740 | 0.34491 | 0.833 | 0.404698 |
| 2019 v. 2020 | 0.23872 | 0.35038 | 0.681 | 0.495677 |
| 2019 v. 2021 | −0.78207 | 0.43313 | −1.806 | 0.070976 |
| 2019 v. 2022 | −2.01711 | 1.04650 | −1.927 | 0.053920 . |
| Adult v. juvenile | −1.31092 | 0.44867 | −2.922 | 0.003480 ** |
| Female v. male | 0.09002 | 0.31743 | 0.284 | 0.776736 |
| Apodemus flavicollis | ||||
| Intercept | −3.9593 | 1.1871 | −3.335 | 0.000852 *** |
| Autumn v. spring | 1.4047 | 0.9242 | 1.520 | 0.128515 |
| Autumn v. summer | 1.0298 | 0.7217 | 1.427 | 0.153607 |
| Haselmuehl v. Heselbach | 2.7111 | 1.0687 | 2.537 | 0.011184 * |
| 2019 v. 2020 | −0.5673 | 0.7925 | −0.716 | 0.474066 |
| 2019 v. 2021 | −1.8405 | 1.1040 | −1.667 | 0.095504 |
| 2019 v. 2022 | −1.7362 | 1.1382 | −1.525 | 0.127161 |
| Adult v. juvenile | −2.3298 | 1.1194 | −2.081 | 0.037411 * |
| Female v. male | −1.2373 | 0.5951 | −2.079 | 0.037597 * |
| Clethrionomys glareolus | ||||
| Intercept | −1.935855 | 0.330835 | −5.851 | 4.87 × 10−9 *** |
| Autumn v. spring | 0.524291 | 0.361727 | 1.449 | 0.1472 |
| Autumn v. summer | 0.494920 | 0.288599 | 1.715 | 0.0864 |
| Haselmuehl v. Heselbach | −0.084538 | 0.266944 | −0.317 | 0.7515 |
| 2019 v. 2020 | 0.001682 | 0.310668 | 0.005 | 0.9957 |
| 2019 v. 2021 | −0.268220 | 0.399430 | −0.672 | 0.5019 |
| 2019 v. 2022 | −0.796784 | 0.540969 | −1.473 | 0.1408 |
| Female v. male | 1.363863 | 0.289338 | 4.714 | 2.43 × 10−6 *** |
| TBEV Seropositive/Sampled Rodents | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Haselmuehl | Heselbach | ||||||||||||||||||
| Clethrionomys glareolus | Apodemus flavicollis | Clethrionomys glareolus | Apodemus flavicollis | ||||||||||||||||
| Year | Season | M | F | Juv. | M | F | Juv. | Total | Seropos. (%) | M | F | Juv. | M | F | Juv. | Total | Seropos. (%) | ||
| Y | S | Y | S | ||||||||||||||||
| 2019 | Spring | 3/5 | 2/19 | 2/6 | 0/8 | 0/2 | 0/0 | 7/40 | 16.8 | 17.5 | 2/7 | 5/16 | 0/3 | 1/2 | 3/5 | 0/3 | 11/36 | 19.8 | 30.6 |
| Summer | 9/15 | 2/17 | 3/27 | 0/7 | 0/4 | 0/0 | 14/70 | 20.0 | 3/5 | 0/11 | 2/16 | 3/18 | 4/8 | 1/7 | 13/65 | 20.0 | |||
| Autumn | 3/13 | 1/14 | 5/25 | 0/8 | 0/8 | 0/1 | 9/69 | 13.0 | 1/6 | 3/12 | 2/24 | 2/10 | 0/6 | 0/3 | 8/61 | 13.1 | |||
| 2020 | Spring | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 9.1 | 0 | 1/4 | 0/0 | 2/3 | 0/0 | 0/0 | 0/0 | 3/7 | 23.4 | 42.9 |
| Summer | 2/3 | 0/15 | 0/1 | 0/1 | 0/4 | 0/0 | 2/24 | 8.3 | 8/13 | 6/19 | 0/12 | 0/3 | 3/7 | 0/1 | 17/55 | 30.9 | |||
| Autumn | 1/3 | 0/8 | 1/7 | 0/1 | 0/1 | 0/0 | 2/20 | 10.0 | 3/8 | 1/8 | 1/25 | 0/3 | 0/1 | 0/0 | 5/45 | 11.1 | |||
| 2021 | Spring | 0/0 | 0/0 | 0/2 | 0/0 | 0/0 | 0/0 | 0/2 | 11.4 | 0 | 0/5 | 4/6 | 0/0 | 0/0 | 0/0 | 0/0 | 4/11 | 12.0 | 36.4 |
| Summer | 1/1 | 0/1 | 0/0 | 0/0 | 0/1 | 0/0 | 1/3 | 33.3 | 2/10 | 1/6 | 0/6 | 0/6 | 0/4 | 0/1 | 3/33 | 9.1 | |||
| Autumn | 1/1 | 1/4 | 0/8 | 0/5 | 1/3 | 0/9 | 3/30 | 10.0 | 2/3 | 0/3 | 0/9 | 0/8 | 0/4 | 0/4 | 2/31 | 6.5 | |||
| 2022 | Spring | 0/4 | 0/2 | 0/0 | 0/0 | 0/0 | 0/0 | 0/6 | 31.3 | 0 | 0/2 | 0/1 | 0/0 | 0/0 | 0/1 | 0/0 | 0/4 | 3.0 | 0 |
| Summer | 4/4 | 1/5 | 0/0 | 0/0 | 0/1 | 0/0 | 5/10 | 50.0 | 0/1 | 0/2 | 0/3 | 0/4 | 1/5 | 0/2 | 1/17 | 5.9 | |||
| Autumn | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0 | 0/1 | 0/4 | 0/6 | 0/1 | 0/0 | 0/0 | 0/12 | 0 | |||
| Total | 24/49 | 7/85 | 11/76 | 0/30 | 1/24 | 0/10 | 43/274 | 22/65 | 20/88 | 7/107 | 6/55 | 11/41 | 1/21 | 67/377 | |||||
| Seropos. (%) | 49.0 | 8.2 | 14.5 | 0 | 4.2 | 0 | 15.7 | 33.8 | 22.7 | 6.5 | 10.9 | 26.8 | 4.8 | 17.8 | |||||
| ID | Species | Sex | First Capture | Second Capture | Third Capture | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Sample | IIFA | Days upon 2nd Capture | Sample | IIFA | Days upon 3rd Capture | Sample | IIFA | |||
| Seroconversion | ||||||||||
| 35_Heb | Cl glareolus. | f | serum | neg. | 42 | serum | pos. | |||
| 26_Heb | Cl. glareolus | f | serum | neg. | 43 | lavage | pos. | |||
| 31_Heb | Cl. glareolus | f | serum | neg. | 42 | lavage | pos. | |||
| 220_Heb | Cl. glareolus | f | serum | neg. | 293 | serum | pos. | |||
| 169_Heb | A. flavicollis | m | serum | neg. | 59 | serum | pos. | |||
| 250_Heb | A. flavicollis | f | serum | neg. | 308 | lavage | pos. | |||
| 224_Heb | Cl. glareolus | f | serum | neg. | 47 | serum | neg. | 142 | serum | pos. |
| 194_Heb | Cl. glareolus | m | n.a. | n.a. | 23 | serum | neg. | 293 | serum | pos. |
| 222_Heb | Cl. glareolus | m | serum | neg. | 47 | serum | neg. | 246 | serum | pos. |
| 59_Ham | Cl. glareolus | m | serum | neg. | 48 | serum | pos. | 36 | serum | pos. |
| 297_Heb | Cl. glareolus | f | serum | neg. | 294 | serum | pos. | 37 | serum | pos. |
| 275_Heb | A. flavicollis | f | serum | neg. | 244 | serum | pos. | 20 | serum | pos. |
| 465_Ham | Cl. glareolus | m | serum | neg. | 62 | serum | pos. | 29 | serum | pos. |
| Positive recaptured once | ||||||||||
| 20_Ham | Cl. glareolus | m | serum | pos. | 92 | serum | pos. | |||
| 21_Ham | Cl. glareolus | m | serum | pos. | 92 | serum | pos. | |||
| 146_Ham | Cl. glareolus | m | serum | pos. | 59 | serum | pos. | |||
| 84_Ham | Cl. glareolus | m | serum | pos. | 92 | serum | pos. | |||
| 105_Ham | Cl. glareolus | f | serum | pos. | 59 | serum | pos. | |||
| 232_Heb | Cl. glareolus | f | serum | pos. | 48 | serum | pos. | |||
| 388_Heb | Cl. glareolus | m | serum | pos. | 29 | serum | pos. | |||
| 426_Heb | Cl. glareolus | m | serum | pos. | 30 | serum | pos. | |||
| 290_Heb | Cl. glareolus | f | serum | pos. | 17 | serum | pos. | |||
| 261_Heb | Cl. glareolus | m | serum | pos. | 141 | serum | pos. | |||
| 283_Heb | Cl. glareolus | f | serum | pos. | 17 | lavage | pos. | |||
| Positive recaptured twice | ||||||||||
| 16_Ham | Cl. glareolus | f | serum | pos. | 42 | n.a. | n.a. | 50 | serum | pos. |
| 9_Ham | Cl. glareolus | m | serum | pos. | 91 | serum | pos. | 36 | serum | pos. |
| 80_Heb | Cl. glareolus | m | serum | pos. | 33 | serum | pos. | 59 | serum | pos. |
| 232_Heb | Cl. glareolus | f | serum | pos. | 47 | serum | pos. | 142 | serum | pos. |
| 294_Heb | Cl. glareolus | m | serum | pos. | 15 | serum | pos. | 21 | serum | pos. |
| 305_Heb | Cl. glareolus | m | serum | pos. | 20 | serum | pos. | 36 | serum | pos. |
| 385_Heb | Cl. glareolus | f | serum | pos. | 36 | serum | pos. | 28 | serum | pos. |
| 77_Heb | A. flavicollis | f | serum | pos. | 33 | serum | pos. | 36 | serum | pos. |
| Seropositive to seronegative captures | ||||||||||
| 189_Ham | Cl. glareolus | f | n.a. | n.a. | 23 | serum | pos. | 48 | serum | neg. |
| 136_Heb | A. flavicollis | m | serum | pos. | 36 | lavage | neg. | |||
| 69_Heb | A. flavicollis | m | serum | pos. | 16 | n.a. | n.a. | 91 | serum | neg. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brandenburg, P.J.; Obiegala, A.; Schmuck, H.M.; Dobler, G.; Chitimia-Dobler, L.; Pfeffer, M. Seroprevalence of Tick-Borne Encephalitis (TBE) Virus Antibodies in Wild Rodents from Two Natural TBE Foci in Bavaria, Germany. Pathogens 2023, 12, 185. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens12020185
Brandenburg PJ, Obiegala A, Schmuck HM, Dobler G, Chitimia-Dobler L, Pfeffer M. Seroprevalence of Tick-Borne Encephalitis (TBE) Virus Antibodies in Wild Rodents from Two Natural TBE Foci in Bavaria, Germany. Pathogens. 2023; 12(2):185. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens12020185
Chicago/Turabian StyleBrandenburg, Philipp Johannes, Anna Obiegala, Hannah Maureen Schmuck, Gerhard Dobler, Lidia Chitimia-Dobler, and Martin Pfeffer. 2023. "Seroprevalence of Tick-Borne Encephalitis (TBE) Virus Antibodies in Wild Rodents from Two Natural TBE Foci in Bavaria, Germany" Pathogens 12, no. 2: 185. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens12020185