
Development of Bioluminescent Virulent Aeromonas hydrophila for Understanding Pathogenicity

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Growth Conditions, and Plasmids
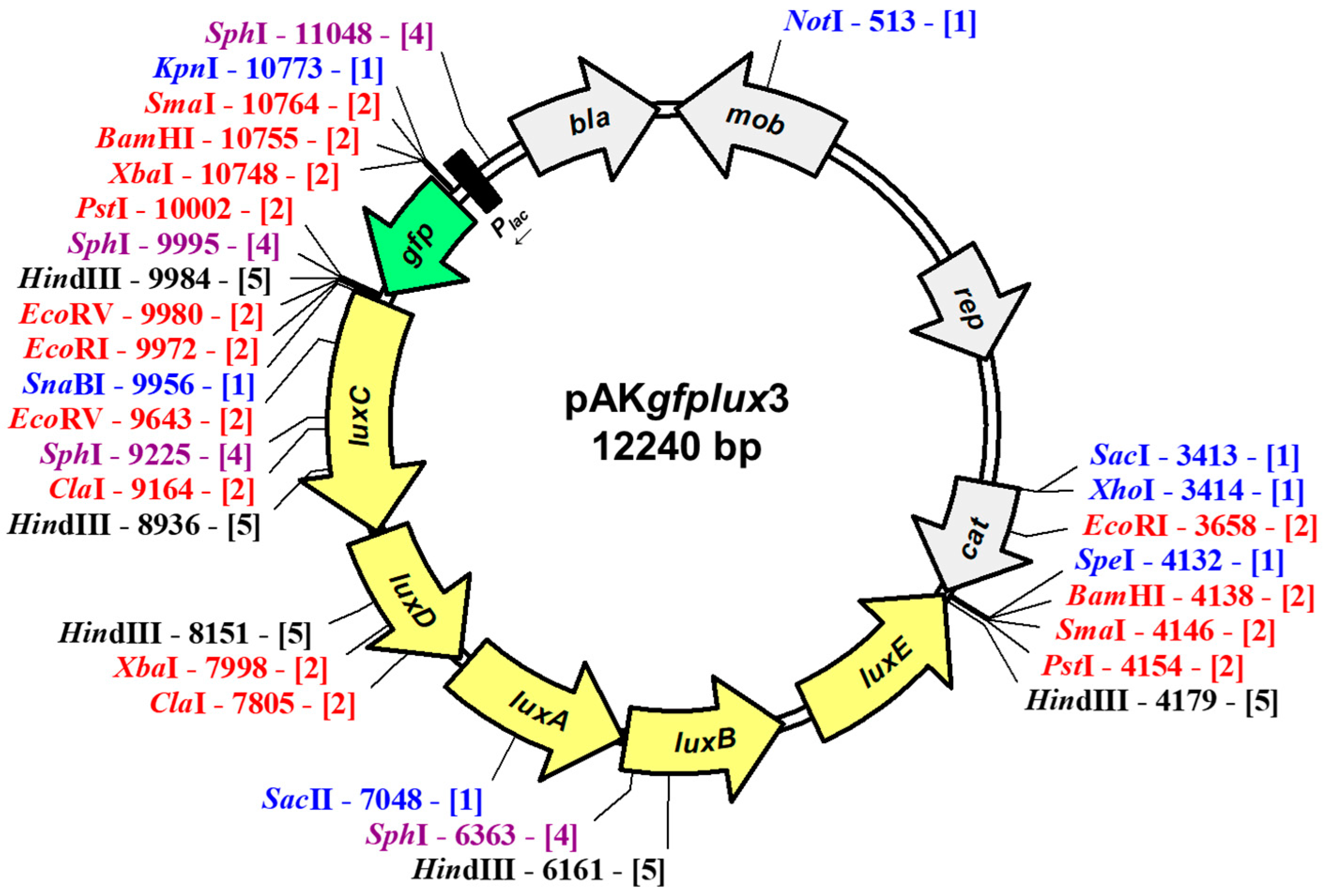
2.2. Construction of pAKgfplux3
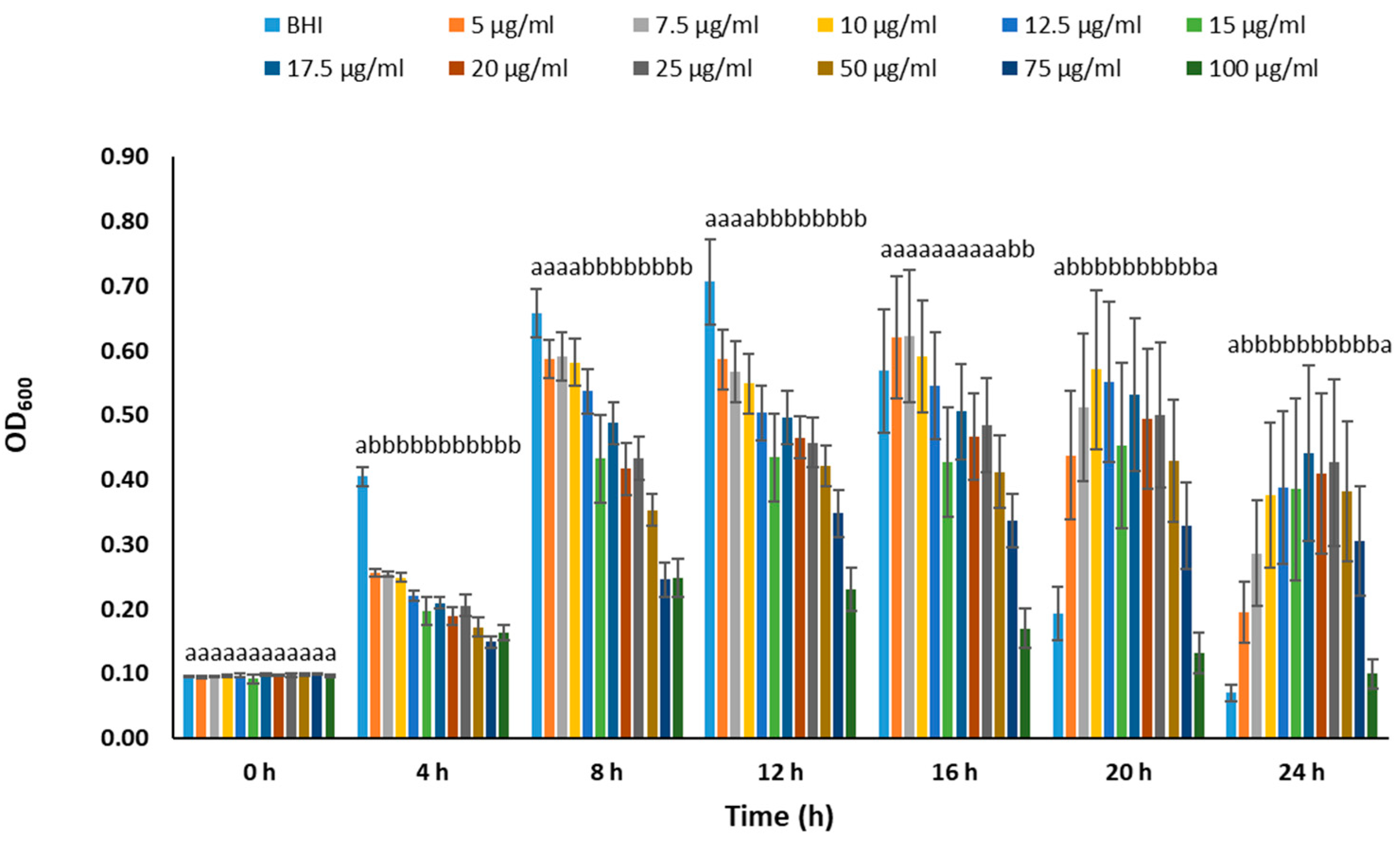
2.3. Determination of Optimal Chloramphenicol Concentration
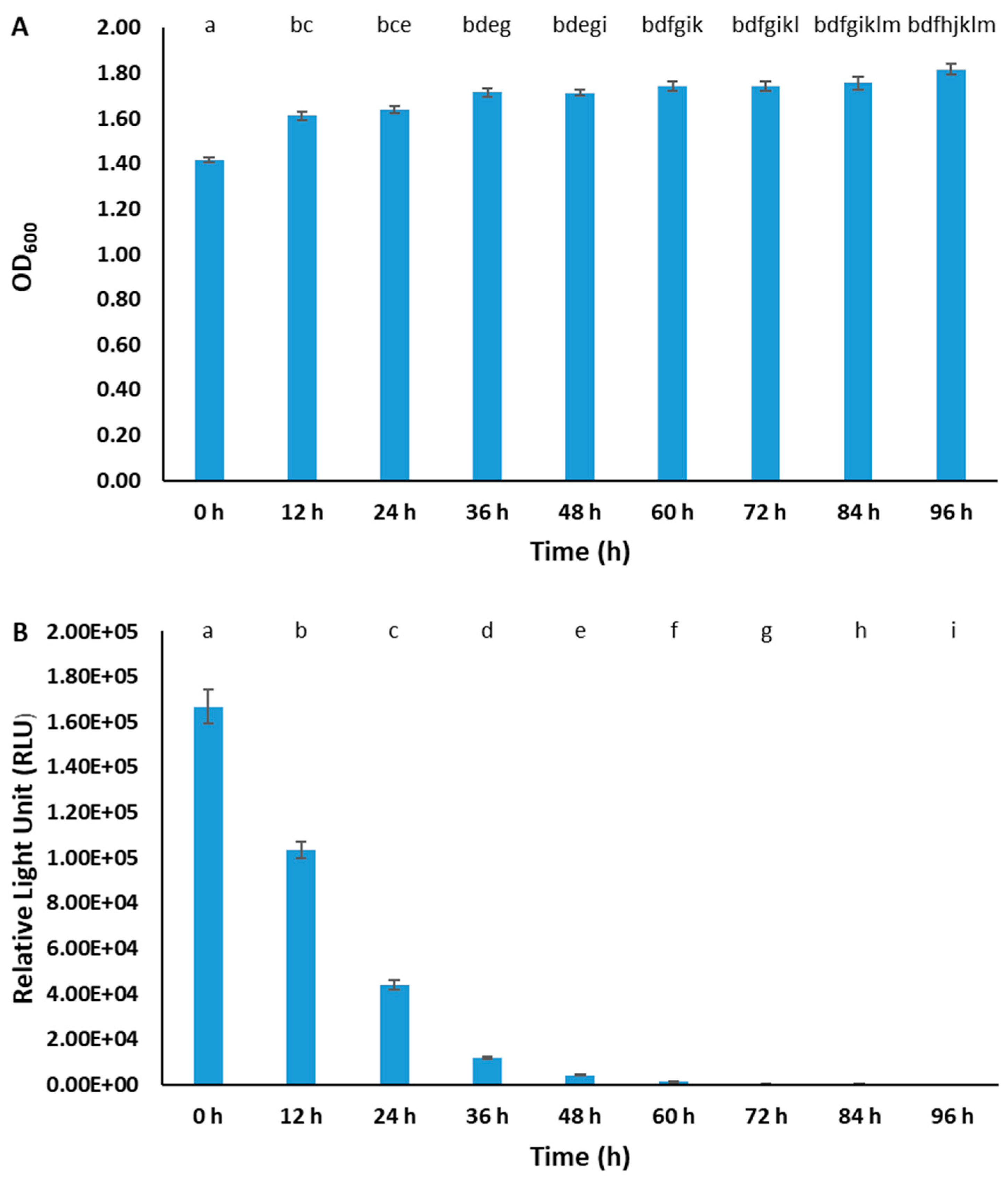
2.4. Plasmid Stability
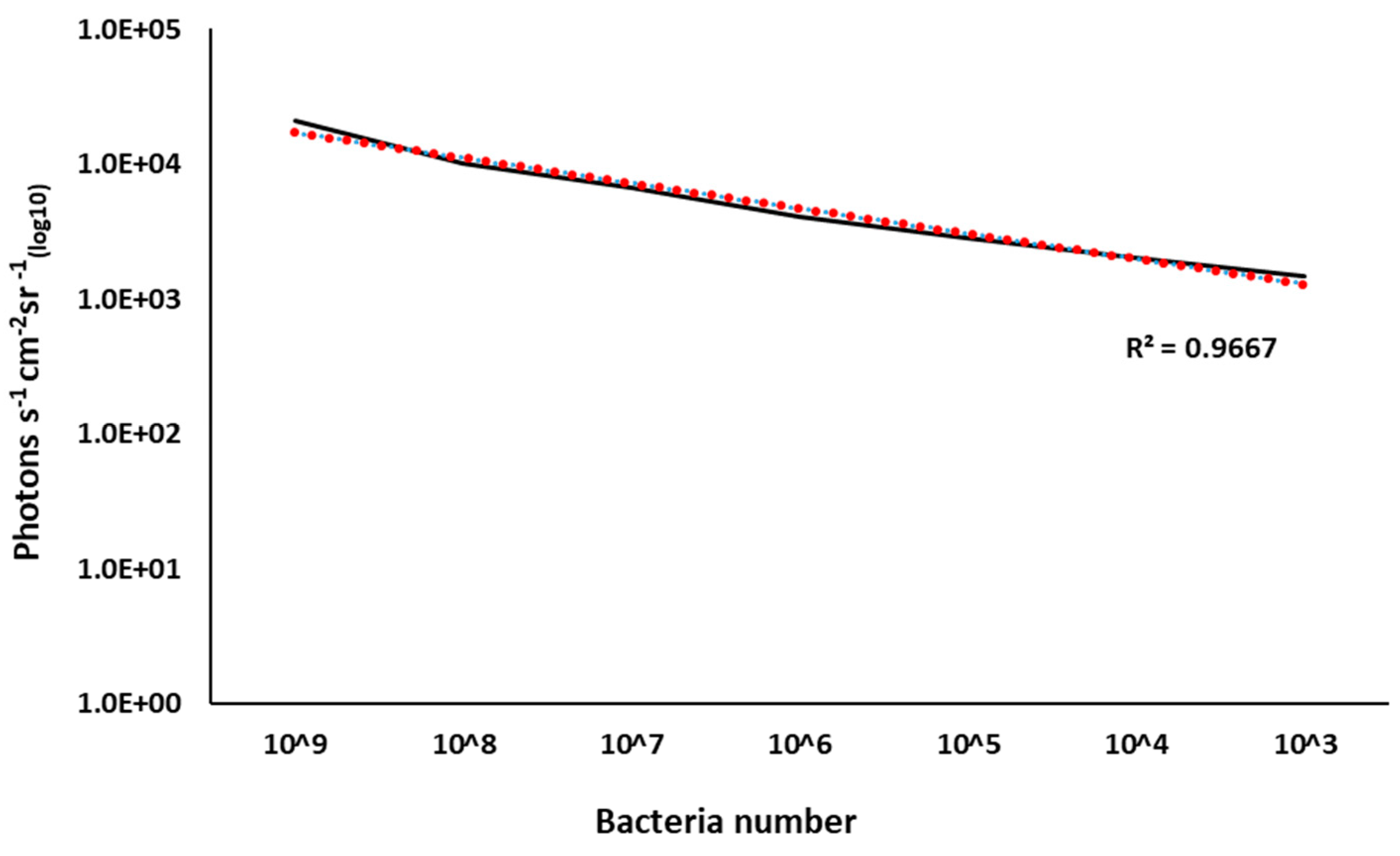
2.5. BvAh Number and Bioluminescence Relationship
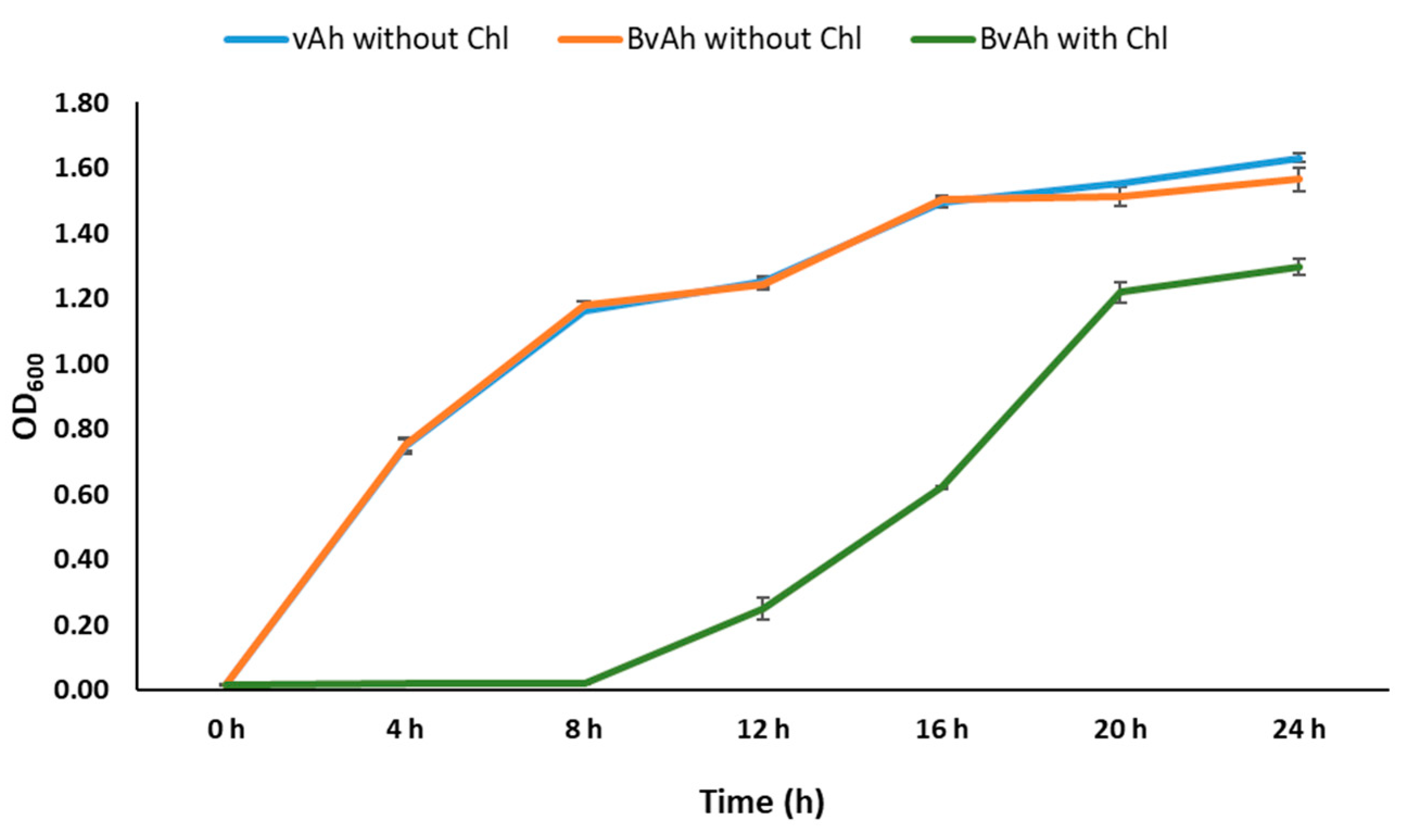
2.6. Growth of BvAh and vAh
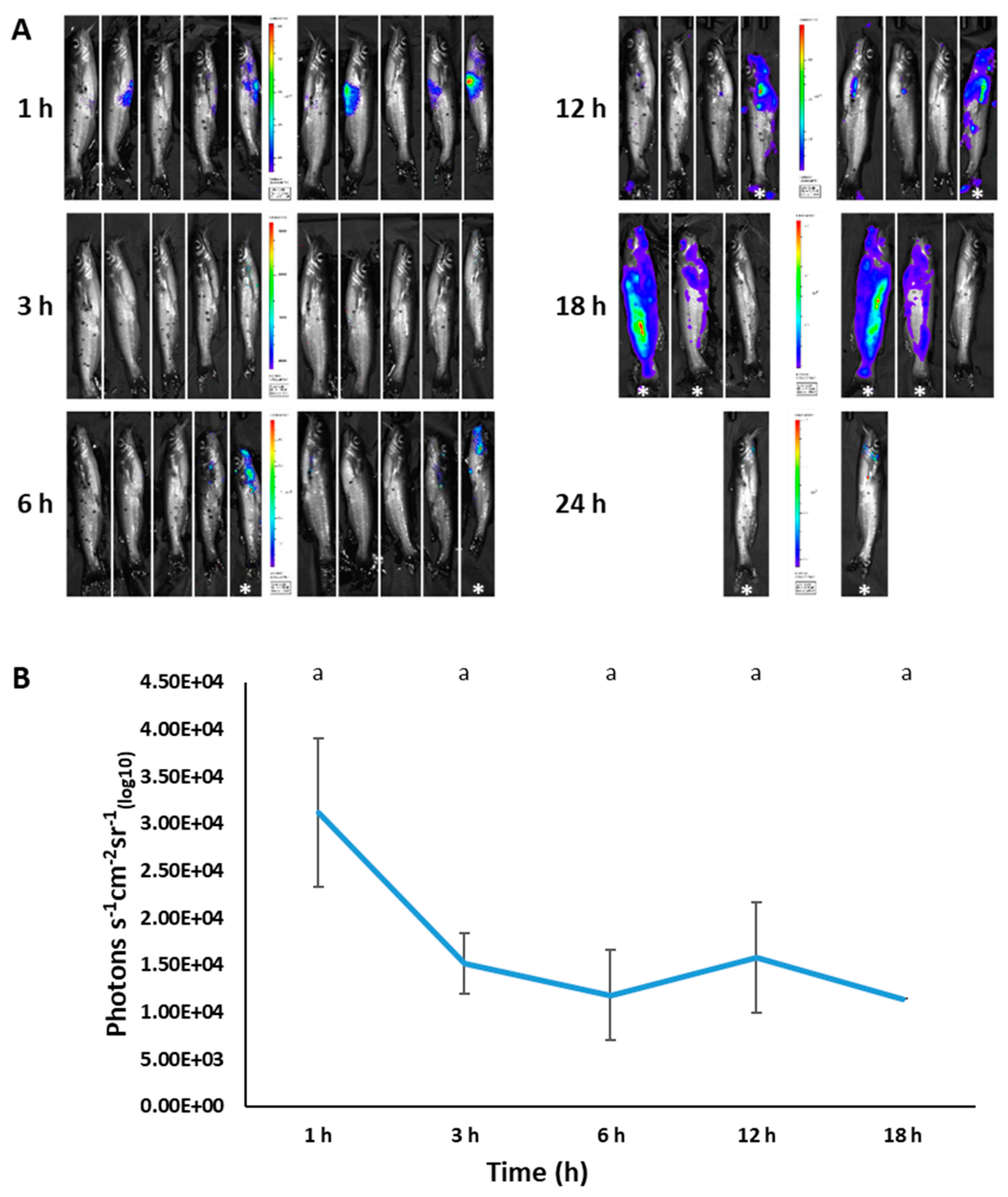
2.7. Imaging of BvAh in Live Catfish
2.8. Estimation of BvAh Quantities in Catfish Organs
2.9. Statistical Analysis
3. Results
3.1. Construction of pAKgfplux3
3.2. Determination of Optimal Chloramphenicol Concentration
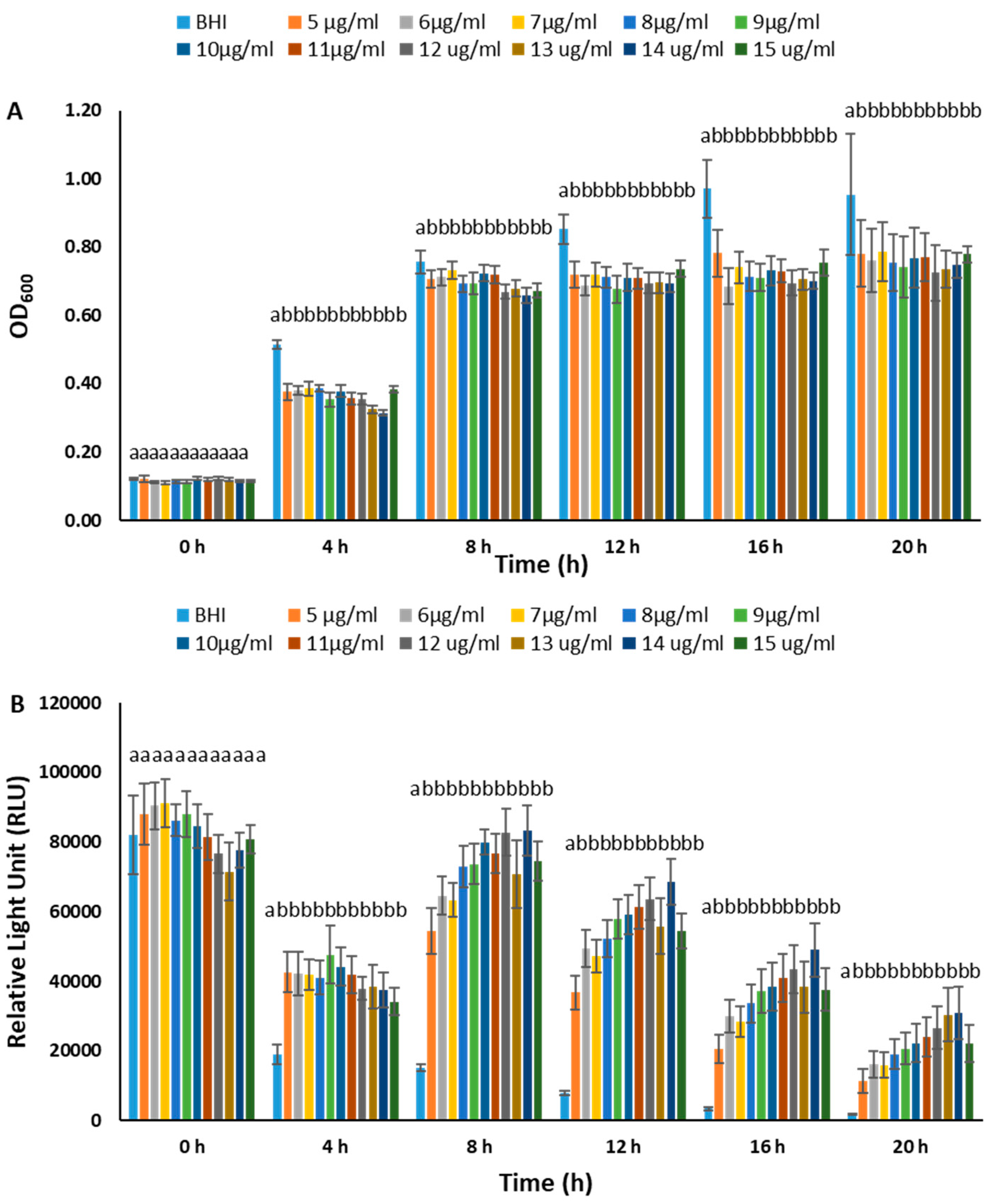
3.3. Stability of pAKgfplux3 in BvAh Strain
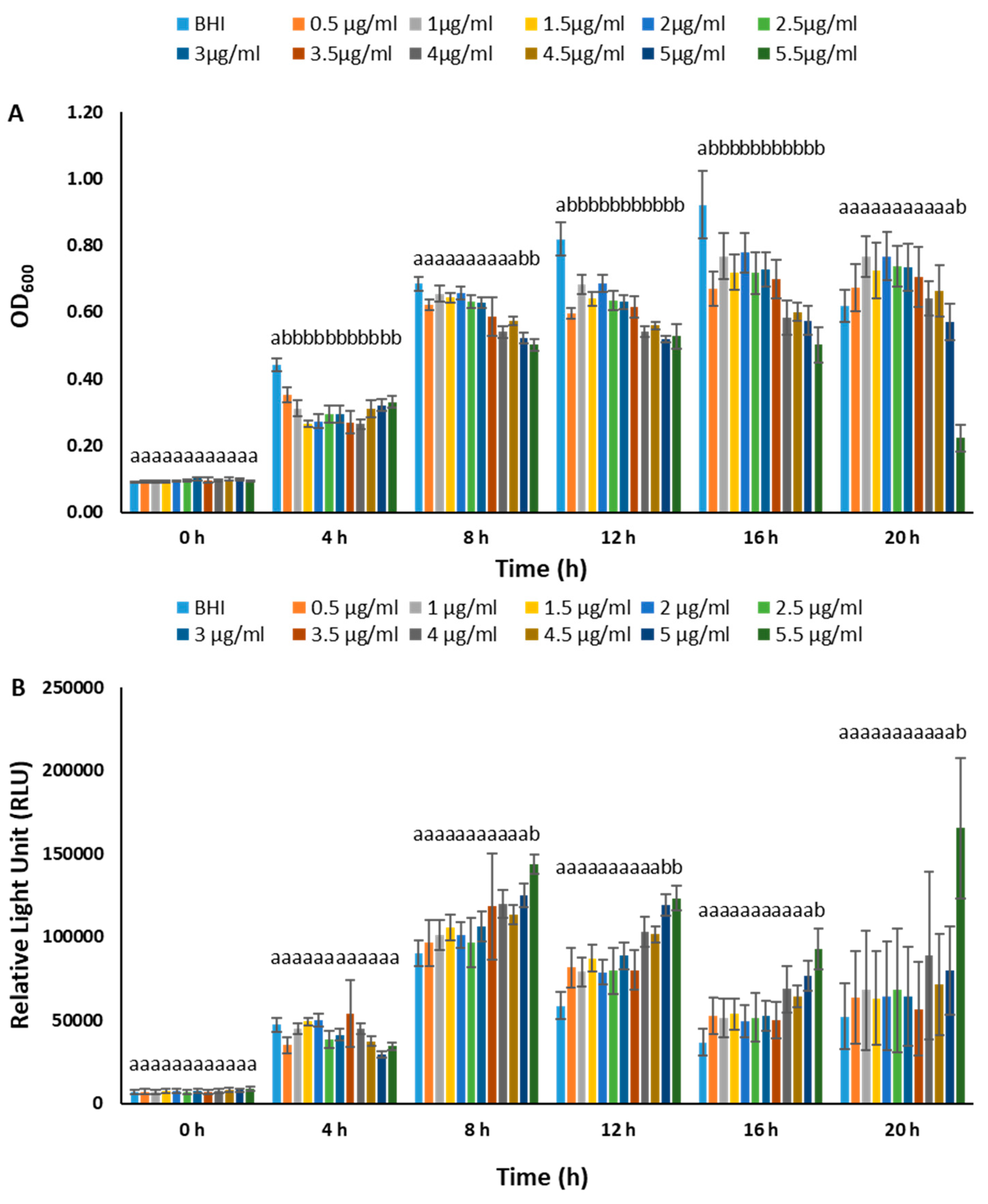
3.4. BvAh Number and Bioluminescence Relationship
3.5. Growth of BvAh and vAh
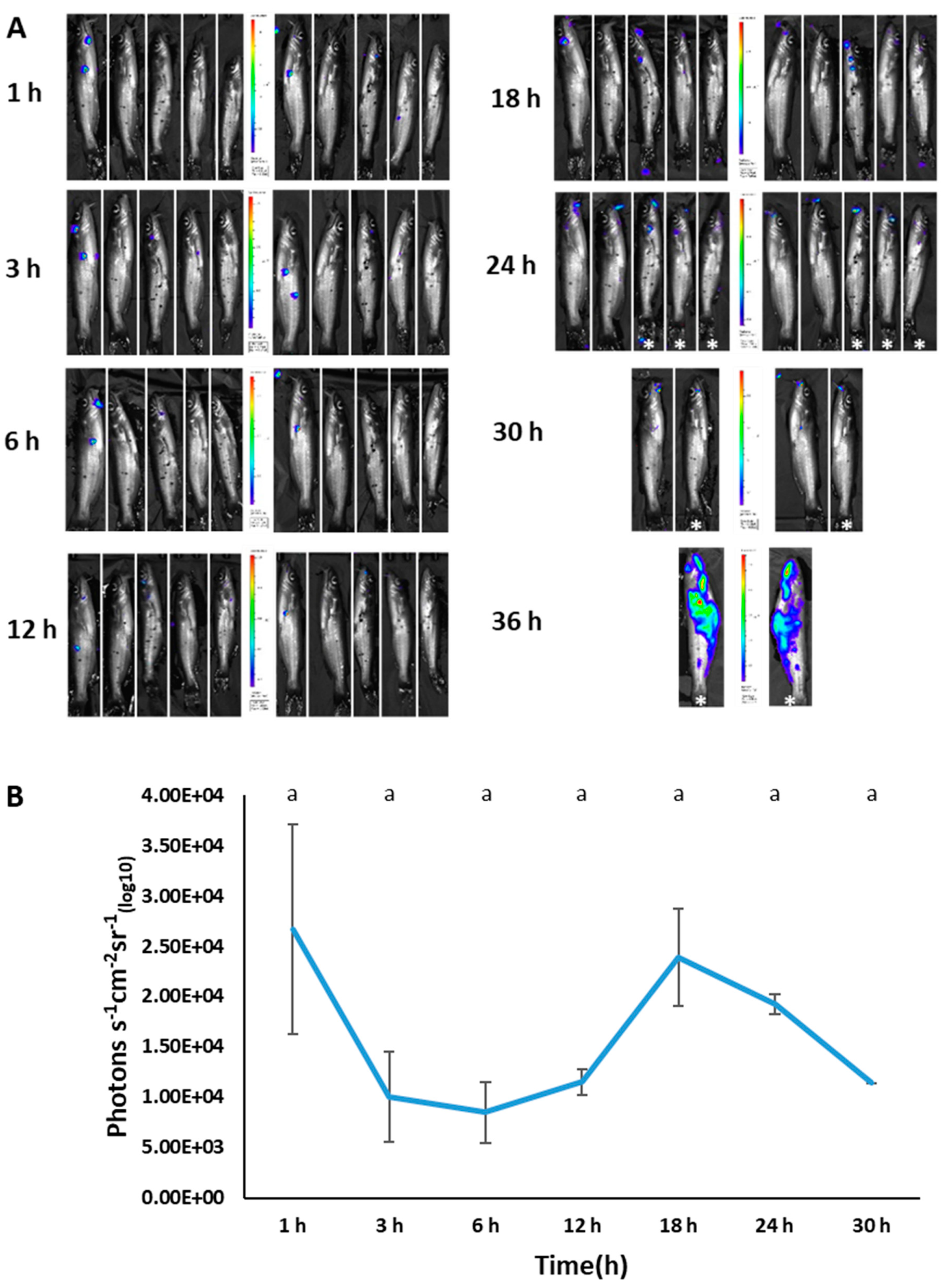
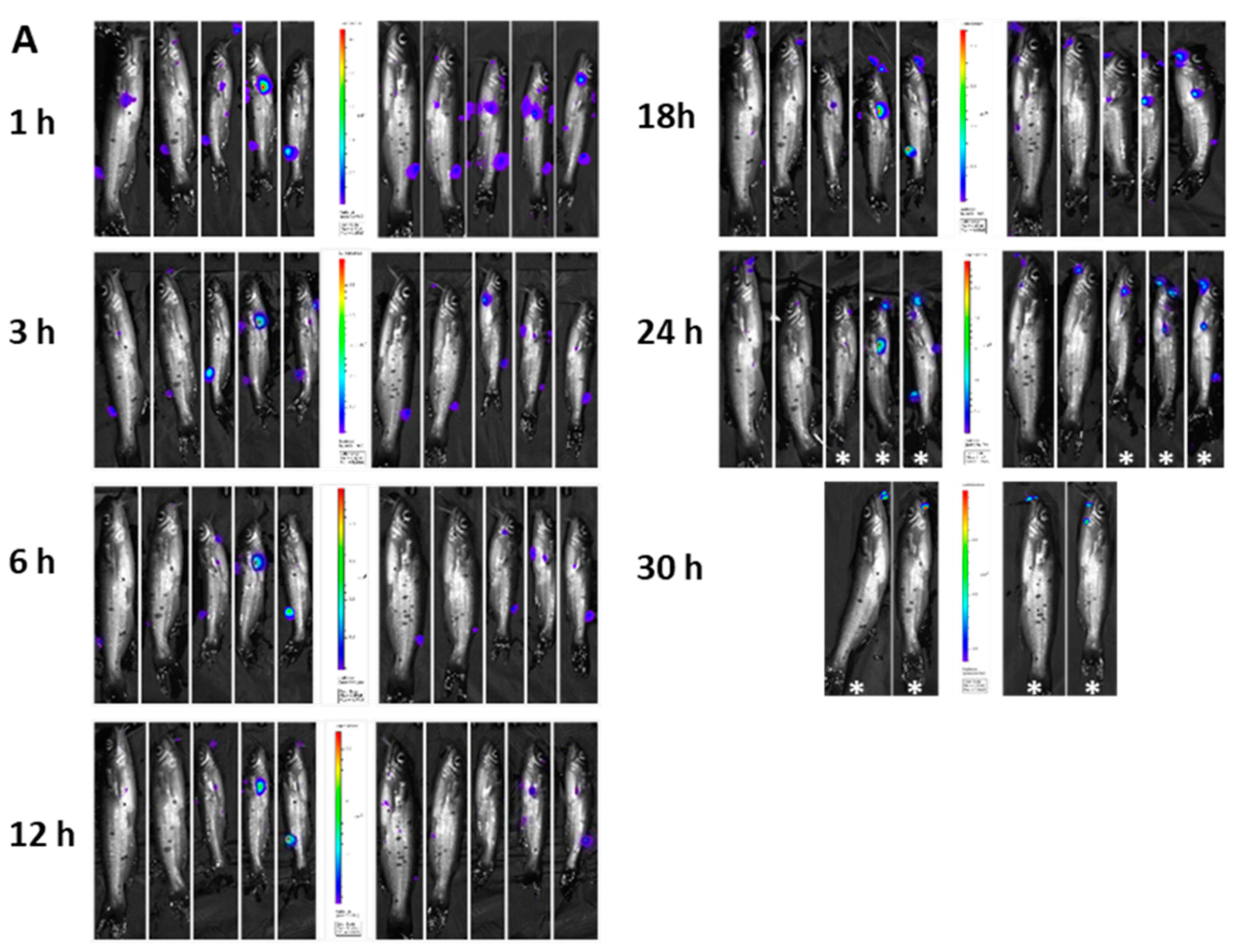
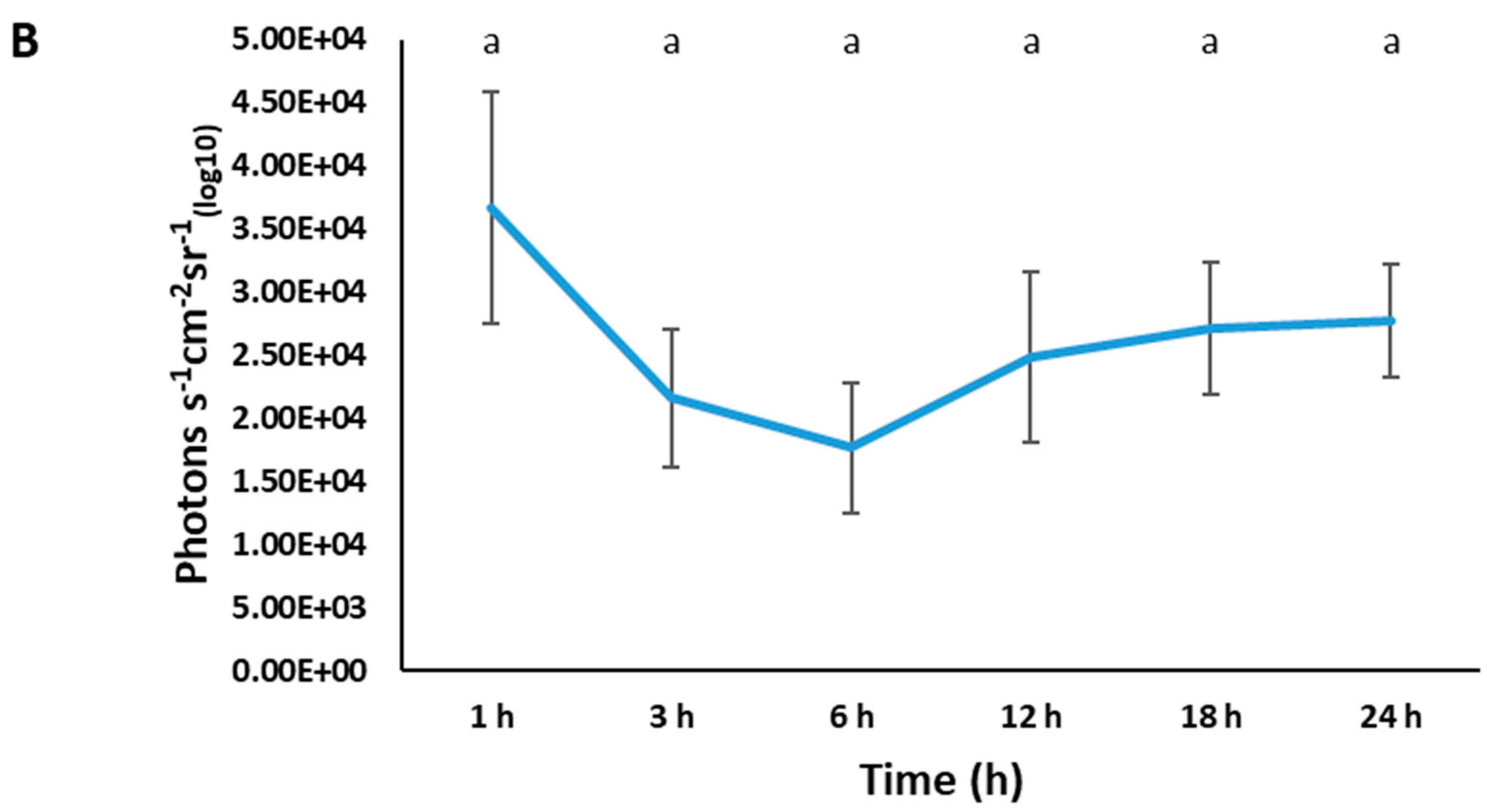
3.6. Imaging of BvAh in Live Catfish
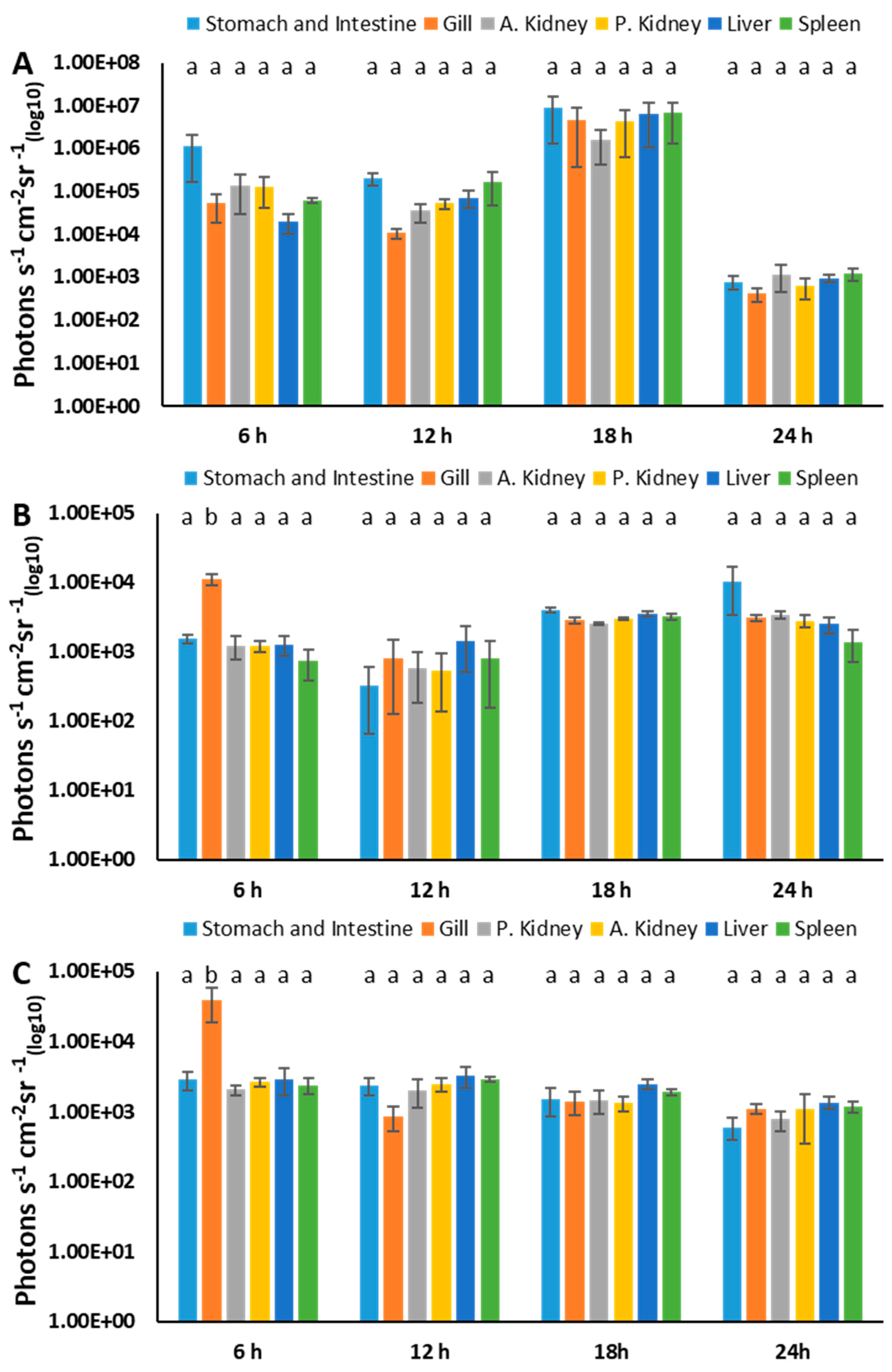
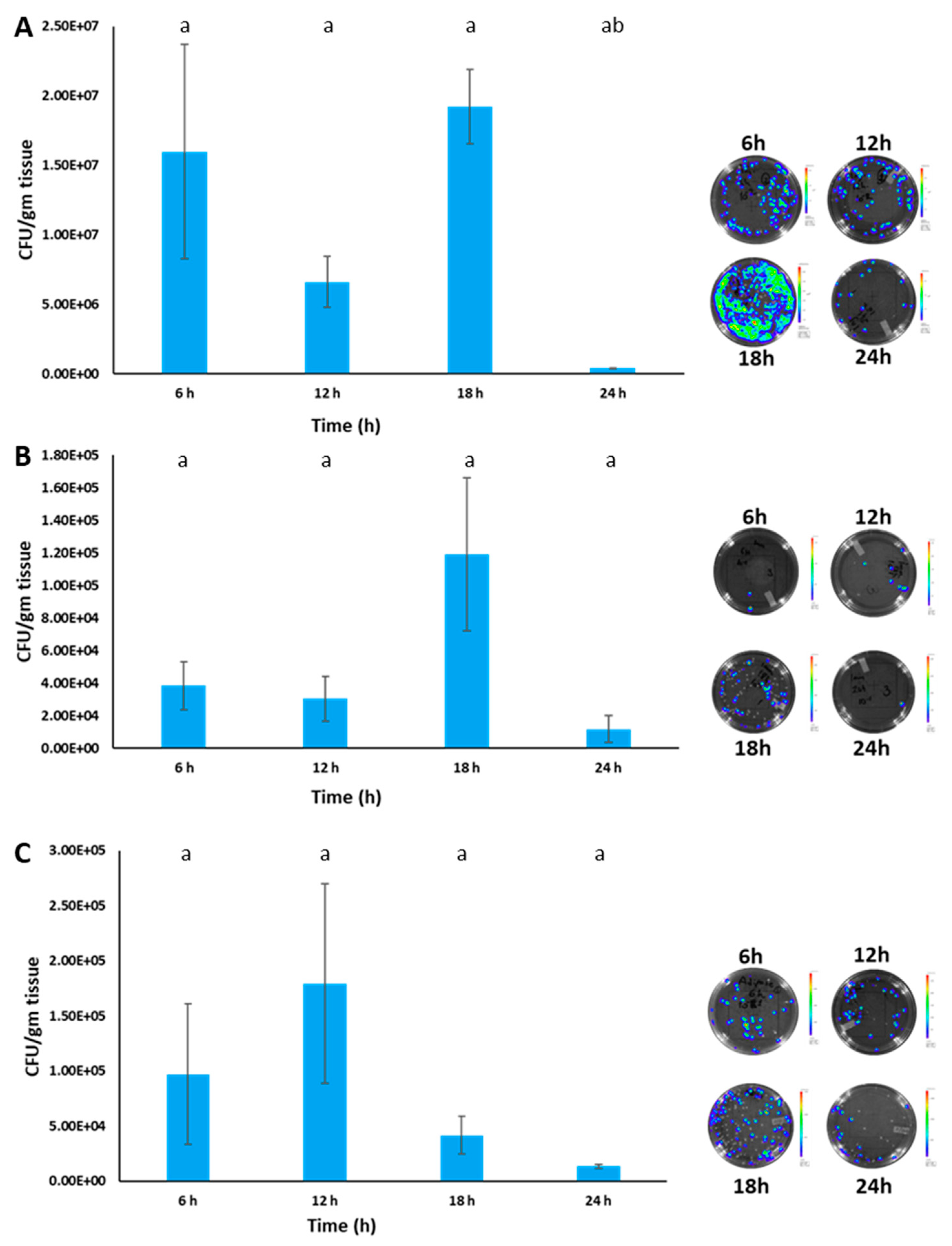
3.7. Estimation of BvAh Quantities in Catfish Organs
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Posadas, B.U.S. catfish production, farm-gate and wholesale prices, and imports. Miss. Mark. Newsl. 2020, 10, 1–9. [Google Scholar]
- Janda, J.M.; Abbott, S.L. The genus Aeromonas: Taxonomy, pathogenicity, and infection. Clin. Microbiol. Rev. 2010, 23, 35–73. [Google Scholar] [CrossRef] [PubMed]
- Camus, A.; Durborow, R.; Hemstreet, W.; Thune, R.; Hawke, J. Aeromonas Bacterial Infections-Motile Aeromonad Septicemia; Southern Regional Aquaculture Center: Stoneville, MS, USA, 1998. [Google Scholar]
- Plumb, J.A.; Grizzle, J.M.; Defigueiredo, J. Necrosis and bacterial infection in channel catfish (Ictalurus punctatus) following hypoxia. J. Wildl. Dis. 1976, 12, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Walters, G.R.; Plumb, J.A. Environmental stress and bacterial infection in channel catfish, Ictalurus punctatus Rafinesque. J. Fish Biol. 1980, 17, 177–185. [Google Scholar] [CrossRef]
- Hemstreet, W.B. An update on Aeromonas hydrophila from a fish health specialist for summer 2010. Catfish J. 2010, 24, 4. [Google Scholar]
- Bebak, J.A.; Hemstreet, W.B.; Garcia, J.C. Aeromonas hydrophila in 2010: Characteristics of Alabama outbreaks. Ala. Fish Farming Cent. Fish Farming News 2011, 8, 1–2. [Google Scholar]
- Pridgeon, J.; Klesius, P. Molecular identification and virulence of three Aeromonas hydrophila isolates cultured from infected channel catfish during a disease outbreak in west Alabama (USA) in 2009. Dis. Aquat. Org. 2011, 94, 249–253. [Google Scholar] [CrossRef]
- Pridgeon, J.W.; Klesius, P.H.; Mu, X.; Carter, D.; Fleming, K.; Xu, D.; Srivastava, K.; Reddy, G. Identification of unique DNA sequences present in highly virulent 2009 Alabama isolates of Aeromonas hydrophila. Veter- Microbiol. 2011, 152, 117–125. [Google Scholar] [CrossRef]
- Pridgeon, J.W.; Zhang, D.; Zhang, L. Complete Genome Sequence of the Highly Virulent Aeromonas hydrophila AL09-71 Isolated from Diseased Channel Catfish in West Alabama. Genome Announc. 2014, 2, e00450-14. [Google Scholar] [CrossRef]
- Tekedar, H.C.; Waldbieser, G.C.; Karsi, A.; Liles, M.R.; Griffin, M.J.; Vamenta, S.; Sonstegard, T.; Hossain, M.; Schroeder, S.G.; Khoo, L.; et al. Complete Genome Sequence of a Channel Catfish Epidemic Isolate, Aeromonas hydrophila Strain ML09-119. Genome Announc. 2013, 1, e00755-13. [Google Scholar] [CrossRef]
- Hossain, M.J.; Waldbieser, G.C.; Sun, D.; Capps, N.K.; Hemstreet, W.B.; Carlisle, K.; Griffin, M.J.; Khoo, L.; Goodwin, A.E.; Sonstegard, T.S.; et al. Implication of Lateral Genetic Transfer in the Emergence of Aeromonas hydrophila Isolates of Epidemic Outbreaks in Channel Catfish. PLoS ONE 2013, 8, e80943. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.J.; Sun, D.; McGarey, D.J.; Wrenn, S.; Alexander, L.M.; Martino, M.E.; Xing, Y.; Terhune, J.S.; Liles, M.R. An Asian Origin of Virulent Aeromonas hydrophila Responsible for Disease Epidemics in United States-Farmed Catfish. mBio 2014, 5, e00848-14. [Google Scholar] [CrossRef] [PubMed]
- Tekedar, H.C.; Abdelhamed, H.; Kumru, S.; Blom, J.; Karsi, A.; Lawrence, M.L. Comparative Genomics of Aeromonas hydrophila Secretion Systems and Mutational Analysis of hcp1 and vgrG1 Genes from T6SS. Front. Microbiol. 2019, 9, 3216. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen-Ivey, C.R.; Hossain, M.J.; Odom, S.E.; Terhune, J.S.; Hemstreet, W.G.; Shoemaker, C.A.; Zhang, D.; Xu, D.-H.; Griffin, M.J.; Liu, Y.-J.; et al. Classification of a Hypervirulent Aeromonas hydrophila Pathotype Responsible for Epidemic Outbreaks in Warm-Water Fishes. Front. Microbiol. 2016, 7, 1615. [Google Scholar] [CrossRef] [PubMed]
- Baumgartner, W.A.; Griffin, M.J.; Tekedar, H.C.; Lawrence, M.L.; Rasmussen-Ivey, C.; Liles, M.R. Experience with mortalities of cultured Catfish Ictalurus punctatus (Rafinesque 1818) and I. punctatus X I. furcatus (valenciennes 1840) caused by highly virulent strains of Aeromonas hydrophila. Asian Fish. Sci. 2018, 31, 59–75. [Google Scholar] [CrossRef]
- Pridgeon, J.W.; Klesius, P.H.; Song, L.; Zhang, D.; Kojima, K.; Mobley, J.A. Identification, virulence, and mass spectrometry of toxic ECP fractions of West Alabama isolates of Aeromonas hydrophila obtained from a 2010 disease outbreak. Veter- Microbiol. 2013, 164, 336–343. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Pridgeon, J.W.; Klesius, P.H. Expression and activity of recombinant proaerolysin derived from Aeromonas hydrophila cultured from diseased channel catfish. Veter- Microbiol. 2013, 165, 478–482. [Google Scholar] [CrossRef]
- Abdelhamed, H.; Nho, S.W.; Turaga, G.; Banes, M.M.; Karsi, A.; Lawrence, M.L. Protective efficacy of four recombinant fimbrial proteins of virulent Aeromonas hydrophila strain ML09-119 in channel catfish. Veter- Microbiol. 2016, 197, 8–14. [Google Scholar] [CrossRef]
- Abdelhamed, H.; Ibrahim, I.; Nho, S.W.; Banes, M.M.; Wills, R.W.; Karsi, A.; Lawrence, M.L. Evaluation of three recombinant outer membrane proteins, OmpA1, Tdr, and TbpA, as potential vaccine antigens against virulent Aeromonas hydrophila infection in channel catfish (Ictalurus punctatus). Fish Shellfish. Immunol. 2017, 66, 480–486. [Google Scholar] [CrossRef]
- Yeh, H.-Y.; Klesius, P.H. Over-expression, purification and immune responses to Aeromonas hydrophila AL09-73 flagellar proteins. Fish Shellfish. Immunol. 2011, 31, 1278–1283. [Google Scholar] [CrossRef]
- Rasmussen-Ivey, C.R.; Figueras, M.J.; McGarey, D.; Liles, M.R. Virulence Factors of Aeromonas hydrophila: In the Wake of Reclassification. Front. Microbiol. 2016, 7, 1337. [Google Scholar] [CrossRef]
- Thurlow, C.M.; Hossain, M.J.; Sun, D.; Barger, P.; Foshee, L.; Beck, B.; Newton, J.C.; Terhune, J.S.; Saper, M.A.; Liles, M.R. The gfc operon is involved in the formation of the O antigen capsule in Aeromonas hydrophila and contributes to virulence in channel catfish. Aquaculture 2019, 512, 73433. [Google Scholar] [CrossRef]
- Barger, P.C.; Liles, M.R.; Beck, B.H.; Newton, J.C. Differential production and secretion of potentially toxigenic extracellular proteins from hypervirulent Aeromonas hydrophila under biofilm and planktonic culture. BMC Microbiol. 2021, 21, 132. [Google Scholar] [CrossRef]
- Barger, P.C.; Liles, M.R.; Newton, J.C. Type II Secretion Is Essential for Virulence of the Emerging Fish Pathogen, Hypervirulent Aeromonas hydrophila. Front. Veter- Sci. 2020, 7. [Google Scholar] [CrossRef] [PubMed]
- Jubirt, M.M.; Hanson, L.A.; Hanson-Dorr, K.C.; Ford, L.; Lemmons, S.; Fioranelli, P.; Cunningham, F.L. Potential for great egrets (Ardea alba) to transmit a virulent strain of Aeromonas hydrophila among channel catfish (Ictalurus punctatus) culture ponds. J. Wildl. Dis. 2015, 51, 634–639. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, F.L.; Jubirt, M.M.; Hanson-Dorr, K.C.; Ford, L.; Fioranelli, P.; Hanson, L.A. Potential of Double-crested Cormorants (Phalacrocorax auritus), American White Pelicans (Pelecanus erythrorhynchos), and Wood Storks (Mycteria americana) to Transmit a Hypervirulent Strain of Aeromonas hydrophila between Channel Catfish Culture Ponds. J. Wildl. Dis. 2018, 54, 548–552. [Google Scholar] [CrossRef]
- Zhang, X.; Cai, W.; Tao, Z.; Arias, C.R. Survival of Fish-Pathogenic Strains of Aeromonas hydrophila under Starvation. J. Aquat. Anim. Health 2014, 26, 190–193. [Google Scholar] [CrossRef]
- Baumgartner, W.A.; Ford, L.; Hanson, L. Lesions caused by virulent Aeromonas hydrophila in farmed catfish (Ictalurus punctatus and I. punctatus × I. furcatus) in Mississippi. J. Veter- Diagn. Investig. 2017, 29, 747–751. [Google Scholar] [CrossRef]
- Li, C.; Wang, R.; Su, B.; Luo, Y.; Terhune, J.; Beck, B.; Peatman, E. Evasion of mucosal defenses during Aeromonas hydrophila infection of channel catfish (Ictalurus punctatus) skin. Dev. Comp. Immunol. 2013, 39, 447–455. [Google Scholar] [CrossRef]
- Zhang, D.; Xu, D.-H.; Shoemaker, C. Experimental induction of motile Aeromonas septicemia in channel catfish (Ictalurus punctatus) by waterborne challenge with virulent Aeromonas hydrophila. Aquac. Rep. 2016, 3, 18–23. [Google Scholar] [CrossRef]
- Zhang, D.; Moreira, G.S.A.; Shoemaker, C.; Newton, J.C.; Xu, D.-H. Detection and quantification of virulent Aeromonas hydrophila in channel catfish tissues following waterborne challenge. FEMS Microbiol. Lett. 2016, 363. [Google Scholar] [CrossRef]
- Abdelhamed, H.; Ibrahim, I.; Baumgartner, W.; Lawrence, M.L.; Karsi, A. Characterization of Histopathological and Ultrastructural Changes in Channel Catfish Experimentally Infected with Virulent Aeromonas hydrophila. Front. Microbiol. 2017, 8, 1519. [Google Scholar] [CrossRef] [PubMed]
- Peatman, E.; Mohammed, H.; Kirby, A.; Shoemaker, C.A.; Yildirim-Aksoy, M.; Beck, B.H. Mechanisms of pathogen virulence and host susceptibility in virulent Aeromonas hydrophila infections of channel catfish (Ictalurus punctatus). Aquaculture 2018, 482, 1–8. [Google Scholar] [CrossRef]
- Meighen, E.A. Molecular biology of bacterial bioluminescence. Microbiol. Rev. 1991, 55, 123–142. [Google Scholar] [CrossRef]
- Contag, C.H.; Contag, P.R.; Mullins, J.I.; Spilman, S.D.; Stevenson, D.K.; Benaron, D.A. Photonic detection of bacterial pathogens in living hosts. Mol. Microbiol. 1995, 18, 593–603. [Google Scholar] [CrossRef] [PubMed]
- Contag, C.H.; Spilman, S.D.; Contag, P.R.; Oshiro, M.; Eames, B.; Dennery, P.; Stevenson, D.K.; Benaron, D.A. Visualizing Gene Expression in Living Mammals Using a Bioluminescent Reporter. Photochem. Photobiol. 1997, 66, 523–531. [Google Scholar] [CrossRef]
- Contag, P.R.; Olomu, I.N.; Stevenson, D.K.; Contag, C. Bioluminescent indicators in living mammals. Nat. Med. 1998, 4, 245–247. [Google Scholar] [CrossRef]
- Karsi, A.; Lawrence, M.L. Broad host range fluorescence and bioluminescence expression vectors for Gram-negative bacteria. Plasmid 2007, 57, 286–295. [Google Scholar] [CrossRef]
- Karsi, A.; Menanteau-Ledouble, S.; Lawrence, M.L. Development of bioluminescent Edwardsiella ictaluri for noninvasive disease monitoring. FEMS Microbiol. Lett. 2006, 260, 216–223. [Google Scholar] [CrossRef]
- Dahal, N.; Abdelhamed, H.; Lu, J.; Karsi, A.; Lawrence, M.L. Tricarboxylic Acid Cycle and One-Carbon Metabolism Pathways Are Important in Edwardsiella ictaluri Virulence. PLoS ONE 2013, 8, e65973. [Google Scholar] [CrossRef]
- Dahal, N.; Abdelhamed, H.; Lu, J.; Karsi, A.; Lawrence, M. Effect of multiple mutations in tricarboxylic acid cycle and one-carbon metabolism pathways on Edwardsiella ictaluri pathogenesis. Veter- Microbiol. 2014, 169, 107–112. [Google Scholar] [CrossRef]
- Menanteau-Ledouble, S.; Karsi, A.; Lawrence, M.L. Importance of skin abrasion as a primary site of adhesion for Edwardsiella ictaluri and impact on invasion and systematic infection in channel catfish Ictalurus punctatus. Veter- Microbiol. 2011, 148, 425–430. [Google Scholar] [CrossRef] [PubMed]
- Kalindamar, S.; Abdelhamed, H.; Kordon, A.O.; Pinchuk, L.M.; Karsi, A. Hemolysin Co-regulated Family Proteins Hcp1 and Hcp2 Contribute to Edwardsiella ictaluri Pathogenesis. Front. Veter- Sci. 2021, 8, 681609. [Google Scholar] [CrossRef] [PubMed]
- Miller, V.; Mekalanos, J.J. A novel suicide vector and its use in construction of insertion mutations: Osmoregulation of outer membrane proteins and virulence determinants in Vibrio cholerae requires toxR. J. Bacteriol. 1988, 170, 2575–2583. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.J.; Thurlow, C.M.; Sun, D.; Nasrin, S.; Liles, M.R. Genome modifications and cloning using a conjugally transferable recombineering system. Biotechnol. Rep. 2015, 8, 24–35. [Google Scholar] [CrossRef]
- Karsi, A.; Howe, K.; Kirkpatrick, T.B.; Wills, R.; Bailey, R.H.; Lawrence, M.L. Development of bioluminescent Salmonella strains for use in food safety. BMC Microbiol. 2008, 8, 10. [Google Scholar] [CrossRef]
- Bartkova, S.; Kokotovic, B.; Dalsgaard, I. Infection routes of Aeromonas salmonicida in rainbow trout monitored in vivo by real-time bioluminescence imaging. J. Fish Dis. 2017, 40, 73–82. [Google Scholar] [CrossRef]
- Summers, D. The kinetics of plasmid loss. Trends Biotechnol. 1991, 9, 273–278. [Google Scholar] [CrossRef]
- Vander Haegen, G.; Blankenship, H.; Hoffmann, A.; Thompson, D. The effects of adipose fin clipping and coded wire tagging on the survival and growth of spring Chinook salmon. N. Am. J. Fish. Manag. 2005, 25, 1161–1170. [Google Scholar] [CrossRef]
- Temple, N.F.; Reimchen, T.E. Adipose fin condition and flow regime in catfish. Can. J. Zool. 2008, 86, 1079–1082. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains and Plasmids | Relevant Characteristics | References |
|---|---|---|
| Aeromonas hydrophila | ||
| ML09-119 | Wild type; Ampr, Colr; Chls | [12] |
| Escherichia coli | ||
| DH5α | F−; Φ80lacZΔM15; Δ(lacZYA-argF) U169; recA1; endA1; hsdR17(rk−, mk+); phoA; supE44; thi-1; gyrA96; relA1; λ− | ThermoFisher |
| SM10λpir | thiL; thrL; leuB6; tonA21; lacY1; supE44; recA::RP4-2-Tc:MuλpirR6K; Kmr | [45] |
| Plasmids | ||
| pAKgfplux1 | pBBR1MCS4; gfpmut3a; luxCDABE | [39] |
| pMJH46 | pKD46; oriT; traJ; traK; cat | [46] |
| pAKgfplux3 | pAKgfplux1; cat | This Study |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ozdemir, E.; Abdelhamed, H.; Ozdemir, O.; Lawrence, M.; Karsi, A. Development of Bioluminescent Virulent Aeromonas hydrophila for Understanding Pathogenicity. Pathogens 2023, 12, 670. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens12050670
Ozdemir E, Abdelhamed H, Ozdemir O, Lawrence M, Karsi A. Development of Bioluminescent Virulent Aeromonas hydrophila for Understanding Pathogenicity. Pathogens. 2023; 12(5):670. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens12050670
Chicago/Turabian StyleOzdemir, Eda, Hossam Abdelhamed, Ozan Ozdemir, Mark Lawrence, and Attila Karsi. 2023. "Development of Bioluminescent Virulent Aeromonas hydrophila for Understanding Pathogenicity" Pathogens 12, no. 5: 670. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens12050670