
Evaluation of the Effect of Plectranthus amboinicus L. Leaf Extracts on the Bacterial Antioxidant System and Cell Membrane Integrity of Pseudomonas aeruginosa PA01 and Staphylococcus aureus NCTC8325

Abstract
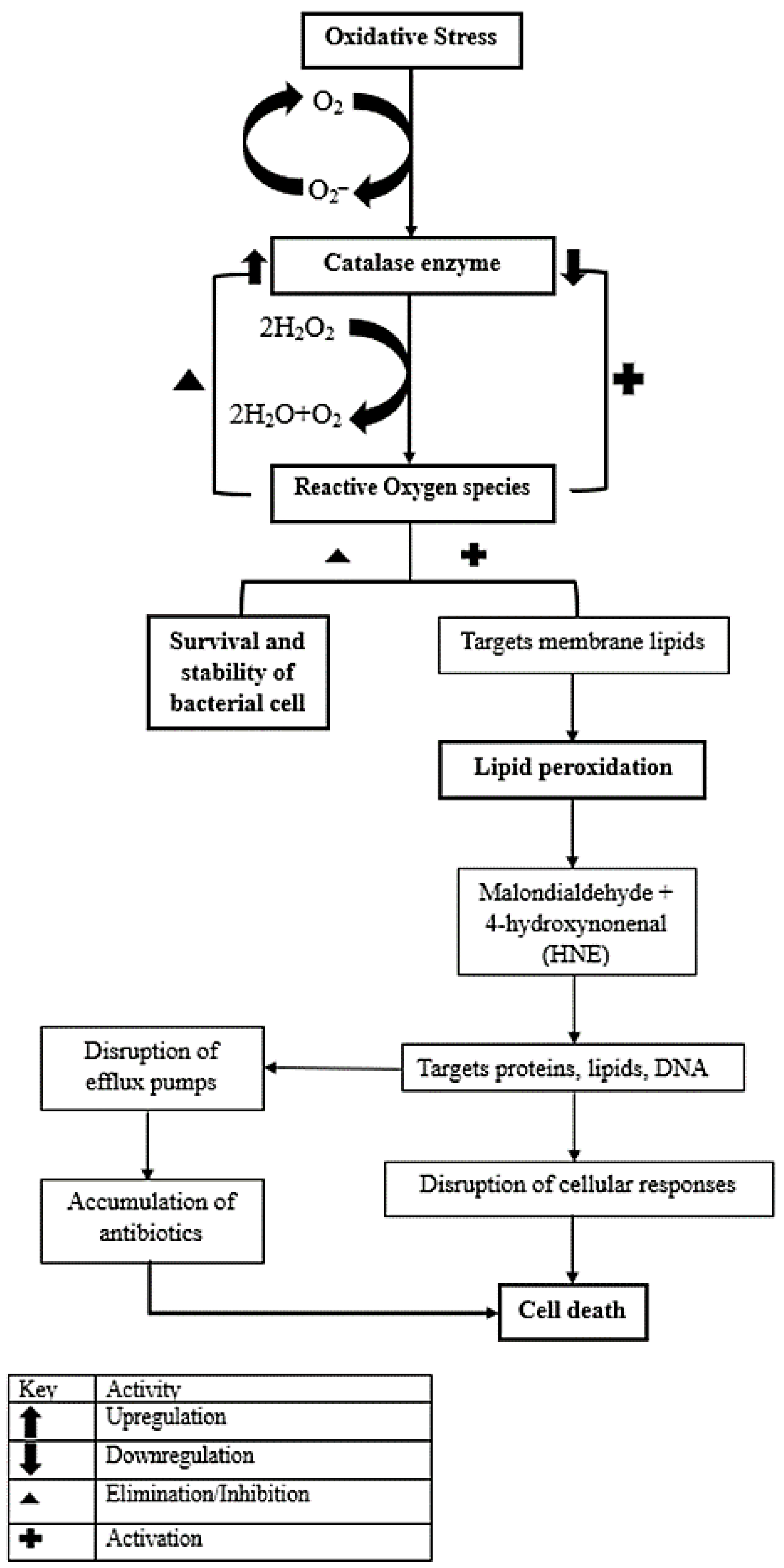
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Extract Preparation
2.3. Effect of IB Extracts on Catalase Activity in S. aureus NCTC8325 and P. aeruginosa PA01
Calculations
2.4. Lipid Peroxidation Assay
2.5. Measurement of Reactive Oxygen Species Concentration with 2′,7′-Dichlorodihydrofluorescein Diacetate (H2DCFDA)
2.6. Evaluation of Cell Membrane Integrity as a Potential Antimicrobial Target
2.6.1. Cytoplasmic Membrane Permeability Assay
2.6.2. Rhodamine 6-G Uptake Assay
2.7. Statistical Analysis
3. Results
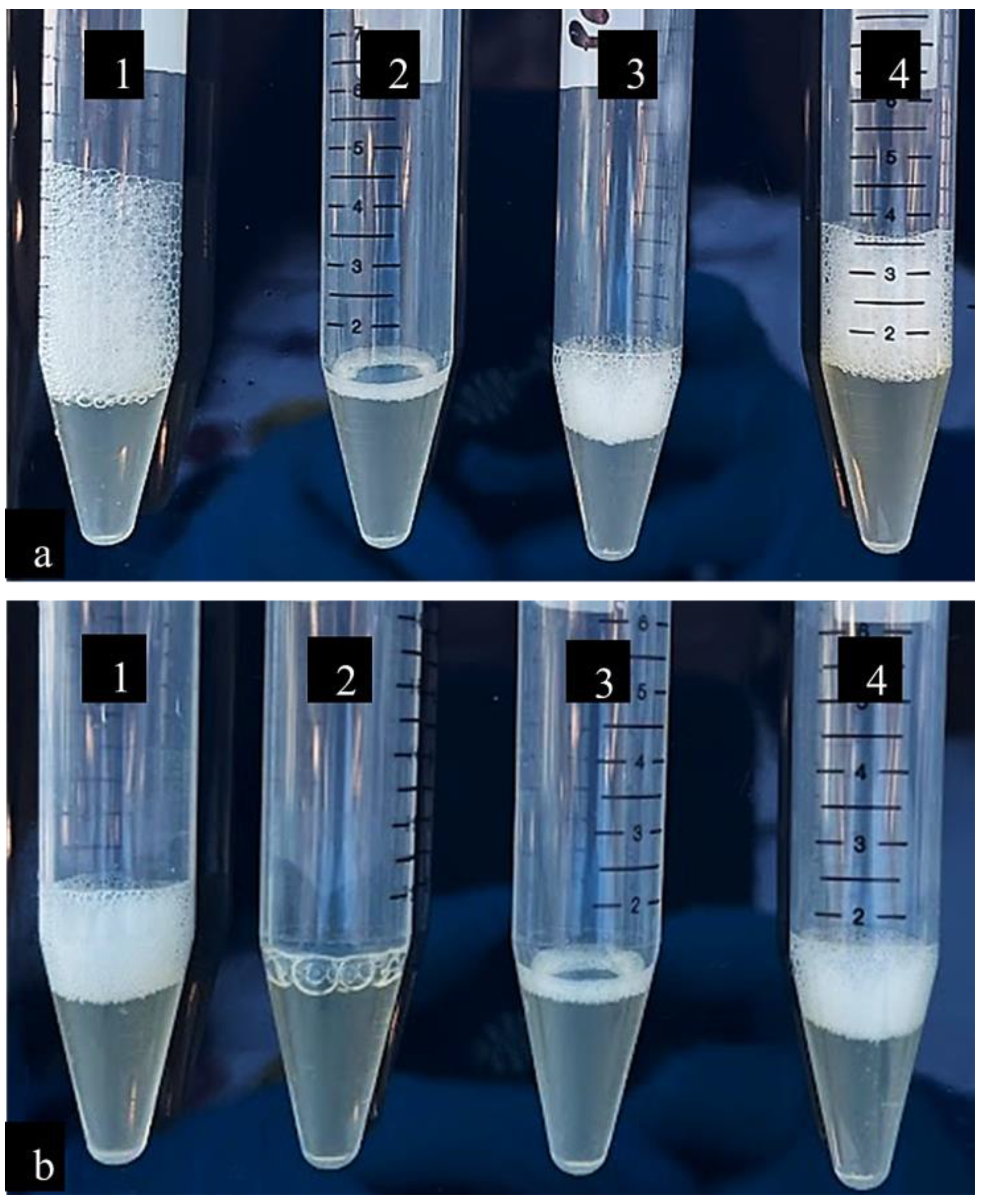
3.1. Catalase Activity
3.2. Lipid Peroxidation
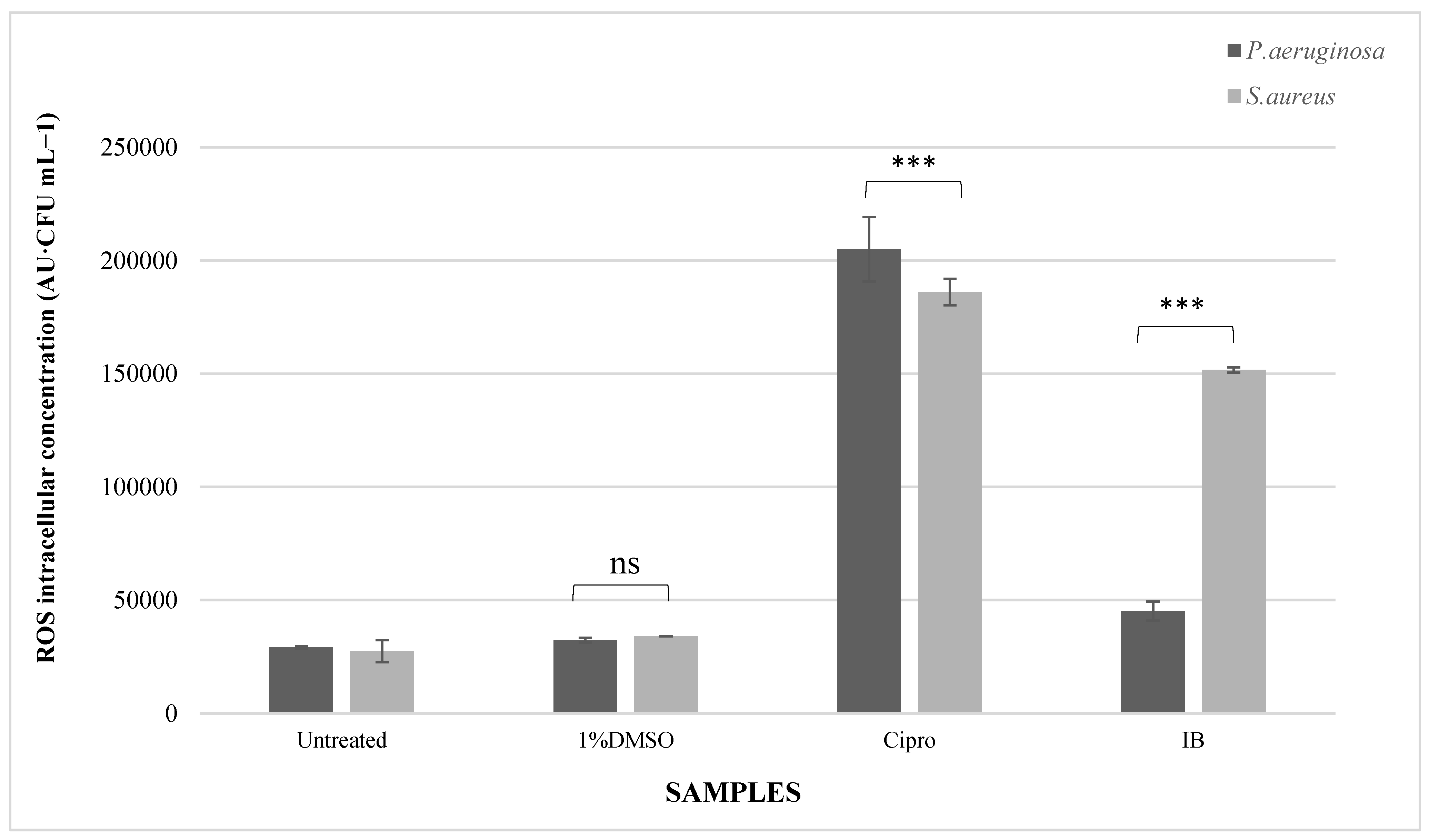
3.3. Reactive Oxygen Species
3.4. Evaluation of Cell Membrane Integrity as a Potential Antimicrobial Target
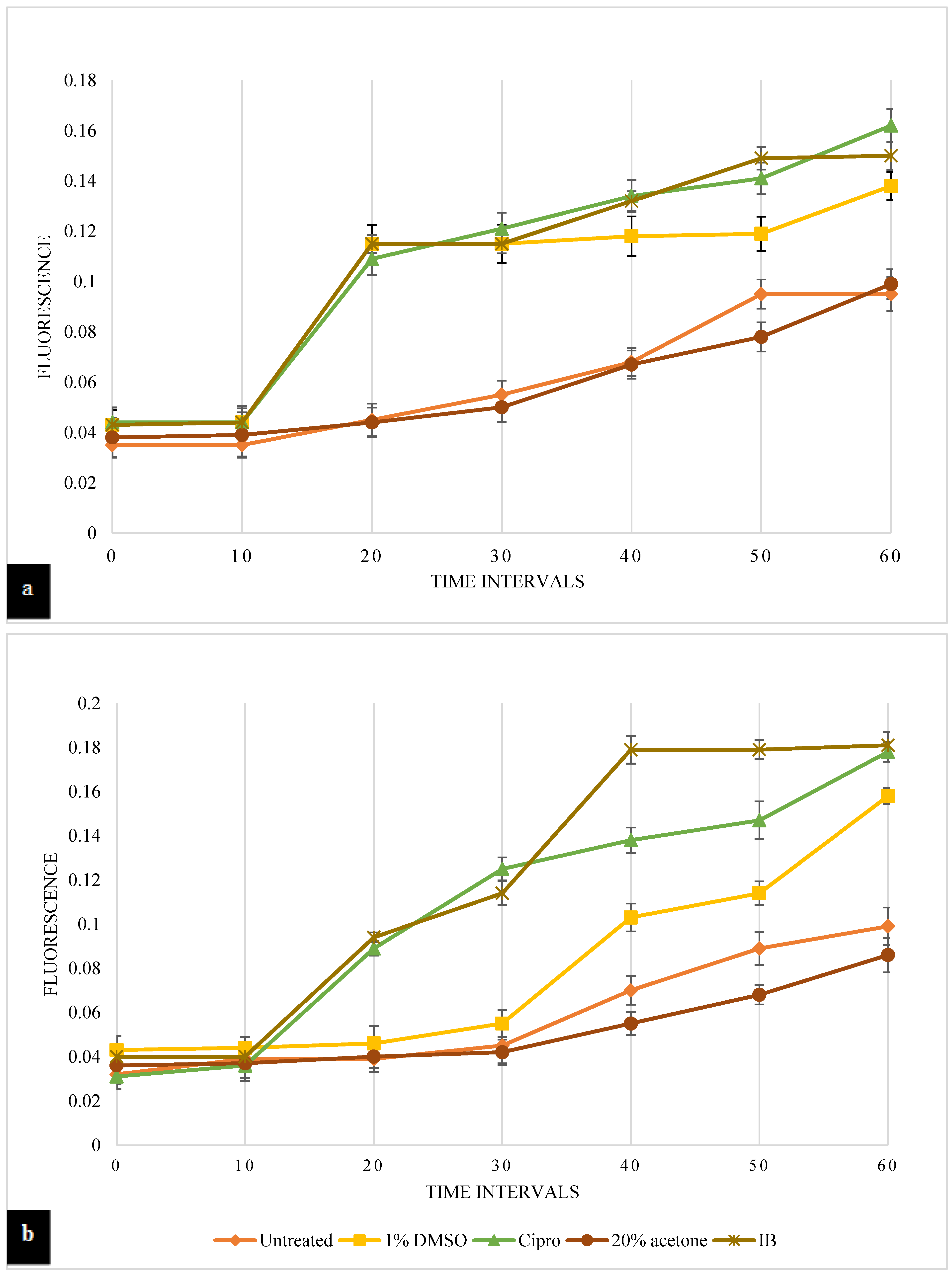
3.4.1. Cytoplasmic Membrane Permeability Assay
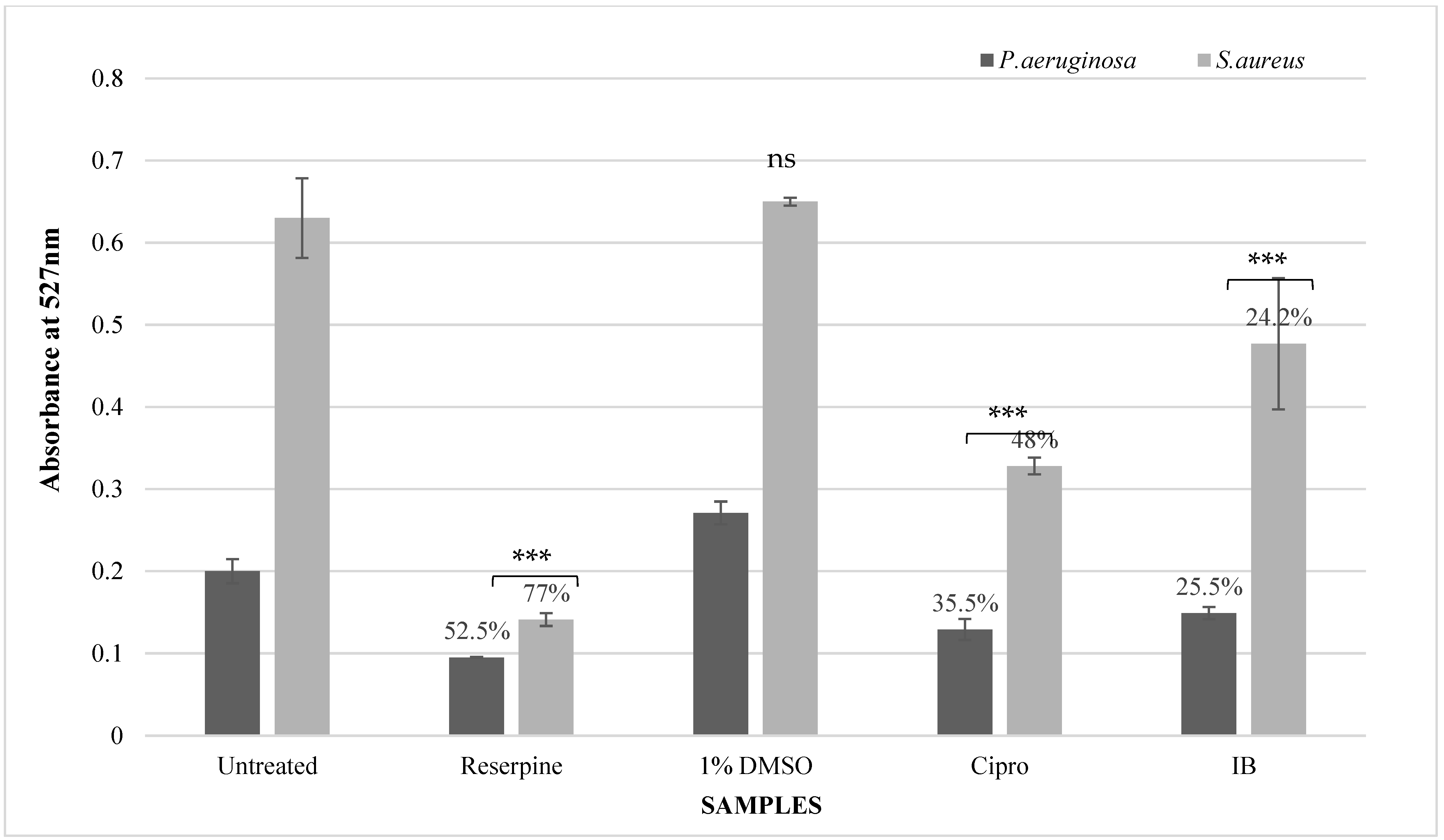
3.4.2. Rhodamine-6-G Uptake Assay
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reygaert, W.C. An Overview of the Antimicrobial Resistance Mechanisms of Bacteria. AIMS Microbiol. 2018, 4, 482–501. [Google Scholar] [CrossRef]
- Vasconcelos, S.E.C.B.; Melo, H.M.; Cavalcante, T.T.A.; Júnior, F.E.A.C.; de Carvalho, M.G.; Menezes, F.G.R.; de Sousa, O.V.; Costa, R.A. Plectranthus amboinicus Essential Oil and Carvacrol Bioactive against Planktonic and Biofilm of Oxacillin- and Vancomycin-Resistant Staphylococcus aureus. BMC Complement. Altern. Med. 2017, 17, 462. [Google Scholar] [CrossRef] [Green Version]
- Yuan, F.; Yin, S.; Xu, Y.; Xiang, L.; Wang, H.; Li, Z.; Fan, K.; Pan, G. The Richness and Diversity of Catalases in Bacteria. Front. Microbiol. 2021, 12, 5477. [Google Scholar] [CrossRef] [PubMed]
- Johnson, L.A.; Hug, L.A. Distribution of Reactive Oxygen Species Defense Mechanisms across Domain Bacteria. Free. Radic. Biol. Med. 2019, 140, 93–102. [Google Scholar] [CrossRef]
- Juan, C.A.; Pérez de la Lastra, J.M.; Plou, F.J.; Pérez-Lebeña, E. The Chemistry of Reactive Oxygen Species (ROS) Revisited: Outlining Their Role in Biological Macromolecules (DNA, Lipids and Proteins) and Induced Pathologies. Int. J. Mol. Sci. 2021, 22, 4642. [Google Scholar] [CrossRef]
- Sánchez, E.; García, S.; Heredia, N. Extracts of Edible and Medicinal Plants Damage Membranes of Vibrio cholerae. Appl. Environ. Microbiol. 2010, 76, 6888–6894. [Google Scholar] [CrossRef] [Green Version]
- Chitemerere, T.A.; Mukanganyama, S. Evaluation of Cell Membrane Integrity as a Potential Antimicrobial Target for Plant Products. BMC Complement. Altern. Med. 2014, 14, 278. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, V.; Ahmad, A. Abrogation of Pathogenic Attributes in Drug Resistant Candida Auris Strains by Farnesol. PLoS ONE 2020, 15, e0233102. [Google Scholar] [CrossRef] [PubMed]
- Gbelska, Y.; Toth Hervay, N.; Dzugasova, V.; Konecna, A. Measurement of Energy-Dependent Rhodamine 6G Efflux in Yeast Species. Bio. Protoc. 2017, 7, 2428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arumugam, G.; Swamy, M.K.; Sinniah, U.R. Plectranthus amboinicus (Lour.) Spreng: Botanical, Phytochemical, Pharmacological and Nutritional Significance. Molecules 2016, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alpaslan, E.; Geilich, B.M.; Yazici, H.; Webster, T.J. PH-Controlled Cerium Oxide Nanoparticle Inhibition of Both Gram-Positive and Gram-Negative Bacteria Growth. Sci. Rep. 2017, 7, 58–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erny Sabrina, M.N.; Razali, M.; Mirfat, A.H.S.; Mohd Shukri, M.A. Antimicrobial Activity and Bioactive Evaluation of Plectranthus amboinicus Essential Oil. Am. J. Res. Commun. 2014, 2, 121–127. [Google Scholar]
- Lukhoba, C.W.; Simmonds, M.S.J.; Paton, A.J. Plectranthus: A Review of Ethnobotanical Uses. J. Ethnopharmacol. 2006, 103, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Rice, L.J.; Brits, G.J.; Potgieter, C.J.; Van Staden, J. Plectranthus: A Plant. Future? S. Afr. J. Bot. 2011, 77, 947–959. [Google Scholar] [CrossRef] [Green Version]
- Sawant, S.; Baldwin, T.C.; Khan, H.; Rahman, A. Evaluation of the Effect of Leaf Development in Plectranthus Amboinicus, L. on Antimicrobial Activity and Virulence Factors of Pseudomonas Aeruginosa PAO1 and Staphylococcus aureus NCTC8325. Curr. Microbiol. 2023, 80, 24. [Google Scholar] [CrossRef]
- Olwal, C.O.; Ang’ienda, P.O.; Ochiel, D.O. Alternative Sigma Factor B (σ B ) and Catalase Enzyme Contribute to Staphylococcus Epidermidis Biofilm’s Tolerance against Physico-Chemical Disinfection. Sci. Rep. 2019, 9, 7–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsaturyan, V.; Poghosyan, A.; Toczyłowski, M.; Pepoyan, A. Evaluation of Malondialdehyde Levels, Oxidative Stress and Host–Bacteria Interactions: Escherichia Coli and Salmonella Derby. Cells 2022, 11, 2989. [Google Scholar] [CrossRef] [PubMed]
- Nüsslein, K.; Arnt, L.; Rennie, J.; Owens, C.; Tew, G.N. Broad-Spectrum Antibacterial Activity by a Novel Abiogenic Peptide Mimic. Microbiology 2006, 152, 1913–1918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maesaki, S.; Marichal, P.; Bossche, H.V.; Sanglard, D.; Kohno, S. Rhodamine 6G Efflux for the Detection of CDR1-Overexpressing Azole-Resistant Candidaalbicans Strains. J. Antimicrob. Chemother. 1999, 44, 27–31. [Google Scholar] [CrossRef]
- Iwase, T.; Tajima, A.; Sugimoto, S.; Okuda, K.; Hironaka, I.; Kamata, Y.; Takada, K.; Mizunoe, Y. A Simple Assay for Measuring Catalase Activity: A Visual Approach. Sci. Rep. 2013, 3, 3081. [Google Scholar] [CrossRef] [Green Version]
- Day, W.A.; Sajecki, J.L.; Pitts, T.M.; Joens, L.A. Role of Catalase in Campylobacter jejuni Intracellular Survival. Infect. Immun. 2000, 68, 6337–6345. [Google Scholar] [CrossRef] [PubMed]
- Galvão Rodrigues, F.F.; Costa, J.G.M.; Rodrigues, F.F.G.; Campos, A.R. Study of the Interference between Plectranthus Species Essential Oils from Brazil and Aminoglycosides. Evid.-Based Complement. Altern. Med. 2013, 2013, e724161. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Conover, R.C.; Benoit, S.; Olczak, A.A.; Olson, J.W.; Johnson, M.K.; Maier, R.J. Role of a Bacterial Organic Hydroperoxide Detoxification System in Preventing Catalase Inactivation. J. Biol. Chem. 2004, 279, 51908–51914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noonpakdee, W.; Sitthimonchai, S.; Panyim, S.; Lertsiri, S. Expression of the Catalase Gene KatA in Starter Culture Lactobacillus Plantarum TISTR850 Tolerates Oxidative Stress and Reduces Lipid Oxidation in Fermented Meat Product. Int. J. Food Microbiol. 2004, 95, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Nandi, A.; Yan, L.-J.; Jana, C.K.; Das, N. Role of Catalase in Oxidative Stress- and Age-Associated Degenerative Diseases. Oxid. Med. Cell. Longev. 2019, 2019, 9613090. [Google Scholar] [CrossRef] [Green Version]
- Cosgrove, K.; Coutts, G.; Jonsson, I.-M.; Tarkowski, A.; Kokai-Kun, J.F.; Mond, J.J.; Foster, S.J. Catalase (KatA) and Alkyl Hydroperoxide Reductase (AhpC) Have Compensatory Roles in Peroxide Stress Resistance and Are Required for Survival, Persistence, and Nasal Colonization in Staphylococcus aureus. J. Bacteriol. 2007, 189, 1025–1035. [Google Scholar] [CrossRef] [Green Version]
- Silhavy, T.J.; Kahne, D.; Walker, S. The Bacterial Cell Envelope. Cold Spring Harb. Perspect. Biol. 2010, 2, a000414. [Google Scholar] [CrossRef] [PubMed]
- Wada, A.; Kono, M.; Kawauchi, S.; Takagi, Y.; Morikawa, T.; Funakoshi, K. Rapid Discrimination of Gram-Positive and Gram-Negative Bacteria in Liquid Samples by Using NaOH-Sodium Dodecyl Sulfate Solution and Flow Cytometry. PLoS ONE 2012, 7, e47093. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J. Confronting the Threat of Multidrug-Resistant Gram-Negative Bacteria in Critically Ill Patients. J. Antimicrob. Chemother. 2013, 68, 490–491. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.K.; Negi, P.S. Antibacterial Activity of Indian Borage (Plectranthus amboinicus Benth) Leaf Extracts in Food Systems and Against Natural Microflora in Chicken Meat. Food Technol. Biotechnol. 2016, 54, 90–102. [Google Scholar] [CrossRef]
- Balasopoulou, A.; Κokkinos, P.; Pagoulatos, D.; Plotas, P.; Makri, O.E.; Georgakopoulos, C.D.; Vantarakis, A.; Li, Y.; Liu, J.J.; Qi, P.; et al. Symposium Recent Advances and Challenges in the Management of Retinoblastoma Globe—Saving Treatments. BMC Ophthalmol. 2017, 17, 1. [Google Scholar]
- Nicolson, K.; Evans, G.; O’Toole, P.W. Potentiation of Methicillin Activity against Methicillin-Resistant Staphylococcus aureus by Diterpenes. FEMS Microbiol. Lett. 1999, 179, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Veras, H.N.H.; Rodrigues, F.F.G.; Colares, A.V.; Menezes, I.R.A.; Coutinho, H.D.M.; Botelho, M.A.; Costa, J.G.M. Synergistic Antibiotic Activity of Volatile Compounds from the Essential Oil of Lippia Sidoides and Thymol. Fitoterapia 2012, 83, 508–512. [Google Scholar] [CrossRef] [Green Version]
- Trombetta, D.; Castelli, F.; Sarpietro, M.G.; Venuti, V.; Cristani, M.; Daniele, C.; Saija, A.; Mazzanti, G.; Bisignano, G. Mechanisms of Antibacterial Action of Three Monoterpenes. Antimicrob. Agents Chemother. 2005, 49, 2474–2478. [Google Scholar] [CrossRef] [Green Version]
- Jimmy, J.L. Coleus Aromaticus Benth.: An Update on Its Bioactive Constituents and Medicinal Properties. All Life 2021, 14, 756–773. [Google Scholar] [CrossRef]
- Tsuchiya, H. Membrane Interactions of Phytochemicals as Their Molecular Mechanism Applicable to the Discovery of Drug Leads from Plants. Molecules 2015, 20, 18923–18966. [Google Scholar] [CrossRef] [PubMed]
- Panda, G.; Dash, S.; Sahu, S.K. Harnessing the Role of Bacterial Plasma Membrane Modifications for the Development of Sustainable Membranotropic Phytotherapeutics. Membranes 2022, 12, 914. [Google Scholar] [CrossRef] [PubMed]
- Qayyum, S.; Oves, M.; Khan, A.U. Obliteration of Bacterial Growth and Biofilm through ROS Generation by Facilely Synthesized Green Silver Nanoparticles. PLoS ONE 2017, 12, e0181363. [Google Scholar] [CrossRef] [Green Version]
- Chovanová, R.; Mikulášová, M.; Vaverková, Š. In Vitro Antibacterial and Antibiotic Resistance Modifying Effect of Bioactive Plant Extracts on Methicillin-Resistant Staphylococcus epidermidis. Int. J. Microbiol. 2013, 2013, 760969. [Google Scholar] [CrossRef] [Green Version]
- Phatthalung, P.N.; Chusri, S.; Voravuthikunchai, S.P. Thai Ethnomedicinal Plants as Resistant Modifying Agents for Combating Acinetobacter baumannii Infections. BMC Complement. Altern. Med. 2012, 12, 56. [Google Scholar] [CrossRef] [Green Version]
- Jin, J.; Zhang, J.; Guo, N.; Feng, H.; Li, L.; Liang, J.; Sun, K.; Wu, X.; Wang, X.; Liu, M.; et al. The Plant Alkaloid Piperine as a Potential Inhibitor of Ethidium Bromide Efflux in Mycobacterium Smegmatis. J. Med. Microbiol. 2011, 60, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Demgne, O.M.F.; Damen, F.; Fankam, A.G.; Guefack, M.G.F.; Wamba, B.E.N.; Nayim, P.; Mbaveng, A.T.; Bitchagno, G.T.M.; Tapondjou, L.A.; Penlap, V.B.; et al. Botanicals and Phytochemicals from the Bark of Hypericum roeperianum (Hypericaceae) Had Strong Antibacterial Activity and Showed Synergistic Effects with Antibiotics against Multidrug-Resistant Bacteria Expressing Active Efflux Pumps. J. Ethnopharmacol. 2021, 277, e114257. [Google Scholar] [CrossRef] [PubMed]
- Khare, T.; Anand, U.; Dey, A.; Assaraf, Y.G.; Chen, Z.S.; Liu, Z.; Kumar, V. Exploring Phytochemicals for Combating Antibiotic Resistance in Microbial Pathogens. Front. Pharm. 2021, 12, 720726. [Google Scholar] [CrossRef] [PubMed]
- Holler, J.G.; Christensen, S.B.; Slotved, H.-C.; Rasmussen, H.B.; Gúzman, A.; Olsen, C.-E.; Petersen, B.; Mølgaard, P. Novel Inhibitory Activity of the Staphylococcus aureus NorA Efflux Pump by a Kaempferol Rhamnoside Isolated from Persea Lingue Nees. J. Antimicrob. Chemother. 2012, 67, 1138–1144. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Height of Foam (cm) | Percentage Change |
|---|---|---|
| Untreated culture control | 2.5 | - |
| Ciprofloxacin | 0.3 | 88% |
| Indian borage | 1 | 60% |
| 1% DMSO | 2 | 20% |
| Samples | Height of Foam (cm) | Percentage Change |
|---|---|---|
| Untreated culture control | 1 | - |
| Ciprofloxacin | No foam | - |
| Indian borage | 0.5 | 50% |
| 1% DMSO | 1 | - |
| Samples | Mean Concentration of Malondialdehyde [nm] (% Change in Lipid Peroxidation) | Std.dev | p Value | Mean Concentration of Malondialdehyde [nm] (% Change in Lipid Peroxidation) | Std.dev | p Value |
|---|---|---|---|---|---|---|
| P. aeruginosa | S. aureus | |||||
| Untreated | 571 | 0.00057 | - | 256.2 | 0 | - |
| 1% DMSO | 560 (↓ 2%) | 0.0006 | *** p < 0.001 | 230.6 (↓ 10%) | 0.0065 | *** p < 0.001 |
| Ciprofloxacin | 864 (↑ 51.31%) | 0.00057 | *** p < 0.001 | 403.5 (↑ 57%) | 0.0015 | *** p < 0.001 |
| IB | 813 (↑ 42.38%) | 0.00023 | *** p < 0.001 | 365.1 (↑ 42.5%) | 0.0005 | *** p < 0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sawant, S.; Baldwin, T.C.; Metryka, O.; Rahman, A. Evaluation of the Effect of Plectranthus amboinicus L. Leaf Extracts on the Bacterial Antioxidant System and Cell Membrane Integrity of Pseudomonas aeruginosa PA01 and Staphylococcus aureus NCTC8325. Pathogens 2023, 12, 853. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens12060853
Sawant S, Baldwin TC, Metryka O, Rahman A. Evaluation of the Effect of Plectranthus amboinicus L. Leaf Extracts on the Bacterial Antioxidant System and Cell Membrane Integrity of Pseudomonas aeruginosa PA01 and Staphylococcus aureus NCTC8325. Pathogens. 2023; 12(6):853. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens12060853
Chicago/Turabian StyleSawant, Sheeba, Timothy C. Baldwin, Oliwia Metryka, and Ayesha Rahman. 2023. "Evaluation of the Effect of Plectranthus amboinicus L. Leaf Extracts on the Bacterial Antioxidant System and Cell Membrane Integrity of Pseudomonas aeruginosa PA01 and Staphylococcus aureus NCTC8325" Pathogens 12, no. 6: 853. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens12060853