Distribution and Genetic Characterization of Border Disease Virus Circulating in Sardinian Ovine Flocks

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Distribution of Pestivirus in Ovine Flocks in Sardinia
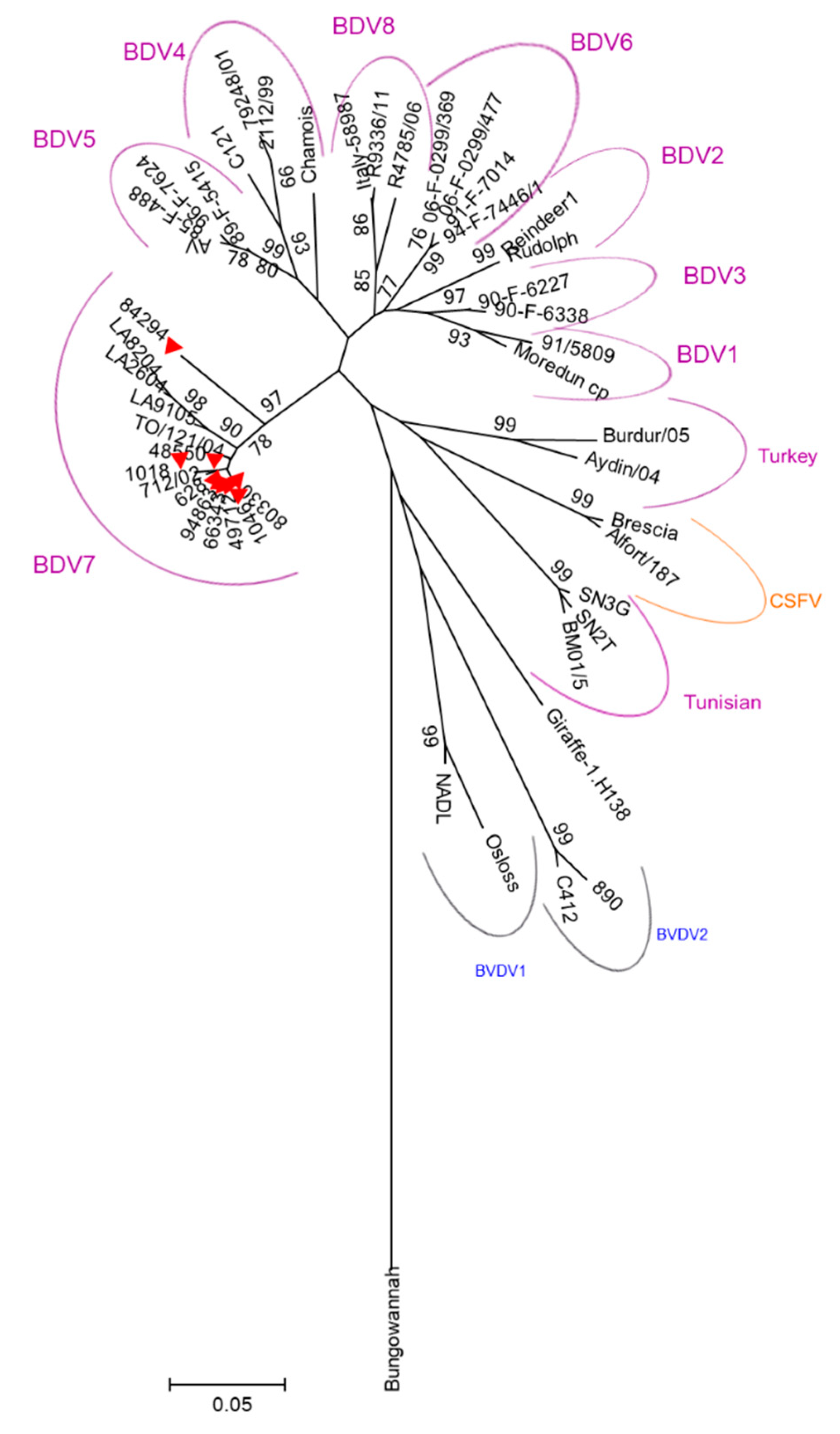
2.2. Genetic Characterization of the Circulating Strains in the Island.
3. Discussion
4. Materials and Methods
4.1. Selection of Sheep Flocks
4.2. Analysis of Samples and Collection of Data
4.3. Serological Testing
4.4. Virological Testing
4.5. Sequencing and Phylogenetic Analysis
4.6. GIS Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nettleton, P.F.; A Gilray, J.; Russo, P.; Dlissi, E. Border disease of sheep and goats. Vet. Res. 1998, 29, 327–340. [Google Scholar] [PubMed]
- Menzies, P.I. CHAPTER 90—Abortion in Sheep: Diagnosis and Control. Available online: https://0-www-sciencedirect-com.brum.beds.ac.uk/science/article/pii/B9780721693231500933 (accessed on 23 April 2020).
- Cebra, C.; Cebra, M. Chapter 16—Diseases of the Hematologic, Immunologic, and Lymphatic Systems (Multisystem Diseases). Available online: https://0-www-sciencedirect-com.brum.beds.ac.uk/science/article/pii/B9781437723533100162?via%3Dihub (accessed on 11 April 2020).
- ICTV Virus Taxonomy. Genus: Pestivirus-Flaviviridae-Positive-sense RNA Viruses-International Committee on Taxonomy of Viruses (ICTV). Available online: https://talk.ictvonline.org/ictv-reports/ictv_online_report/positive-sense-rna-viruses/w/flaviviridae/361/genus-pestivirus (accessed on 14 April 2020).
- Vilcek, S.; Nettleton, P.F.; Paton, D.J.; Belák, S. Molecular characterization of ovine pestiviruses. J. Gen. Virol. 1997, 78, 725–735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becher, P.; König, M.; Shannon, A.D.; Orlich, M.; Thiel, H.J.; Horner, G. Phylogenetic analysis of pestiviruses from domestic and wild ruminants. J. Gen. Virol. 1997, 78, 1357–1366. [Google Scholar] [CrossRef] [PubMed]
- Vilcek, S.; Paton, D.J.; Durkovic, B.; Strojny, L.; Ibata, G.; Moussa, A.; Loitsch, A.; Rossmanith, W.; Vega, S.; Scicluna, M.T.; et al. Bovine viral diarrhoea virus genotype 1 can be separated into at least eleven genetic groups. Arch. Virol. 2001, 146, 99–115. [Google Scholar] [CrossRef] [PubMed]
- Valdazo-Gonzalez, B.; Álvarez-Martínez, M.; Sandvik, T. Genetic and antigenic typing of border disease virus isolates in sheep from the Iberian Peninsula. Vet. J. 2007, 174, 316–324. [Google Scholar] [CrossRef]
- Giammarioli, M.; La Rocca, S.A.; Steinbach, F.; Casciari, C.; De Mia, G.M. Genetic and antigenic typing of border disease virus (BDV) isolates from Italy reveals the existence of a novel BDV group. Vet. Microbiol. 2011, 147, 231–236. [Google Scholar] [CrossRef]
- Schirrmeier, H.; Strebelow, G.; Depner, K.; Hoffmann, B.; Beer, M. Genetic and antigenic characterization of an atypical pestivirus isolate, a putative member of a novel pestivirus species. J. Gen. Virol. 2004, 85, 3647–3652. [Google Scholar] [CrossRef]
- Stalder, H.; Meier, P.; Pfaffen, G.; Canal, C.W.; Rufenacht, J.; Schaller, P.; Bachofen, C.; Marti, S.; Vogt, H.; Peterhans, E. Genetic heterogeneity of pestiviruses of ruminants in Switzerland. Prev. Vet. Med. 2005, 72, 37–41. [Google Scholar] [CrossRef]
- Becher, P.; Ramirez, R.A.; Orlich, M.; Cedillo-Rosales, S.; König, M.; Schweizer, M.; Stalder, H.; Schirrmeier, H.; Thiel, H.-J. Genetic and antigenic characterization of novel pestivirus genotypes: Implications for classification. Virology 2003, 311, 96–104. [Google Scholar] [CrossRef] [Green Version]
- Arnal, M.C.; Fernández-De-Luco, D.; Riba, L.; Maley, M.; Gilray, J.; Willoughby, K.; Vilcek, S.; Nettleton, P.F. A novel pestivirus associated with deaths in Pyrenean chamois (Rupicapra pyrenaica pyrenaica). J. Gen. Virol. 2004, 85, 3653–3657. [Google Scholar] [CrossRef]
- Dubois, E.; Russo, P.; Prigent, M.; Thiéry, R. Genetic characterization of ovine pestiviruses isolated in France, between 1985 and 2006. Vet. Microbiol. 2008, 130, 69–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caruso, C.; Peletto, S.; Cerutti, F.; Modesto, P.; Robetto, S.; Domenis, L.; Masoero, L.; Acutis, P.L. Evidence of circulation of the novel border disease virus genotype 8 in chamois. Arch. Virol. 2016, 162, 511–515. [Google Scholar] [CrossRef]
- Peletto, S.; Caruso, C.; Cerutti, F.; Modesto, P.; Zoppi, S.; Dondo, A.; Acutis, P.L.; Masoero, L. A new genotype of border disease virus with implications for molecular diagnostics. Arch. Virol. 2015, 161, 471–477. [Google Scholar] [CrossRef] [PubMed]
- Stalder, H.; Marti, S.; Flückiger, F.; Renevey, N.; Hofmann, M.A.; Schweizer, M. Complete Genome Sequences of Three Border Disease Virus Strains of the Same Subgenotype, BDSwiss, Isolated from Sheep, Cattle, and Pigs in Switzerland. Genome Announc. 2017, 5, e01238-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buonavoglia, C.; Marsilio, F.; Tempesta, M.; Buonavoglia, D.; Cavalli, A. Persistent pestivirus infection in sheep in Apulia (southern Italy). New Microbiol. 1994, 17, 163–165. [Google Scholar]
- Pratelli, A.; Bollo, E.; Martella, V.; Guarda, F.; Chiocco, D.; Buonavoglia, C. Pestivirus infection in small ruminants: Virological and histopathological findings. New Microbiol. 1999, 22, 351–356. [Google Scholar]
- Giovannetti, Y. I Numeri Dell’Allevamento Ovino in Italia. Available online: https://www.ruminantia.it/i-numeri-dellallevamento-ovino-in-italia/ (accessed on 13 April 2020).
- Caldelli, M.; Giannone, M. Allevamento Ovino in Toscana e Razza Sarda. Available online: http://www.rivistadiagraria.org/articoli/anno-2008/allevamento-ovino-in-toscana-e-razza-sarda/ (accessed on 13 April 2020).
- Rossi, A. CLAL-Italia: Export Pecorino e Fiore Sardo. Available online: https://www.clal.it/index.php?section=imp_exp_istat&cod=04069063&mov=E (accessed on 14 April 2020).
- Austin, P.C.; Hux, J.E. A brief note on overlapping confidence intervals. J. Vasc. Surg. 2002, 36, 194–195. [Google Scholar] [CrossRef] [Green Version]
- Felsenstein, J. Confidence Limits on Phylogenies: An Approach Using the Bootstrap. Evology 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Berriatua, E.; Barandika, J.; Aduriz, G.; Hurtado, A.; Estevez, L.; Atxaerandio, R.; García-Pérez, A. Flock-prevalence of border disease virus infection in Basque dairy-sheep estimated by bulk-tank milk analysis. Vet. Microbiol. 2006, 118, 37–46. [Google Scholar] [CrossRef]
- Corbière, F.; Pouget, C.; Bernardin, E.; Brugidou, R.; Schelcher, F. Short communication: Performance of a blocking antibody ELISA bulk-tank milk test for detection of dairy sheep flocks exposed to border disease virus. J. Dairy Sci. 2012, 95, 6542–6545. [Google Scholar] [CrossRef]
- García-Pérez, A.; Ruiz-Fons, J.F.; Barandika, J.; Aduriz, G.; Juste, R.; Hurtado, A. Border disease virus seroprevalence correlates to antibodies in bulk-tank milk and reproductive performance of dairy sheep flocks. J. Dairy Sci. 2010, 93, 2444–2449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krametter-Frotscher, R.; Loitsch, A.; Kohler, H.; Schleiner, A.; Schiefer, P.; Mostl, K.; Golja, F.; Baumgartner, W. Serological survey for antibodies against pestiviruses in sheep in Austria. Vet. Rec. 2007, 160, 726–730. [Google Scholar] [CrossRef] [PubMed]
- Schaller, P.; Vogt, H.R.; Strasser, M.; Nettleton, P.F.; Peterhans, E.; Zanoni, R. Seroprävalenz von Maedi–Visna und border disease in der Schweiz. Schweizer Archiv für Tierheilkunde 2000, 142, 145–153. [Google Scholar] [PubMed]
- Danuser, R.; Vogt, H.-R.; Kaufmann, T.; Peterhans, E.; Zanoni, R. Seroprevalence and characterization of pestivirus infections in small ruminants and new world camelids in Switzerland. Schweizer Archiv für Tierheilkunde 2009, 151, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Feknous, N.; Hanon, J.-B.; Tignon, M.; Khaled, H.; Bouyoucef, A.; Cay, A.B. Seroprevalence of border disease virus and other pestiviruses in sheep in Algeria and associated risk factors. BMC Vet. Res. 2018, 14, 339. [Google Scholar] [CrossRef] [Green Version]
- Fihri, O.F.; Jammar, N.; Amrani, N.; El Berbri, I.; Alali, S. Sheep pestivirus in Morocco: Sero-epidemiological and molecular study. Vet. Rec. Open 2019, 6, e000324. [Google Scholar] [CrossRef] [Green Version]
- Rosamilia, A.; Grattarola, C.; Caruso, C.; Peletto, S.; Gobbi, E.; Tarello, V.; Caroggio, P.; Dondo, A.; Masoero, L.; Acutis, P.L. Detection of border disease virus (BDV) genotype 3 in Italian goat herds. Vet. J. 2014, 199, 446–450. [Google Scholar] [CrossRef]
- Giammarioli, M.; Rossi, E.; Casciari, C.; Bazzucchi, M.; Claudia, T.; De Mia, G.M. Genetic characterization of border disease virus (BDV) isolates from small ruminants in Italy. Virus Genes 2015, 50, 321–324. [Google Scholar] [CrossRef]
- Ciulli, S.; Purpari, G.; Agnello, S.; Di Marco, P.; Di Bella, S.; Volpe, E.; Mira, F.; Pinheiro, A.C.D.A.S.; Vullo, S.; Guercio, A. Evidence for Tunisian-Like Pestiviruses Presence in Small Ruminants in Italy Since 2007. Transbound. Emerg. Dis. 2016, 64, 1243–1253. [Google Scholar] [CrossRef]
- Associazione Nazionale Della Pastorizia. AssoNaPa. www.aia.it–Home. Available online: http://www.aia.it/aia-website/en/home/postdetail/news/indici-assonapa (accessed on 13 April 2020).
- Vilček, Š.; Herring, A.J.; Herring, J.A.; Nettleton, P.F.; Lowings, J.P.; Paton, D.J. Pestiviruses isolated from pigs, cattle and sheep can be allocated into at least three genogroups using polymerase chain reaction and restriction endonuclease analysis. Arch. Virol. 1994, 136, 309–323. [Google Scholar] [CrossRef]
- Hall, T.A. Bioedit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular Evolutionary Genetics Analysis Using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Mol. Boil. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Province | Flocks | Total Number of Ewes | Tested Flocks (% of Total Flocks) | Ewes | ELISA Positive Flocks | % ELISA Positive Flocks (95% CI) | Flocks with Estimated Seroprevalence >30% 10–30% | PCR Positive Flocks |
|---|---|---|---|---|---|---|---|---|
| SS | 3.460 | 928,003 | 291 (8.41) | 90.268 | 17 | 5.84 (3.68–9.16) | 7 10 | 2 |
| NU | 3.226 | 771,312 | 430 (13.33) | 104.316 | 32 | 7.44 (5.32–10.32) | 15 17 | 0 |
| OR | 2.118 | 470,560 | 189 (8.92) | 60.239 | 21 | 11.11 (7.38–16.39) | 11 10 | 1 |
| SU | 2.478 | 645,836 | 312 (12.59) | 82.606 | 51 | 16.35 (12.66–20.85) | 35 16 | 3 |
| CA | 253 | 58,370 | 64 (25.30) | 17.409 | 24 | 37.5 (26.67–49.75) | 18 6 | 3 |
| TOT | 11.535 | 2,874,081 | 1286 (11.15) | 354.838 | 145 | 11.28 (9.66–13.12) | 86 59 | 9 |
| Strain | Municipality/Province | Host | Year | Genotype | Accession Number |
|---|---|---|---|---|---|
| 628 | Barrali (SU) | Sheep | 2015 | BDV-7 | MH733598 |
| 1018 | Mandas (SU) | Sheep | 2015 | BDV-7 | MH733599 |
| 1046 | Ortacesus (SU) | Sheep | 2015 | BDV-7 | MH733600 |
| 48550 | Sedilo (OR) | Sheep | 2014 | BDV-7 | MH733601 |
| 94863 | Trinità d’Agultu (SS) | Sheep | 2014 | BDV-7 | MH733602 |
| 66343 | Siliqua (CA) | Sheep | 2014 | BDV-7 | MH733603 |
| 49712 | Perfugas (SS) | Sheep | 2014 | BDV-7 | MH733604 |
| 80330 | Las Plassas (CA) | Sheep | 2014 | BDV-7 | MH733605 |
| 84294 | Orroli (CA) | Sheep | 2014 | BDV-7 | MH733606 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piras, I.M.; Dei Giudici, S.; Fadda, M.; Anfossi, A.G.; Oggiano, A.; Pittau, M.; Chessa, B. Distribution and Genetic Characterization of Border Disease Virus Circulating in Sardinian Ovine Flocks. Pathogens 2020, 9, 360. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9050360
Piras IM, Dei Giudici S, Fadda M, Anfossi AG, Oggiano A, Pittau M, Chessa B. Distribution and Genetic Characterization of Border Disease Virus Circulating in Sardinian Ovine Flocks. Pathogens. 2020; 9(5):360. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9050360
Chicago/Turabian StylePiras, Ilaria M., Silvia Dei Giudici, Manlio Fadda, Antonio G. Anfossi, Annalisa Oggiano, Marco Pittau, and Bernardo Chessa. 2020. "Distribution and Genetic Characterization of Border Disease Virus Circulating in Sardinian Ovine Flocks" Pathogens 9, no. 5: 360. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9050360