First Isolation and Characterization of Chryseobacterium cucumeris SKNUCL01, Isolated from Diseased Pond loach (Misgurnus anguillicaudatus) in Korea


Abstract
:1. Introduction
2. Results
2.1. Isolation of Bacteria SKNUCL01
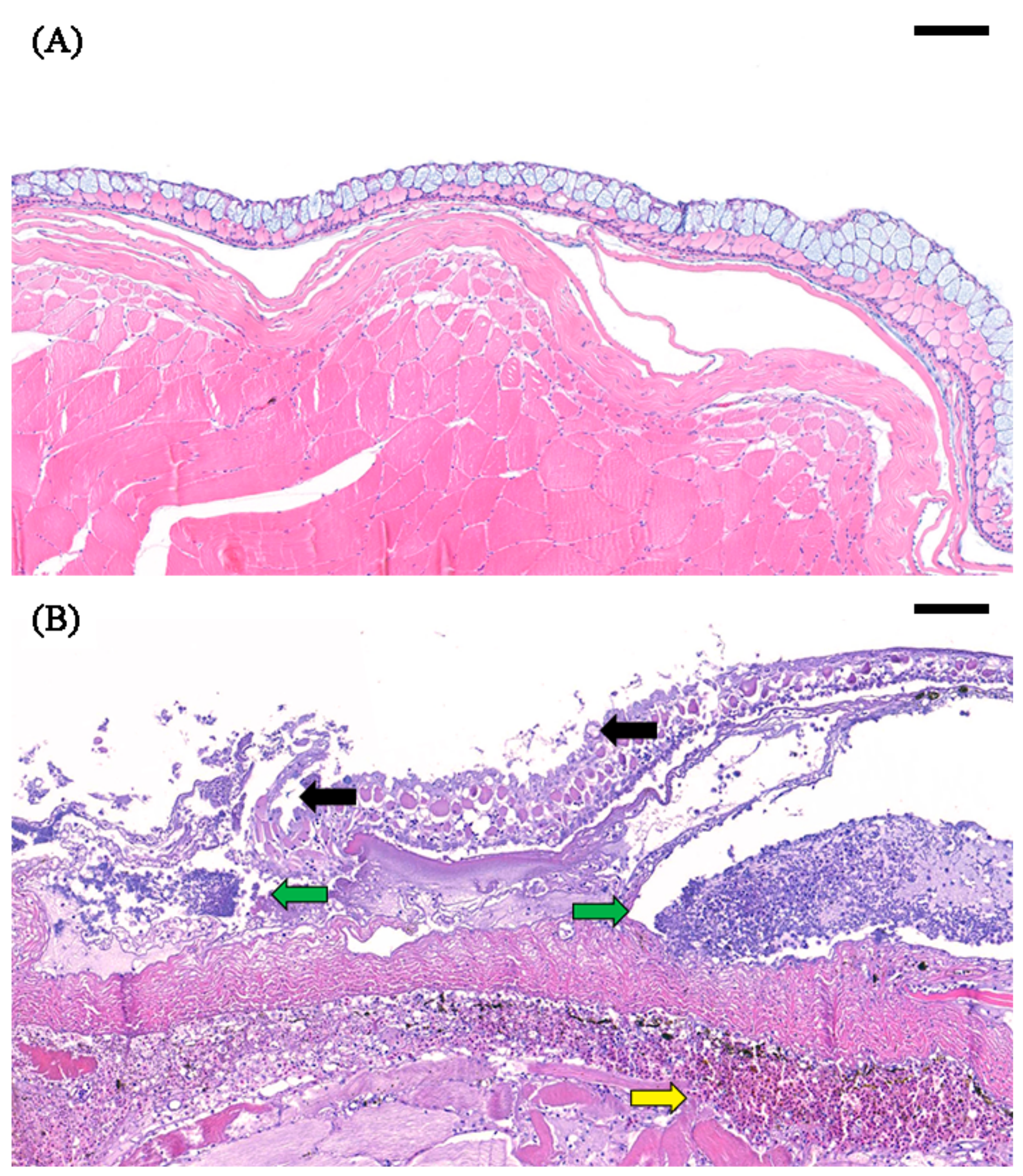
2.2. Histological Analysis of the Skin
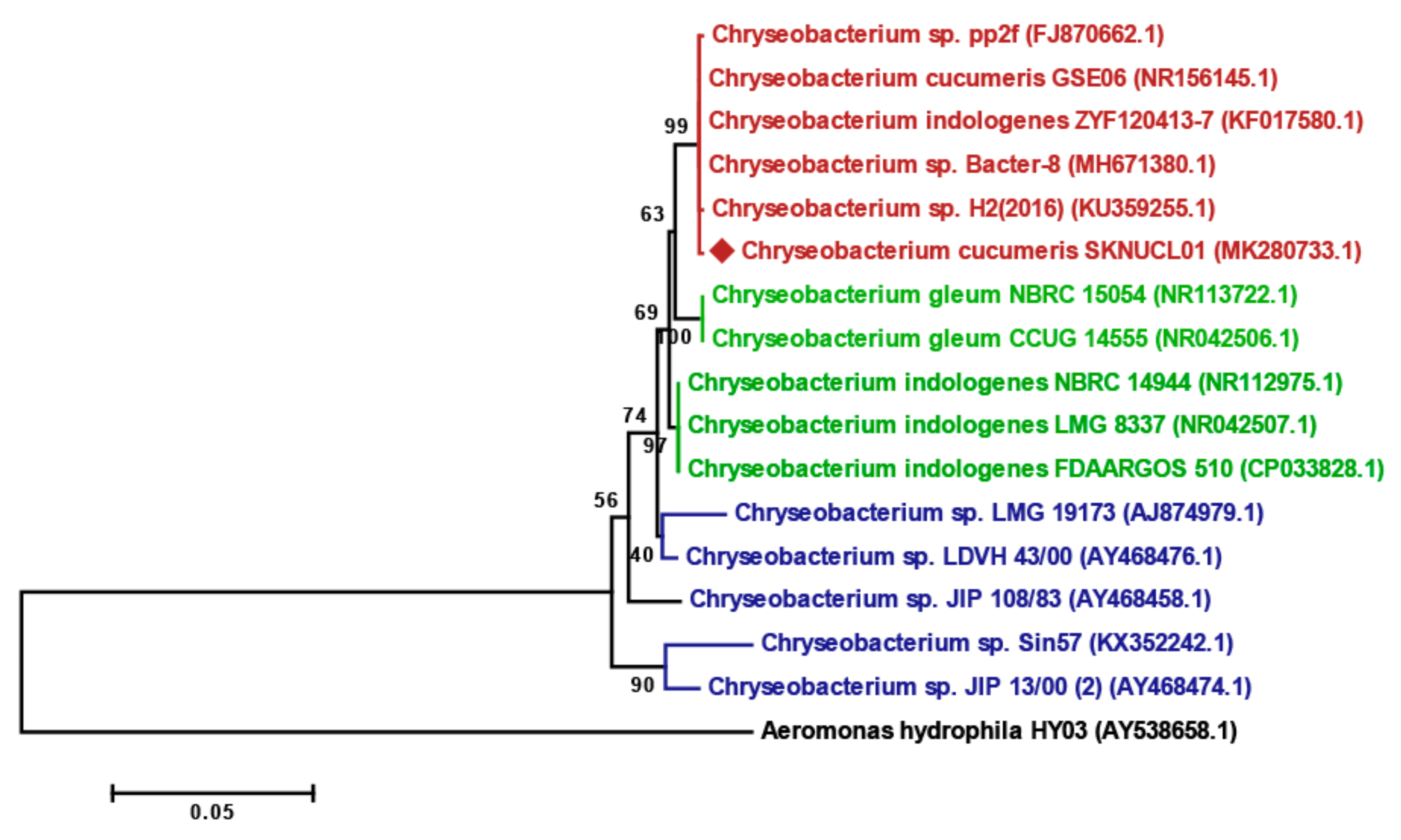
2.3. Identification of Bacteria SKNUCL01
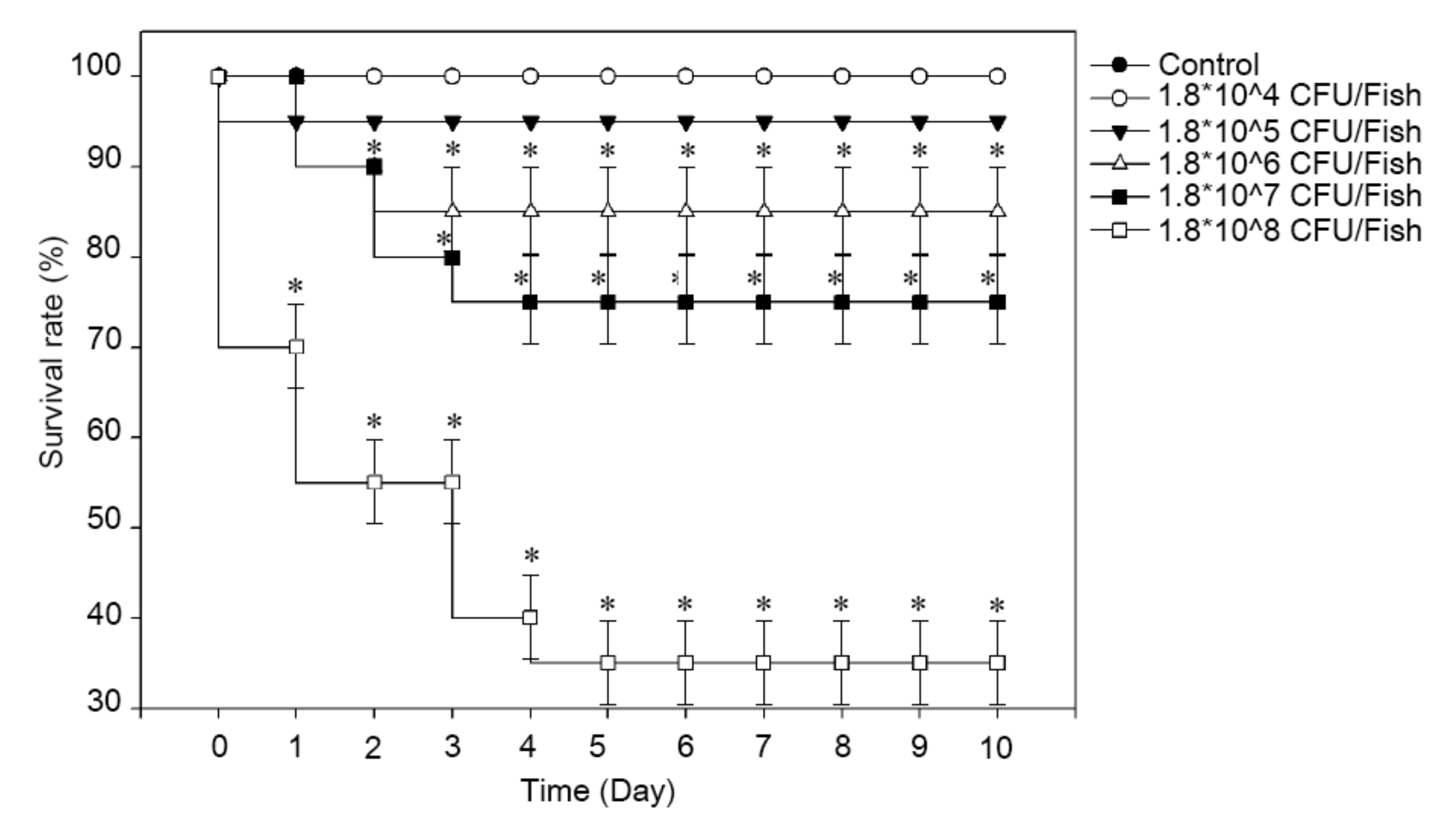
2.4. Virulence Test of SKNUCL01
2.5. Antibiotic Susceptibility Test of SKNUCL01
2.6. Phenotypic Screening for β-Lactamases
2.7. Phenotypic Screening for Efflux Pump against Antibiotics
2.8. Genetic Screening for Antibiotic Resistance
3. Discussion
4. Materials and Methods
4.1. Ethical Statement
4.2. Bacterial Isolates and Culture Condition
4.3. Histological Analysis of the Skin Lesion
4.4. Identification of Isolated Bacterial Strains
4.5. Virulence of C. cucumeris SKNUCL01 in Pond Loach
4.6. Biofilm Formation of C. cucumeris SKNUCL01
4.7. Antibiotic Susceptibility Test
4.8. Phenotypic Tests for Detection of β-Lactamases
4.9. Effect of PAβN on the MIC
4.10. Detection of Antibiotic Resistance-Related Genes
4.11. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Saito, K. Movement and spawning of several freshwater fishes in temporary waters around paddy fields. Jpn. J. Ecol. 1988, 38, 35–47. [Google Scholar]
- Kim, J.O.; Shin, H.S.; Yoo, J.H.; Lee, S.H.; Jang, K.S.; Kim, B.C. Functional evaluation of small-scale pond at paddy field as a shelter for mudfish during midsummer drainage period. Korean J. Environ. Biol. 2011, 30, 37–42. [Google Scholar]
- Zhang, H.; Lu, X.; Zhang, Y.; Ma, X.; Wang, S.; Ni, Y.; Chen, J. Bioaccumulation of organochlorine pesticides and polychlorinated biphenyls by loaches living in rice paddy fields of Northeast China. Environ. Pollut. 2016, 216, 893–901. [Google Scholar] [CrossRef] [PubMed]
- You, L.; Zhao, M.; Regenstein, J.M.; Ren, J. In vitro antioxidant activity and in vivo anti-fatigue effect of loach (Misgurnus anguillicaudatus) peptides prepared by papain digestion. Food Chem. 2011, 124, 188–194. [Google Scholar] [CrossRef]
- You, L.; Zhao, M.; Liu, R.H.; Regenstein, J.M. Antioxidant and antiproliferative activities of loach (Misgurnus anguillicaudatus) peptides prepared by papain digestion. J. Agric. Food Chem. 2011, 59, 7948–7953. [Google Scholar] [CrossRef]
- Chowdhury, M.B.R.; Wakabayashi, H. A study on Flexibacter columnaris infection in loach, Misgurnus anguillicaudatus (Bleeker, Günther). J. Fish Dis. 1991, 14, 389–394. [Google Scholar] [CrossRef]
- Zhang, X.J.; Yao, D.R.; Yan, B.L.; Bi, K.R.; Liang, L.G.; Qin, G.M. Identification of Vibrio cholerae as a causative bacterium for an ulcer disease of cultured loach Misgurnus anguillicaudatus in China. Afr. J. Microbiol. Res. 2012, 6, 2060–2070. [Google Scholar]
- Jun, J.W.; Kim, J.H.; Gomez, D.K.; Choresca, C.H.; Han, J.E.; Shin, S.P.; Park, S.C. Occurrence of tetracycline-resistant Aeromonas hydrophila infection in Korean cyprinid loach (Misgurnus anguillicaudatus). Afr. J. Microbiol. Res. 2010, 4, 849–855. [Google Scholar]
- Zhu, M.; Wang, X.R.; Li, J.; Li, G.Y.; Liu, Z.P.; Mo, Z.L. Identification and virulence properties of Aeromonas veronii bv. sobria isolates causing an ulcerative syndrome of loach Misgurnus anguillicaudatus. J. Fish Dis. 2016, 39, 777–781. [Google Scholar] [CrossRef]
- Campbell, S.; Harada, R.M.; Li, Q.X. Chryseobacterium arothri sp. nov., isolated from the kidneys of a pufferfish. Int. J. Syst. Evol. Microbiol. 2008, 58, 290–293. [Google Scholar] [CrossRef]
- Zamora, L.; Vela, A.I.; Palacios, M.A.; Domínguez, L.; Fernández-Garayzábal, J.F. First isolation and characterization of Chryseobacterium shigense from rainbow trout. BMC Vet. Res. 2012, 8, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahi, N.; Sharma, P.; Pandey, J.; Bisht, I.; Mallik, S.K. Characterization and pathogenicity study of Chryseobacterium scophthalmum recovered from gill lesions of diseased golden mahseer, Tor putitora (Hamilton, 1822) in India. Aquaculture 2018, 485, 81–92. [Google Scholar] [CrossRef]
- Bernardet, J.F.; Vancanneyt, M.; Matte-Tailliez, O.; Grisez, L.; Tailliez, P.; Bizet, C.; Nowakowskie, M.; Kerouaulta, B.; Swings, J. Polyphasic study of Chryseobacterium strains isolated from diseased aquatic animals. Syst. Appl. Microbiol. 2005, 28, 640–660. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.C.; Lo, H.H.; Hsieh, H.Y.; Chang, S.M. Identification, epidemiological relatedness, and biofilm formation of clinical Chryseobacterium indologenes isolates from central Taiwan. J. Microbiol. Immunol. Infect. 2015, 48, 559–564. [Google Scholar] [CrossRef] [Green Version]
- Lambiase, A.; Del Pezzo, M.; Raia, V.; Sepe, A.; Ferri, P.; Rossano, F. Chryseobacterium respiratory tract infections in patients with cystic fibrosis. J. Infect. 2007, 55, 518–523. [Google Scholar] [CrossRef]
- Alfouzan, W.; Dhar, R.; Al-Hashemi, H.; Al-Sweih, N.; Albert, M.J. Clinical and microbiological characteristics of Chryseobacterium spp. isolated from neonates in Kuwait. JMM Case Rep. 2014, 1, 1. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.T.; Jeng, Y.Y.; Lin, M.L.; Yu, K.W.; Wang, F.D.; Liu, C.Y. Clinical and microbiological characteristics of Chryseobacterium indologenes bacteremia. J. Microbiol. Immunol. Infect. 2010, 43, 498–505. [Google Scholar] [CrossRef] [Green Version]
- Omar, A.; Camara, M.; Fall, S.; Ngom-Cisse, S.; Fall, B.; Ba-Diallo, A.; Diop-Ndiaye, H.; Toure-Kane, C.; Mboup, S.; Gaye-Diallo, A. Chryseobacterium indologenes in a woman with acute leukemia in Senegal: A case report. J. Med. Case Rep. 2014, 8, 138. [Google Scholar] [CrossRef] [Green Version]
- Didinen, B.I.; Onuk, E.E.; Öztürk, T.; Metin, S.; Meryem, Ö.Z.; Çayli, Ö.; Kubilay, A. First report of Chryseobacterium sp. from Koi (Cyprinus carpio) in Turkey. Isr. J. Aquacult.-Bamid 2016, 68. Available online: https://www.researchgate.net/publication/316996691_First_report_of_Chryseobacterium_sp_From_koi_Cyprinus_carpio_in_Turkey (accessed on 16 May 2020).
- Michel, C.; Matte-Tailliez, O.; Kerouault, B.; Bernardet, J.F. Resistance pattern and assessment of phenicol agents’ minimum inhibitory concentration in multiple drug resistant Chryseobacterium isolates from fish and aquatic habitats. J. Appl. Microbiol. 2005, 99, 323–332. [Google Scholar] [CrossRef]
- Maravić, A.; Skočibušić, M.; Šamanić, I.; Puizina, J. Profile and multidrug resistance determinants of Chryseobacterium indologenes from seawater and marine fauna. World J. Microbiol. Biotechnol. 2013, 29, 515–522. [Google Scholar] [CrossRef] [PubMed]
- Mielmann, A. Food spoilage characteristics of Chryseobacterium species. Master’s Thesis, University of the Free State, Bloemfontein, South Africa, 2006. [Google Scholar]
- Bekker, A. Growth and spoilage characteristics of Chryseobacterium species in milk. Master’s Thesis, University of the Free State, Bloemfontein, South Africa, 2011. [Google Scholar]
- Zheng, L.; Bae, Y.M.; Jung, K.S.; Heu, S.; Lee, S.Y. Antimicrobial activity of natural antimicrobial substances against spoilage bacteria isolated from fresh produce. Food Control 2013, 32, 665–672. [Google Scholar] [CrossRef]
- Sharma, P.; Gupta, S.K.; Diene, S.M.; Rolain, J.M. Whole-genome sequence of Chryseobacterium oranimense, a colistin-resistant bacterium isolated from a cystic fibrosis patient in France. Antimicrob. Agents Chemother. 2015, 59, 1696–1706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, J.J.; Lee, D.W.; Park, B.; Sang, M.K.; Choi, I.G.; Kim, K.D. Chryseobacterium cucumeris sp. nov., an endophyte isolated from cucumber (Cucumis sativus L.) root, and emended description of Chryseobacterium arthrosphaerae. Int. J. Syst. Evol. Microbiol. 2017, 67, 610–616. [Google Scholar] [CrossRef] [PubMed]
- Qin, L.; Zhu, M.; Xu, J. First report of Shewanella sp. and Listonella sp. infection in freshwater cultured loach, Misgurnus anguillicaudatus. Aquac. Res. 2014, 45, 602–608. [Google Scholar] [CrossRef]
- Loch, T.P.; Faisal, M. Chryseobacterium aahli sp. nov., isolated from lake trout (Salvelinus namaycush) and brown trout (Salmo trutta), and emended descriptions of Chryseobacterium ginsenosidimutans and Chryseobacterium gregarium. Int. J. Syst. Evol. Microbiol. 2014, 64, 1573–1579. [Google Scholar] [CrossRef]
- Pridgeon, J.W.; Klesius, P.H.; Garcia, J.C. Identification and virulence of Chryseobacterium indologenes isolated from diseased yellow perch (Perca flavescens). J. Appl. Microbiol. 2013, 114, 636–643. [Google Scholar] [CrossRef]
- Ilardi, P.; Fernandez, J.; Avendano-Herrera, R. Chryseobacterium piscicola sp. nov., isolated from diseased salmonid fish. Int. J. Syst. Evol. Microbiol. 2009, 59, 3001–3005. [Google Scholar] [CrossRef] [Green Version]
- Menanteau-Ledouble, S.; Karsi, A.; Lawrence, M.L. Importance of skin abrasion as a primary site of adhesion for Edwardsiella ictaluri and impact on invasion and systematic infection in channel catfish Ictalurus punctatus. Vet. Microbiol. 2011, 148, 425–430. [Google Scholar] [CrossRef]
- Abraham, T.J.; Sarker, S.; Dash, G.; Patra, A.; Adikesavalu, H. Chryseobacterium sp. PLI2 and Aeromonas hydrophila co-infection in pacu, Piaractus brachypomus (Cuvier, 1817) fries cultured in West Bengal, India. Aquaculture 2017, 473, 223–227. [Google Scholar] [CrossRef]
- Pan, H.J.; Teng, L.J.; Chen, Y.C.; Hsueh, P.R.; Yang, P.C.; Ho, S.W.; Luh, K.T. High protease activity of Chryseobacterium indologenes isolates associated with invasive infection. J. Mcrobiol. Immunol. Infect. 2000, 33, 223–226. [Google Scholar]
- Snoussi, M.I.; Noumi, E.; Cheriaa, J.; Usai, D.; Sechi, L.A.; Zanetti, S.; Bakhrouf, A. Adhesive properties of environmental Vibrio alginolyticus strains to biotic and abiotic surfaces. New Microbiol. 2008, 31, 489–500. [Google Scholar]
- Kirby, J.T.; Sader, H.S.; Walsh, T.R.; Jones, R.N. Antimicrobial susceptibility and epidemiology of a worldwide collection of Chryseobacterium spp.: Report from the SENTRY Antimicrobial Surveillance Program (1997–2001). J. Clin. Microbiol. 2004, 42, 445–448. [Google Scholar] [CrossRef] [Green Version]
- Chen, F.L.; Wang, G.C.; Teng, S.O.; Ou, T.Y.; Yu, F.L.; Lee, W.S. Clinical and epidemiological features of Chryseobacterium indologenes infections: Analysis of 215 cases. J. Mcrobiol. Immunol. Infect. 2013, 46, 425–432. [Google Scholar] [CrossRef] [Green Version]
- Clark, S.E.; Jude, B.A.; Danner, G.R.; Fekete, F.A. Identification of a multidrug efflux pump in Flavobacterium johnsoniae. Vet. Res. 2009, 40, 1–10. [Google Scholar]
- Barguigua, A.; El Otmani, F.; Talmi, M.; Reguig, A.; Jamali, L.; Zerouali, K.; Timinouni, M. Prevalence and genotypic analysis of plasmid-mediated β-lactamases among urinary Klebsiella pneumoniae isolates in Moroccan community. J. Antibiot. Res. 2013, 66, 11–16. [Google Scholar] [CrossRef] [Green Version]
- Poirel, L.; Le Thomas, I.; Naas, T.; Karim, A.; Nordmann, P. Biochemical sequence analyses of GES-1, a novel class A extended-spectrum β-lactamase, and the class 1 integron In52 from Klebsiella pneumoniae. Antimicrob. Agents Chemother. 2000, 44, 622–632. [Google Scholar] [CrossRef] [Green Version]
- Mendes, R.E.; Kiyota, K.A.; Monteiro, J.; Castanheira, M.; Andrade, S.S.; Gales, A.C.; Pignatari, A.C.C.; Tufik, S. Rapid detection and identification of metallo-β-lactamase-encoding genes by multiplex real-time PCR assay and melt curve analysis. J. Clin. Microbiol. 2007, 45, 544–547. [Google Scholar] [CrossRef] [Green Version]
- Bellais, S.; Poirel, L.; Leotard, S.; Naas, T.; Nordmann, P. Genetic diversity of carbapenem-hydrolyzing metallo-β-lactamases from Chryseobacterium (Flavobacterium) indologenes. Antimicrob. Agents Chemother. 2000, 44, 3028–3034. [Google Scholar] [CrossRef] [Green Version]
- Wolter, D.J.; Khalaf, N.; Robledo, I.E.; Vázquez, G.J.; Santé, M.I.; Aquino, E.E.; Goering, R.V.; Hanson, N.D. Surveillance of carbapenem-resistant Pseudomonas aeruginosa isolates from Puerto Rican medical center hospitals: Dissemination of KPC and IMP-18 β-lactamases. Antimicrob. Agents Chemother. 2009, 53, 1660–1664. [Google Scholar] [CrossRef] [Green Version]
- Beyrouthy, R.; Robin, F.; Cougnoux, A.; Dalmasso, G.; Darfeuille-Michaud, A.; Mallat, H.; Dabboussi, F.; Hamzé, M.; Bonnet, R. Chromosome-mediated OXA-48 carbapenemase in highly virulent Escherichia coli. J. Antimicrob. Chemother. 2013, 68, 1558–1561. [Google Scholar] [CrossRef] [Green Version]
- Monteiro, J.; Widen, R.H.; Pignatari, A.C.; Kubasek, C.; Silbert, S. Rapid detection of carbapenemase genes by multiplex real-time PCR. J. Antimicrob. Chemother. 2012, 67, 906–909. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, T.; Nagata, M.; Ishimine, N.; Kawasaki, K.; Yamauchi, K.; Hidaka, E.; Kasuga, E.; Horiuchi, K.; Oana, K.; Kawakami, Y.; et al. Characterization of CIA-1, an Ambler class A extended-spectrum β-lactamase from Chryseobacterium indologenes. Antimicrob. Agents Chemother. 2012, 56, 588–590. [Google Scholar] [CrossRef] [Green Version]
- Bellais, S.; Naas, T.; Nordmann, P. Molecular and biochemical characterization of Ambler class A extended-spectrum β-lactamase CGA-1 from Chryseobacterium gleum. Antimicrob. Agents Chemother. 2002, 46, 966–970. [Google Scholar] [CrossRef] [Green Version]
- Swick, M.C.; Morgan-Linnell, S.K.; Carlson, K.M.; Zechiedrich, L. Expression of multidrug efflux pump genes acrAB-tolC, mdfA, and norE in Escherichia coli clinical isolates as a function of fluoroquinolone and multidrug resistance. Antimicrob. Agents Chemother. 2011, 55, 921–924. [Google Scholar] [CrossRef] [Green Version]
- Olah, P.A.; Doetkott, C.; Fakhr, M.K.; Logue, C.M. Prevalence of the Campylobacter multi-drug efflux pump (CmeABC) in Campylobacter spp. isolated from freshly processed turkeys. Food Microbiol. 2006, 23, 453–460. [Google Scholar] [CrossRef]
- Murugan, N.; Malathi, J.; Therese, K.L.; Madhavan, H.N. Application of six multiplex PCR’s among 200 clinical isolates of Pseudomonas aeruginosa for the detection of 20 drug resistance encoding genes. Kaohsiung J. Med. Sci. 2018, 34, 79–88. [Google Scholar] [CrossRef]
- Dweba, C.C.; Zishiri, O.T.; El Zowalaty, M.E. Isolation and molecular identification of virulence, antimicrobial and heavy metal resistance genes in livestock-associated methicillin-resistant Staphylococcus aureus. Pathogens 2019, 8, 79. [Google Scholar] [CrossRef] [Green Version]
- Lane, D.J. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; John Wiley and Sons: Chichester, UK, 1991; pp. 115–175. [Google Scholar]
- Chun, J.; Lee, J.H.; Jung, Y.; Kim, M.; Kim, S.; Kim, B.K.; Lim, Y.W. EzTaxon: A web-based tool for the identification of prokaryotes based on 16S ribosomal RNA gene sequences. Int. J. Syst. Evol. Microbiol. 2007, 57, 2259–2261. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Miller, L.C.; Tainter, M. Estimation of the ED50 and its error by means of logarithmic-probit graph paper. Proc. Soc. Exp. Biol. Med. 1944, 57, 261–264. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing: Twenty-Fourth Informational Supplement, M100-S24, 24th ed.; National Committee for Clinical Laboratory Standards (NCCLS): Wayne, PA, USA, 2014. [Google Scholar]
- Franklin, C.; Liolios, L.; Peleg, A.Y. Phenotypic detection of carbapenem-susceptible metallo-β-lactamase-producing gram-negative bacilli in the clinical laboratory. J. Clin. Microbiol. 2006, 44, 3139–3144. [Google Scholar] [CrossRef] [Green Version]
- Polsfuss, S.; Bloemberg, G.V.; Giger, J.; Meyer, V.; Böttger, E.C.; Hombach, M. Practical approach for reliable detection of AmpC beta-lactamase-producing Enterobacteriaceae. J. Clin. Microbiol. 2011, 49, 2798–2803. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.; Kim, C.K.; Yong, D.; Jeong, S.H.; Yum, J.H.; Seo, Y.H.; Docquier, J.D.; Chong, Y. Improved performance of the modified Hodge test with MacConkey agar for screening carbapenemase-producing Gram-negative bacilli. J. Microbiol. Methods 2010, 83, 149–152. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Antimicrobial Agent | Interpretation Range a | MIC c | ||
|---|---|---|---|---|
| S b | I | R | ||
| Piperacillin | ≤16 | 32–64 | ≥128 | 64 |
| Piperacillin/ tazobactam | ≤16/4 | 32/4–64/4 | ≥128/4 | ≤4/4 |
| Ceftazidime | ≤8 | 16 | ≥32 | 4 |
| Cefepime | ≤8 | 16 | ≥32 | ≤1 |
| Cefotaxime | ≤8 | 16-32 | ≥64 | ≥64 |
| Aztreonam | ≤8 | 16 | ≥32 | ≥64 |
| Imipenem | ≤4 | 8 | ≥16 | ≥16 |
| Meropenem | ≤4 | 8 | ≥16 | ≥16 |
| Amikacin | ≤16 | 32 | ≥64 | ≥64 |
| Gentamicin | ≤4 | 8 | ≥16 | ≥16 |
| Minocycline | ≤4 | 8 | ≥16 | 8 |
| Ciprofloxacin | ≤1 | 2 | ≥4 | ≥4 |
| Levofloxacin | ≤2 | 4 | ≥8 | ≥8 |
| Trimethoprim/ sulfamethoxazole | ≤2/38 | - | ≥4/76 | ≥4/76 |
| CDT a | DDST a | MHT b | |
|---|---|---|---|
| Ceftazidime | 0 | 0 | - |
| Ceftriaxone | 3 | 0 | - |
| Cefepime | 8 | 0 | - |
| Cefotaxime | 18 | 0 | - |
| Imipenem | 3 | 0 | - |
| meropenem | 2 | 0 | - |
| Antimicrobial Agents | MIC | |||
|---|---|---|---|---|
| Control | PAβN 25 μg/mL | PAβN 50 μg/mL | PAβN 100 μg/mL | |
| Piperacillin | 64 | 32 | 16 * | 8 * |
| Piperacillin/tazobactam | ≤4/4 | ≤4/4 | ≤4/4 | ≤4/4 |
| Ceftazidime | 4 | 4 | 4 | 2 |
| Cefepime | ≤1 | ≤1 | ≤1 | ≤1 |
| Cefotaxime | ≥64 | ≥64 | ≥64 | ≥64 |
| Aztreonam | ≥64 | ≥64 | ≥64 | ≥64 |
| Imipenem | ≥16 | ≥16 | 8 | 4 * |
| Meropenem | ≥16 | ≥16 | ≥16 | ≥16 |
| Amikacin | ≥64 | ≥64 | ≥64 | ≥64 |
| Gentamicin | ≥16 | ≥16 | ≥16 | ≥16 |
| Minocycline | 8 | 4 | 4 | 2 * |
| Ciprofloxacin | ≥4 | ≥4 | ≥4 | ≥4 |
| Levofloxacin | ≥8 | ≥8 | ≥8 | ≥8 |
| Trimethoprim/sulfamethoxazole | ≥4/76 | ≥4/76 | ≥4/76 | ≥4/76 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.G.; Giri, S.S.; Kim, S.W.; Kwon, J.; Lee, S.B.; Park, S.C. First Isolation and Characterization of Chryseobacterium cucumeris SKNUCL01, Isolated from Diseased Pond loach (Misgurnus anguillicaudatus) in Korea. Pathogens 2020, 9, 397. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9050397
Kim SG, Giri SS, Kim SW, Kwon J, Lee SB, Park SC. First Isolation and Characterization of Chryseobacterium cucumeris SKNUCL01, Isolated from Diseased Pond loach (Misgurnus anguillicaudatus) in Korea. Pathogens. 2020; 9(5):397. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9050397
Chicago/Turabian StyleKim, Sang Guen, Sib Sankar Giri, Sang Wha Kim, Jun Kwon, Sung Bin Lee, and Se Chang Park. 2020. "First Isolation and Characterization of Chryseobacterium cucumeris SKNUCL01, Isolated from Diseased Pond loach (Misgurnus anguillicaudatus) in Korea" Pathogens 9, no. 5: 397. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9050397