Infection Dynamics of Mycoplasma bovis and Other Respiratory Mycoplasmas in Newly Imported Bulls on Italian Fattening Farms

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
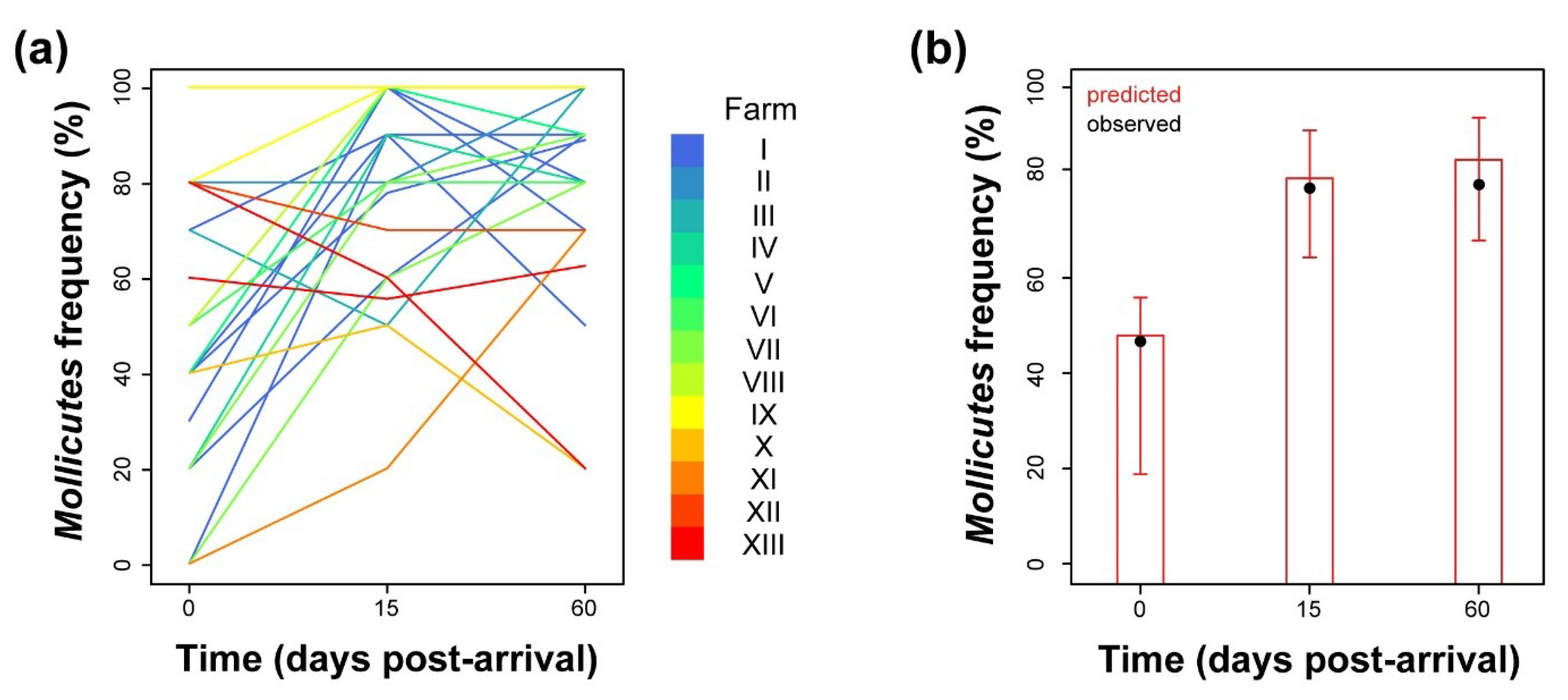
2.1. Analysis of Prevalence of Mollicutes Class Organisms
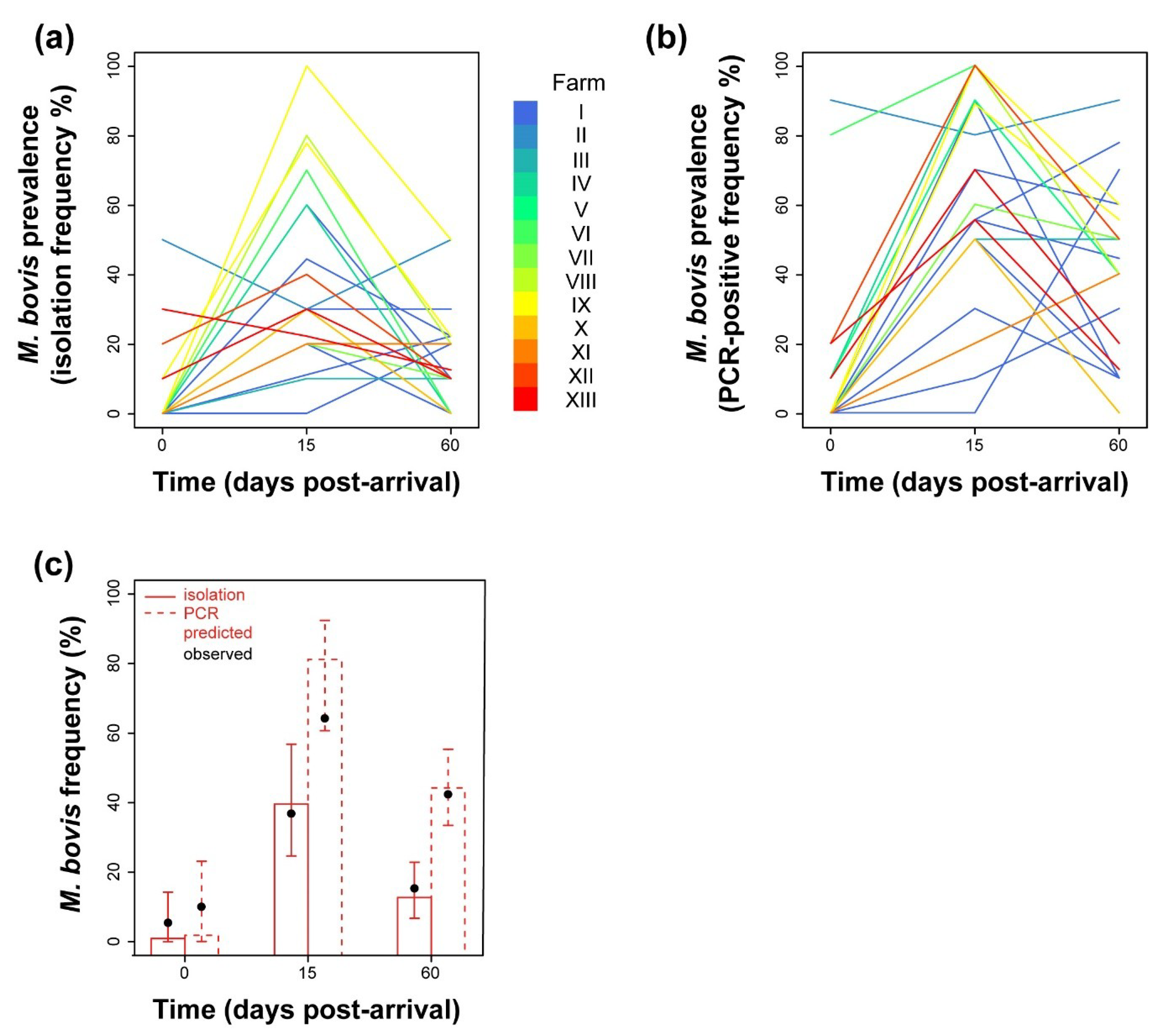
2.2. Analysis of Prevalence of M. bovis
2.3. Analysis of Prevalence of M. dispar
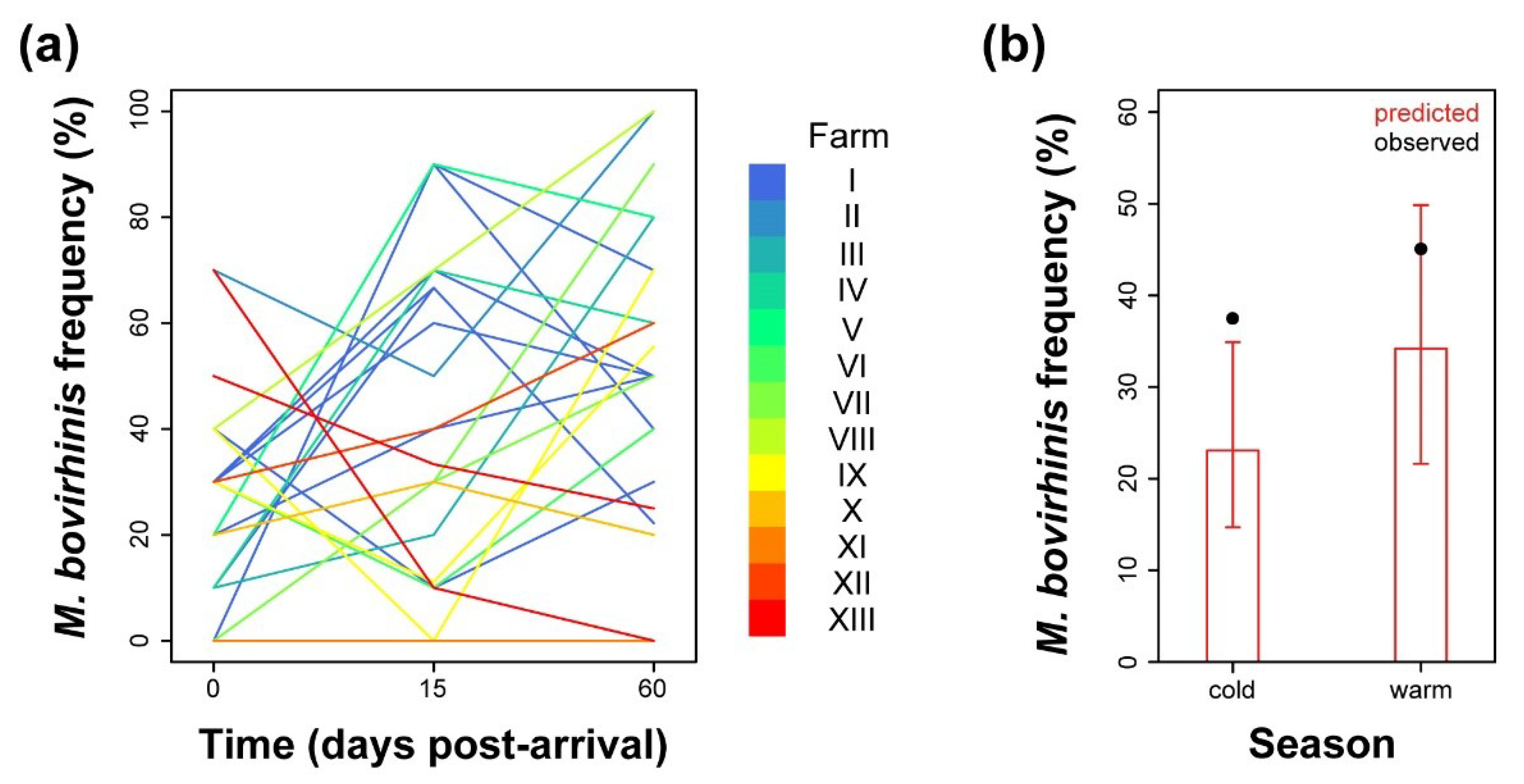
2.4. Analysis of Prevalence of M. bovirhinis
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Mollicute Cultivation
4.3. Mycoplasma Identification
4.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Cozzi, G. Present situation and future challenges of beef cattle production in Italy and the role of the research. Ital. J. Anim. Sci. 2007, 6 (Suppl. 1), 389–396. [Google Scholar] [CrossRef]
- Radaelli, E.; Luini, M.; Loria, G.R.; Nicholas, R.A.J.; Scanziani, E. Bacteriological, serological, pathological and immunohistochemical studies of Mycoplasma bovis respiratory infection in veal calves and adult cattle at slaughter. Res. Vet. Sci. 2008, 85, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Nicholas, R.A.J.; Ayling, R.D. Mycoplasma bovis: Disease, diagnosis, and control. Res. Vet. Sci. 2003, 74, 105–112. [Google Scholar] [CrossRef]
- Nicholas, R.; Ayling, R.; McAuliffe, L. Mycoplasma Diseases of Ruminants; CABI Publishing: Wallingford, UK, 2008; ISBN 9780851990125. [Google Scholar]
- Nicholas, R.A.J.; Fox, L.K.; Lysnyansky, I. Mycoplasma mastitis in cattle: To cull or not to cull. Vet. J. 2016, 216, 142–147. [Google Scholar] [CrossRef] [PubMed]
- Ayling, R.D.; Bashiruddin, S.E.; Nicholas, R.A.J. Mycoplasma species and related organisms isolated from ruminants in Britain between 1990 and 2000. Vet. Rec. 2004, 155, 413–416. [Google Scholar] [CrossRef] [PubMed]
- Pardon, B.; Hostens, M.; Duchateau, L.; Dewulf, J.; De Bleecker, K.; Deprez, P. Impact of respiratory disease, diarrhea, otitis and arthritis on mortality and carcass traits in white veal calves. BMC Vet. Res. 2013, 9, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timsit, E.; Arcangioli, M.-A.; Bareille, N.; Seegers, H.; Assié, S. Transmission dynamics of Mycoplasma bovis in newly received beef bulls at fattening operations. J. Vet. Diagn. Investig. 2012, 24, 1172–1176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angen, Ø.; Thomsen, J.; Larsen, L.E.; Larsen, J.; Kokotovic, B.; Heegaard, P.M.H.; Enemark, J.M.D. Respiratory disease in calves: Microbiological investigations on trans-tracheally aspirated bronchoalveolar fluid and acute phase protein response. Vet. Microbiol. 2009, 137, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Bell, C.J.; Blackburn, P.; Elliott, M.; Patterson, T.I.A.P.; Ellison, S.; Lahuerta-Marin, A.; Ball, H.J. Investigation of polymerase chain reaction assays to improve detection of bacterial involvement in bovine respiratory disease. J. Vet. Diagn. Investig. 2014, 26, 631–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Booker, C.W.; Abutarbush, S.M.; Morley, P.S.; Jim, G.K.; Pittman, T.J.; Schunicht, O.C.; Perrett, T.; Wildman, B.K.; Fenton, R.K.; Guichon, P.T.; et al. Microbiological and histopathological findings in cases of fatal bovine respiratory disease of feedlot cattle in western Canada. Can. Vet. J. 2008, 49, 473. [Google Scholar] [PubMed]
- O’Neill, R.; Mooney, J.; Connaghan, E.; Furphy, C.; Graham, D.A. Patterns of detection of respiratory viruses in nasal swabs from calves in Ireland: A retrospective study. Vet. Rec. 2014, 175, 35. [Google Scholar] [CrossRef] [PubMed]
- Stein, R.A.; Katz, D.E. Escherichia coli, cattle and the propagation of disease. FEMS Microbiol. Lett. 2017, 364, 1–11. [Google Scholar] [CrossRef]
- Abdelhay Kaoud, H. Introductory Chapter: Bacterial Cattle Diseases—Economic Impact and Their Control. In Bacterial Cattle Diseases; IntechOpen: London, UK, 2019. [Google Scholar]
- Catania, S.; Gobbo, F.; Bilato, D.; Gagliazzo, L.; Moronato, M.L.; Terregino, C.; Bradbury, J.M.; Ramírez, A.S. Two strains of Mycoplasma synoviae from chicken flocks on the same layer farm differ in their ability to produce eggshell apex abnormality. Vet. Microbiol. 2016, 193, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Catania, S.; Bilato, D.; Gobbo, F.; Granato, A.; Terregino, C.; Iob, L.; Nicholas, R.A.J. Treatment of Eggshell Abnormalities and Reduced Egg Production Caused by Mycoplasma synoviae Infection. Avian Dis. 2010, 54, 961–964. [Google Scholar] [CrossRef] [PubMed]
- McAuliffe, L.; Ellis, R.J.; Lawes, J.R.; Ayling, R.D.; Nicholas, R.A.J. 16S rDNA PCR and denaturing gradient gel electrophoresis; a single generic test for detecting and differentiating Mycoplasma species. J. Med. Microbiol. 2005, 54, 731–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butler, J.A.; Pinnow, C.C.; Thomson, J.U.; Levisohn, S.; Rosenbusch, R.F. Use of arbitrarily primed polymerase chain reaction to investigate Mycoplasma bovis outbreaks. Vet. Microbiol. 2001, 78, 175–181. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing 2019; R Foundation for Statistical Computing: Vienna, Austria; Available online: https://www.R-project.org/ (accessed on 1 July 2020).
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using {lme4}. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Chung, Y.; Rabe-Hesketh, S.; Dorie, V.; Gelman, A.; Liu, J. A nondegenerate penalized likelihood estimator for variance parameters in multilevel models. Psychometrika 2013, 78, 685–709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lenth, R. Emmeans: Estimated Marginal Means, Aka Least-Squares Means. R Package Version 1.4.6. 2019. Available online: https://CRAN.R-project.org/package=emmean (accessed on 1 July 2020).




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Catania, S.; Gastaldelli, M.; Schiavon, E.; Matucci, A.; Tondo, A.; Merenda, M.; Nicholas, R.A.J. Infection Dynamics of Mycoplasma bovis and Other Respiratory Mycoplasmas in Newly Imported Bulls on Italian Fattening Farms. Pathogens 2020, 9, 537. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9070537
Catania S, Gastaldelli M, Schiavon E, Matucci A, Tondo A, Merenda M, Nicholas RAJ. Infection Dynamics of Mycoplasma bovis and Other Respiratory Mycoplasmas in Newly Imported Bulls on Italian Fattening Farms. Pathogens. 2020; 9(7):537. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9070537
Chicago/Turabian StyleCatania, Salvatore, Michele Gastaldelli, Eliana Schiavon, Andrea Matucci, Annalucia Tondo, Marianna Merenda, and Robin A. J. Nicholas. 2020. "Infection Dynamics of Mycoplasma bovis and Other Respiratory Mycoplasmas in Newly Imported Bulls on Italian Fattening Farms" Pathogens 9, no. 7: 537. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9070537