BLIGHTSIM: A New Potato Late Blight Model Simulating the Response of Phytophthora infestans to Diurnal Temperature and Humidity Fluctuations in Relation to Climate Change

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
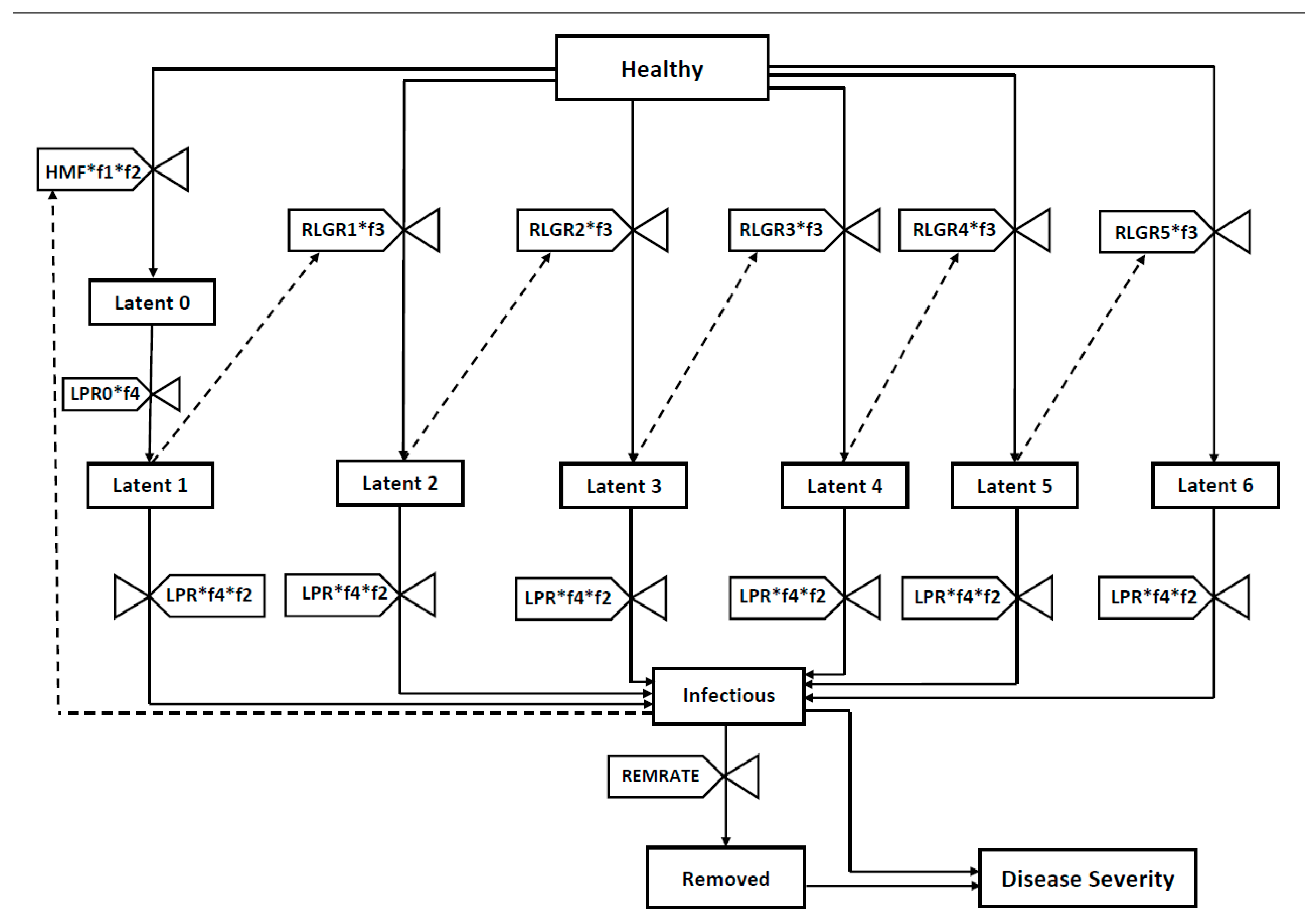
2.1. Model Development
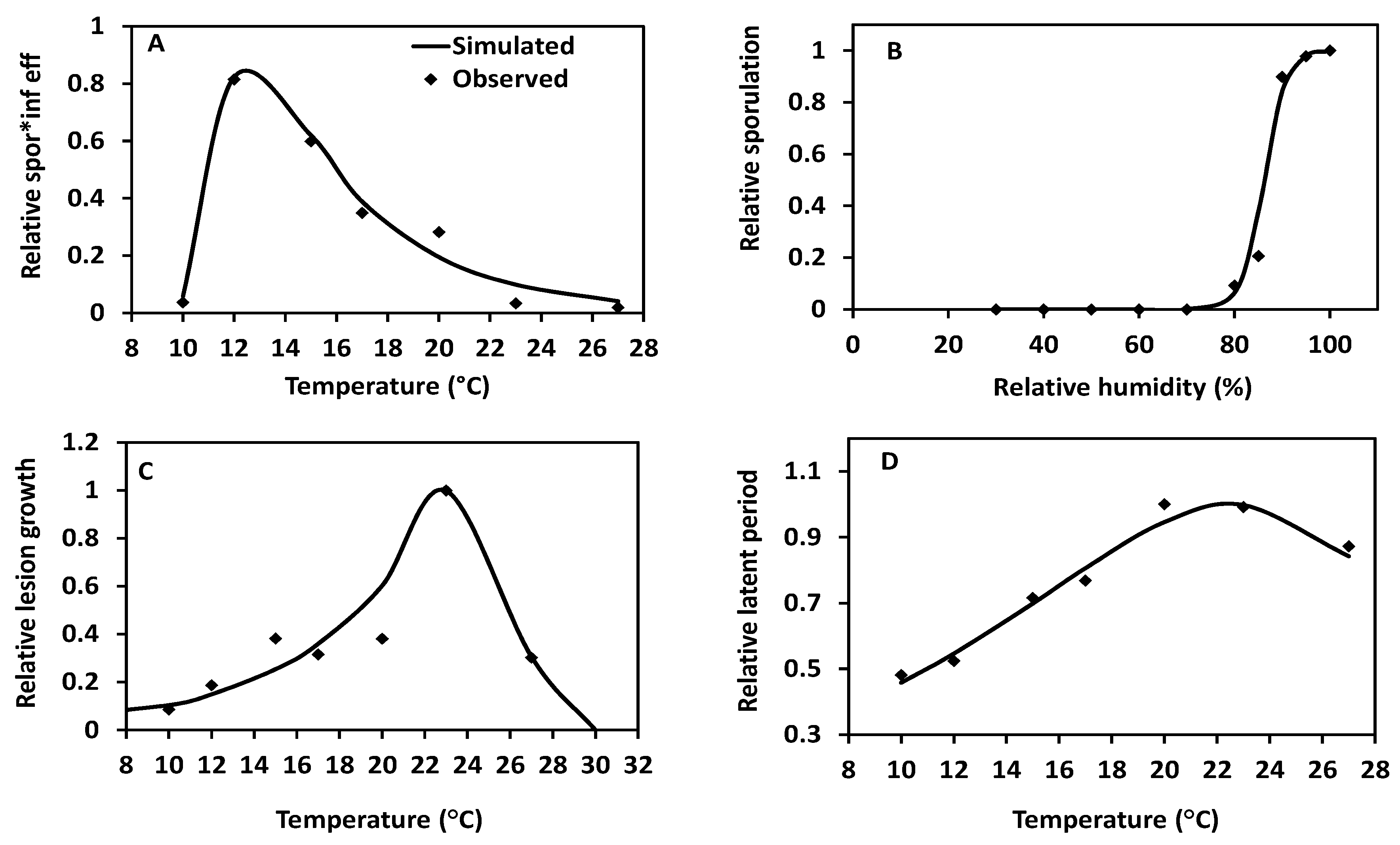
2.2. Temperature and Relative Humidity Response Curves
2.3. Model Calibration with Growth Chamber Data
2.4. Model Output Simulating Growth Chamber Conditions
2.5. Model Output Simulating Field Conditions
2.6. Scenario Testing
3. Discussion
4. Materials and Methods
4.1. Model Assumptions
4.2. Basic Model Structure
4.3. Effects of Environmental Conditions
4.3.1. Effect of Temperature on Relative SporulationXInfection and Derivation of Function f1
4.3.2. Effect of Relative Humidity on Sporulation and Derivation of Function f2
4.3.3. Effect of Temperature on Relative “Lesion Growth” and Derivation of Function f3
4.3.4. Effect of Temperature on Latency Progression Rate and Derivation of Function f4
4.4. Model Equations
(RLGR3*f3*H*L3) − (RLGR4*f3*H*L4) − (RLGR5*f3*H*L5))
(L6*LPR*f4) − (REMRATE*I))
- f1 = a reducing function that describes the effect of temperature on sporulation and infection (Table S1);
- f2 = a reducing function that describes the effect of RH on sporulation;
- f3 = a reducing function that describes the effect of temperature in radial “lesion growth”;
- f4 = a reducing function that describes the effect of temperature on the latency progression rate.
4.5. Estimation of Relative Lesion Growth Rate
4.6. Driving Variables
4.7. Model Calibration with Growth Chamber Data
4.8. Model Calibration and Validation with Field Data
4.9. Scenario Testing
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Luck, J.; Spackman, M.; Freeman, A.; Trebicki, P.; Griffiths, W.; Finlay, K.; Chakraborty, S. Climate change and diseases of food crops. Plant Pathol. 2011, 60, 113–121. [Google Scholar] [CrossRef]
- Baker, K.; Lake, T.; Benston, S.; Trenary, R.; Wharton, P.; Duynslager, L.; Kirk, W. Improved weather-based late blight risk management: Comparing models with a ten year forecast archive. J. Agric. Sci. 2015, 153, 245–256. [Google Scholar] [CrossRef]
- Fry, W.; Apple, A.; Bruhn, J. Evaluation of potato late blight forecasts modified to incorporate host resistance and fungicide weathering. Phytopathology 1983, 73, 1054–1059. [Google Scholar] [CrossRef]
- Garcia, B.I.L.; Sentelhas, P.C.; Tapia, L.R.; Sparovek, G. Climatic risk for potato late blight in the Andes region of Venezuela. Sci. Agric. 2008, 65, 32–39. [Google Scholar] [CrossRef]
- Grünwald, N.J.; Montes, G.R.; Saldaña, H.L.; Covarrubias, O.R.; Fry, W.E. Potato late blight management in the Toluca Valley: Field validation of SimCast modified for cultivars with high field resistance. Plant Dis. 2002, 86, 1163–1168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hijmans, R.J.; Forbes, G.; Walker, T. Estimating the global severity of potato late blight with GIS-linked disease forecast models. Plant Pathol. 2000, 49, 697–705. [Google Scholar] [CrossRef]
- Iglesias, I.; Escuredo, O.; Seijo, C.; Méndez, J. Phytophthora infestans prediction for a potato crop. Am. J. Potato Res. 2010, 87, 32–40. [Google Scholar] [CrossRef]
- Johnson, D.A.; Cummings, T.F.; Fox, A.D. Accuracy of rain forecasts for use in scheduling late blight management tactics in the Columbia Basin of Washington and Oregon. Plant Dis. 2015, 99, 683–690. [Google Scholar] [CrossRef] [Green Version]
- Kaukoranta, T. Impact of global warming on potato late blight: Risk, yield loss and control. Agric. Food Sci. 1996, 5, 311–327. [Google Scholar] [CrossRef]
- Apel, H.; Paudyal, M.; Richter, O. Evaluation of treatment strategies of the late blight Phytophthora infestans in Nepal by population dynamics modelling. Environ. Model. Softw. 2003, 18, 355–364. [Google Scholar] [CrossRef]
- Aylor, D.E.; Fry, W.E.; Mayton, H.; Andrade-Piedra, J.L. Quantifying the rate of release and escape of Phytophthora infestans sporangia from a potato canopy. Phytopathology 2001, 91, 1189–1196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruhn, J.; Fry, W. Analysis of potato late blight epidemiology by simulation modeling. Phytopathology 1981, 71, 612–616. [Google Scholar] [CrossRef]
- Fall, M.; Van der Heyden, H.; Brodeur, L.; Leclerc, Y.; Moreau, G.; Carisse, O. Spatiotemporal variation in airborne sporangia of Phytophthora infestans: Characterization and initiatives towards improving potato late blight risk estimation. Plant Pathol. 2015, 64, 178–190. [Google Scholar] [CrossRef]
- Henshall, W.; Shtienberg, D.; Beresford, R. A new potato late blight disease prediction model and its comparison with two previous models. N. Z. Plant Prot. 2006, 59, 150–154. [Google Scholar] [CrossRef]
- Michaelides, S. A simulation model of the fungus Phytophthora infestans (Mont) De Bary. Ecol. Model. 1985, 28, 121–137. [Google Scholar] [CrossRef]
- Raymundo, R.; Andrade-Piedra, J.; Juárez, H.; Forbes, G.; Hijmans, R.J. Towards an integrated and universal cropdisease model for potato late blight. In Late Blight: Managing the Global Threat, Proceedings of Global Initiative on Late Blight (GILB) Conference, Hamburg, Germany, 11–13 July 2002; Lizárraga, C., Ed.; International Potato Center (CIP): Lima, Peru, 2002; pp. 77–82. [Google Scholar]
- Johnson, A.C.S.; Frost, K.E.; Rouse, D.I.; Gevens, A.J. Effect of temperature on growth and sporulation of US-22, US-23, and US-24 clonal lineages of Phytophthora infestans and implications for late blight epidemiology. Phytopathology 2015, 105, 449–459. [Google Scholar] [CrossRef] [Green Version]
- Shtienberg, D.; Doster, M.; Pelletier, J.; Fry, W. Use of simulation models to develop a low-risk strategy to suppress early and late blight in potato foliage. Phytopathology 1989, 79, 590–595. [Google Scholar] [CrossRef] [Green Version]
- Skelsey, P.; Kessel, G.; Holtslag, A.; Moene, A.; Van Der Werf, W. Regional spore dispersal as a factor in disease risk warnings for potato late blight: A proof of concept. Agric. For. Meteorol. 2009, 149, 419–430. [Google Scholar] [CrossRef] [Green Version]
- Skelsey, P.; Kessel, G.; Rossing, W.; Van Der Werf, W. Parameterization and evaluation of a spatiotemporal model of the potato late blight pathosystem. Phytopathology 2009, 99, 290–300. [Google Scholar] [CrossRef] [Green Version]
- Small, I.M.; Joseph, L.; Fry, W.E. Evaluation of the BlightPro decision support system for management of potato late blight using computer simulation and field validation. Phytopathology 2015, 105, 1545–1554. [Google Scholar] [CrossRef]
- Van Oijen, M. Selection and use of a mathematical model to evaluate components of resistance to Phytophthora infestans in potato. Neth. J. Plant Pathol. 1992, 98, 192–202. [Google Scholar] [CrossRef]
- Sparks, A.H.; Forbes, G.A.; Hijmans, R.J.; Garrett, K.A. Climate change may have limited effect on global risk of potato late blight. Glob. Chang. Biol. 2014, 20, 3621–3631. [Google Scholar] [CrossRef] [PubMed]
- Krause, R.; Massie, L.; Hyre, R. Blitecast: A computerized forecast of potato late blight. Plant Dis. Rep. 1975, 59, 95–98. [Google Scholar]
- De Wolf, E.D.; Isard, S.A. Disease cycle approach to plant disease prediction. Annu. Rev. Phytopathol. 2007, 45, 203–220. [Google Scholar] [CrossRef]
- Berger, R.; Jones, J. A general model for disease progress with functions for variable latency and lesion expansion on growing host plants. Phytopathology 1985, 75, 792–797. [Google Scholar] [CrossRef]
- Andrade-Piedra, J.L.; Hijmans, R.J.; Forbes, G.A.; Fry, W.E.; Nelson, R.J. Simulation of potato late blight in the Andes. I: Modification and parameterization of the LATEBLIGHT model. Phytopathology 2005, 95, 1191–1199. [Google Scholar] [CrossRef]
- Garrett, K.A.; Forbes, G.; Savary, S.; Skelsey, P.; Sparks, A.H.; Valdivia, C.; Van Bruggen, A.; Willocquet, L.; Djurle, A.; Duveiller, E. Complexity in climate-change impacts: An analytical framework for effects mediated by plant disease. Plant Pathol. 2011, 60, 15–30. [Google Scholar] [CrossRef] [Green Version]
- Pautasso, M.; Döring, T.F.; Garbelotto, M.; Pellis, L.; Jeger, M.J. Impacts of climate change on plant diseases—Opinions and trends. Eur. J. Plant Pathol. 2012, 133, 295–313. [Google Scholar] [CrossRef] [Green Version]
- Savary, S.; Nelson, A.; Sparks, A.H.; Willocquet, L.; Duveiller, E.; Mahuku, G.; Forbes, G.; Garrett, K.A.; Hodson, D.; Padgham, J. International agricultural research tackling the effects of global and climate changes on plant diseases in the developing world. Plant Dis. 2011, 95, 1204–1216. [Google Scholar] [CrossRef] [Green Version]
- Scherm, H. Climate change: Can we predict the impacts on plant pathology and pest management? Can. J. Plant Pathol. 2004, 26, 267–273. [Google Scholar] [CrossRef]
- West, J.S.; Townsend, J.A.; Stevens, M.; Fitt, B.D. Comparative biology of different plant pathogens to estimate effects of climate change on crop diseases in Europe. Eur. J. Plant Pathol. 2012, 133, 315–331. [Google Scholar] [CrossRef] [Green Version]
- Field, C.B. Climate Change 2014–Impacts, Adaptation and Vulnerability: Regional Aspects; University Press: Cambridge, UK, 2014. [Google Scholar]
- Braganza, K.; Karoly, D.J.; Arblaster, J.M. Diurnal temperature range as an index of global climate change during the twentieth century. Geophys. Res. Lett. 2004, 31. [Google Scholar] [CrossRef]
- Lindvall, J.; Svensson, G. The diurnal temperature range in the CMIP5 models. Clim. Dyn. 2015, 44, 405–421. [Google Scholar] [CrossRef]
- Rohde, R.; Muller, R.; Jacobsen, R.; Muller, E.; Perlmutter, S.; Rosenfeld, A.; Wurtele, J.; Groom, D.; Wickham, C. A New estimate of the average earth surface land temperature spanning 1753 to 2011. Geoinf. Geostat. Overv. 2013, 7, 2. [Google Scholar]
- Perez, C.; Nicklin, C.; Dangles, O.; Vanek, S.; Sherwood, S.; Halloy, S.; Garrett, K.A.; Forbes, G. Climate change in the high Andes: Implications and adaptation strategies for small-scale farmers. Int. J. Environ. Cult. Econ. Soc. Sustain. 2010, 6, 71–88. [Google Scholar] [CrossRef]
- Scherm, H.; Van Bruggen, A. Effects of fluctuating temperatures on the latent period of lettuce downy mildew (Bremia lactucae). Phytopathology 1994, 84, 853–859. [Google Scholar] [CrossRef]
- Scherm, H.; Van Bruggen, A. Weather variables associated with infection of lettuce by downy mildew (Bremia lactucae) in coastal California. Phytopathology 1994. [Google Scholar] [CrossRef]
- Shakya, S.; Goss, E.; Dufault, N.; van Bruggen, A. Potential effects of diurnal temperature oscillations on potato late blight with special reference to climate change. Phytopathology 2015, 105, 230–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, H.; Van Bruggen, A.; Subbarao, K. Spore release of Bremia lactucae on lettuce is affected by timing of light initiation and decrease in relative humidity. Phytopathology 2000, 90, 67–71. [Google Scholar] [CrossRef] [Green Version]
- Wu, B.; Subbarao, K.; Van Bruggen, A. Factors affecting the survival of Bremia lactucae sporangia deposited on lettuce leaves. Phytopathology 2000, 90, 827–833. [Google Scholar] [CrossRef] [Green Version]
- Scherm, H.; Van Bruggen, A. Global warming and nonlinear growth: How important are changes in average temperature? Phytopathology 1994, 84, 1380–1384. [Google Scholar] [CrossRef]
- van Bruggen, A.H.; Jones, J.W.; Fernandes, J.M.C.; Garrett, K.; Boote, K.J. Crop diseases and climate change in the AgMIP framework. In Handbook of Climate Change and Agroecosystems; The Agricultural Model Intercomparison and Improvement Project (AgMIP) Integrated Crop and Economic AssessmentsICP Series on Climate Change Impacts Adaptation and Mitigation; Imperial College Press: London, UK, 2015. [Google Scholar]
- Chiyaka, C.; Singer, B.H.; Halbert, S.E.; Morris, J.G.; van Bruggen, A.H. Modeling huanglongbing transmission within a citrus tree. Proc. Natl. Acad. Sci. USA 2012, 109, 12213–12218. [Google Scholar] [CrossRef] [Green Version]
- Olanya, O.; Starr, G.; Honeycutt, C.; Griffin, T.; Lambert, D. Microclimate and potential for late blight development in irrigated potato. Crop Prot. 2007, 26, 1412–1421. [Google Scholar] [CrossRef]
- Andrade-Piedra, J.L.; Forbes, G.A.; Shtienberg, D.; Grünwald, N.J.; Chacón, M.G.; Taipe, M.V.; Hijmans, R.J.; Fry, W.E. Qualification of a plant disease simulation model: Performance of the LATEBLIGHT model across a broad range of environments. Phytopathology 2005, 95, 1412–1422. [Google Scholar] [CrossRef] [PubMed]
- Harrison, J.; Lowe, R. Effects of humidity and air speed on sporulation of Phytophthora infestans on potato leaves. Plant Pathol. 1989, 38, 585–591. [Google Scholar] [CrossRef]
- Savary, S.; Stetkiewicz, S.; Brun, F.; Willocquet, L. Modelling and mapping potential epidemics of wheat diseases—Examples on leaf rust and Septoria tritici blotch using EPIWHEAT. Eur. J. Plant Pathol. 2015, 142, 771–790. [Google Scholar] [CrossRef]
- Su, H.; Van Bruggen, A.; Subbarao, K.; Scherm, H. Sporulation of Bremia lactucae affected by temperature, relative humidity, and wind in controlled conditions. Phytopathology 2004, 94, 396–401. [Google Scholar] [CrossRef] [Green Version]
- Sunseri, M.A.; Johnson, D.A.; Dasgupta, N. Survival of detached sporangia of Phytophthora infestans exposed to ambient, relatively dry atmospheric conditions. Am. J. Potato Res. 2002, 79, 443. [Google Scholar] [CrossRef]
- Yuen, J.E. Modelling pathogen competition and displacement—Phytophthora infestans in Scandinavia. Eur. J. Plant Pathol. 2012, 133, 25–32. [Google Scholar] [CrossRef]
- Knappenberger, P.C.; Michaels, P.J.; Schwartzman, P.D. Observed changes in the diurnal temperature and dewpoint cycles across the United States. Geophys. Res. Lett. 1996, 23, 2637–2640. [Google Scholar] [CrossRef]
- Gordon, R.; Brown, D.; Dixon, M. Estimating potato leaf area index for specific cultivars. Potato Res. 1997, 40, 251–266. [Google Scholar] [CrossRef]
- Sparks, A.H.; Forbes, G.A.; Hijmans, R.; Garrett, K.A. A metamodeling framework for extending the application domain of process-based ecological models. Ecosphere 2011, 2, 1–14. [Google Scholar] [CrossRef]
- Schoolfield, R.M.; Sharpe, P.; Magnuson, C.E. Non-linear regression of biological temperature-dependent rate models based on absolute reaction-rate theory. J. Theor. Biol. 1981, 88, 719–731. [Google Scholar] [CrossRef]
- Sall, M.A. Epidemiology of grape powdery mildew: A model. Phytopathology 1980, 70, 338–342. [Google Scholar] [CrossRef]
- WeatherUnderground. Available online: http://www.wunderground.com (accessed on 1 March 2020).
- White, J.W.; Rassweiler, A.; Samhouri, J.F.; Stier, A.C.; White, C. Ecologists should not use statistical.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Temperature a | ±0 °C b | ±5 °C | ±10 °C | ||||||
|---|---|---|---|---|---|---|---|---|---|
| L0c | R2 | Slope | L0 | R2 | Slope | L0 | R2 | Slope | |
| 10 | 0.0078 | 0.934 | 1.02 | 0.025 | 0.970 | 1.01 | 0.0088 | 0.997 | 1.00 |
| 12 | 0.0312 | 0.991 | 1.02 | 0.056 | 0.996 | 1.02 | 0.0122 | 0.995 | 0.999 |
| 15 | 0.034 | 0.994 | 1.03 | 0.0280 | 0.997 | 1.02 | 0.0101 | 0.997 | 1.01 |
| 17 | 0.0215 | 0.996 | 1.03 | 0.019 | 0.996 | 1.06 | 0.0158 | 0.997 | 1.02 |
| 20 | 0.0122 | 0.995 | 1.04 | 0.0055 | 0.996 | 1.02 | 0.030 | 0.995 | 1.03 |
| 23 | 0.0280 | 0.966 | 1.10 | 0.006 | 0.997 | 1.04 | 0.037 | 0.988 | 1.04 |
| 27 | 0.0195 | 0.994 | 1.04 | 0.0036 | 0.998 | 1.02 | 0.016 | 0.989 | 1.03 |
| Location | Year | Cultivar | RRR | L0 | R2 | |
|---|---|---|---|---|---|---|
| Cutuglahua | 1997 | Bolona | 0.052 | 0.0007 | 0.977 | |
| Cutuglahua | 1997 | Gabriela | 0.049 | 0.0007 | 0.976 | |
| Cutuglahua | 1998 | Gabriela | 0.049 | 0.0001 | 0.991 | |
| La Tola | 1997 | Bolona | 0.052 | 0.000001 | 0.961 | |
| La Tola | 1997 | Gabriela | 0.049 | 0.000001 | 0.875 | |
| La Tola | 1998 | Gabriela | 0.049 | 0.000001 | 0.897 |
| State Variables | Description (Units) | Initial Values |
|---|---|---|
| H | Healthy susceptible sites | Table 1 |
| Lx | Latently infected sites | |
| I | Infectious sites | 0 |
| R | Removed sites | 0 |
| Y | Sum of infectious and removed sites | 0 |
| Driving variables | ||
| T | Hourly temperature (°C) | 0-37 |
| RH | Hourly relative humidity (%) | 60-95 |
| Parameters | ||
| LPR0 | Latency progression rate 1 (h−1) | 1/53 at 23 °C |
| LPR1 | Latency progression rate 2 (h−1) | 1/24 at 23 °C |
| RLGR1 | Relative lesion growth rate 1(h−1) | 0.9544 |
| RLGR2 | Relative lesion growth rate 2(h−1) | 0. 07336 |
| RLGR3 | Relative lesion growth rate 3(h−1) | 0. 03888 |
| RLGR4 | Relative lesion growth rate 4(h−1) | 0.02648 |
| RLGR5 | Relative lesion growth rate 5(h−1) | 0.02009 |
| REMRATE | Relative rate of removal (h−1) | 1/24 |
| HSP | Relative hourly spore production (h−1 ) | 45 |
| DILFAC | Dilution factor (-) | 0.01 |
| RRR ( = HSP*DILFAC) | Relative reproduction rate (h−1) | 0 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Narouei-Khandan, H.A.; Shakya, S.K.; Garrett, K.A.; Goss, E.M.; Dufault, N.S.; Andrade-Piedra, J.L.; Asseng, S.; Wallach, D.; Bruggen, A.H.C.v. BLIGHTSIM: A New Potato Late Blight Model Simulating the Response of Phytophthora infestans to Diurnal Temperature and Humidity Fluctuations in Relation to Climate Change. Pathogens 2020, 9, 659. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9080659
Narouei-Khandan HA, Shakya SK, Garrett KA, Goss EM, Dufault NS, Andrade-Piedra JL, Asseng S, Wallach D, Bruggen AHCv. BLIGHTSIM: A New Potato Late Blight Model Simulating the Response of Phytophthora infestans to Diurnal Temperature and Humidity Fluctuations in Relation to Climate Change. Pathogens. 2020; 9(8):659. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9080659
Chicago/Turabian StyleNarouei-Khandan, Hossein A., Shankar K. Shakya, Karen A. Garrett, Erica M. Goss, Nicholas S. Dufault, Jorge L. Andrade-Piedra, Senthold Asseng, Daniel Wallach, and Ariena H.C van Bruggen. 2020. "BLIGHTSIM: A New Potato Late Blight Model Simulating the Response of Phytophthora infestans to Diurnal Temperature and Humidity Fluctuations in Relation to Climate Change" Pathogens 9, no. 8: 659. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9080659