
Metabolic Potential of Epichloë Endophytes for Host Grass Fungal Disease Resistance

 and
and
Abstract
:1. Introduction
2. Epichloë Endophytes
3. Novel Endophytes
4. Disease Stress to Pasture Grasses
5. Epichloë sp.-Derived Antifungal Activity and Host Disease Resistance Studies
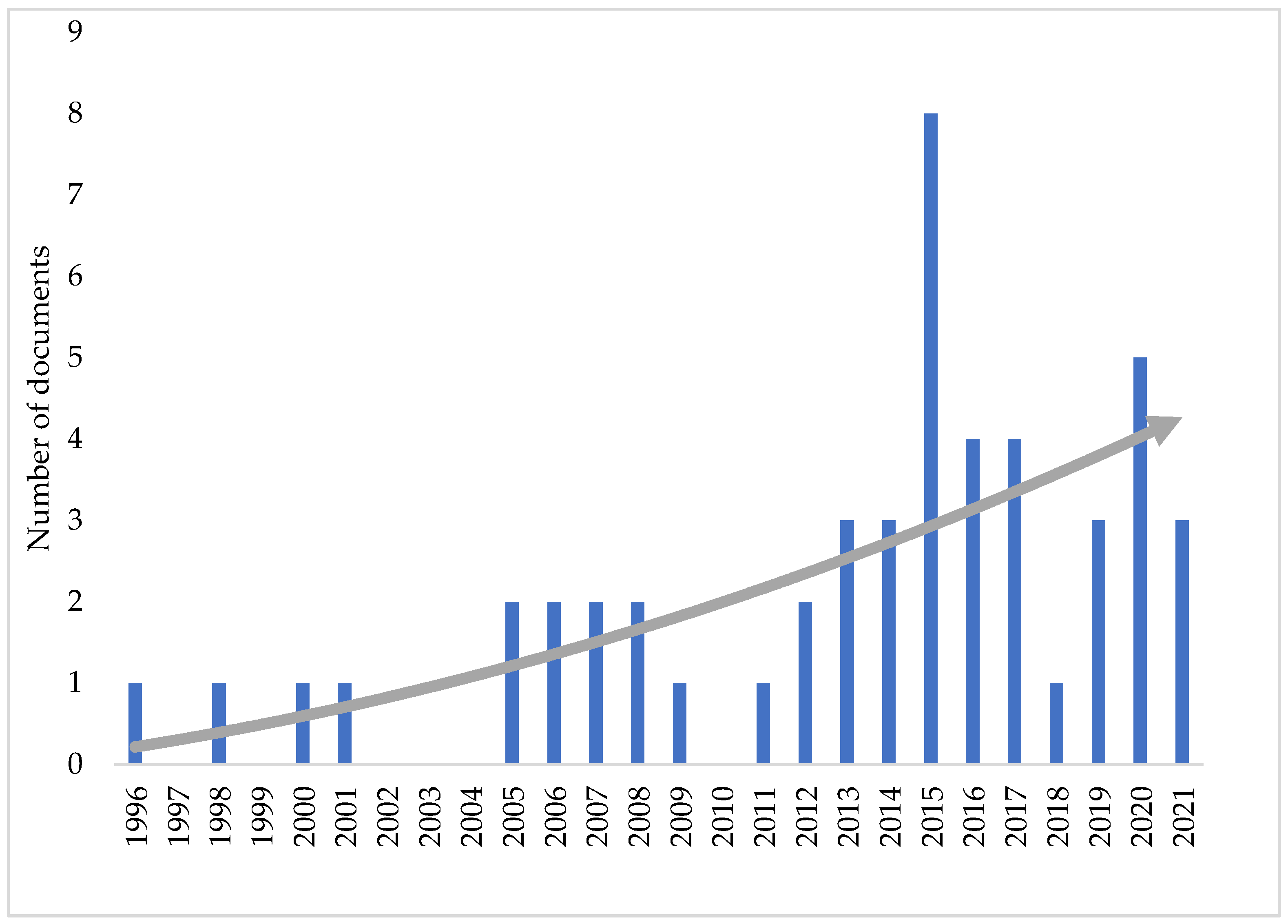
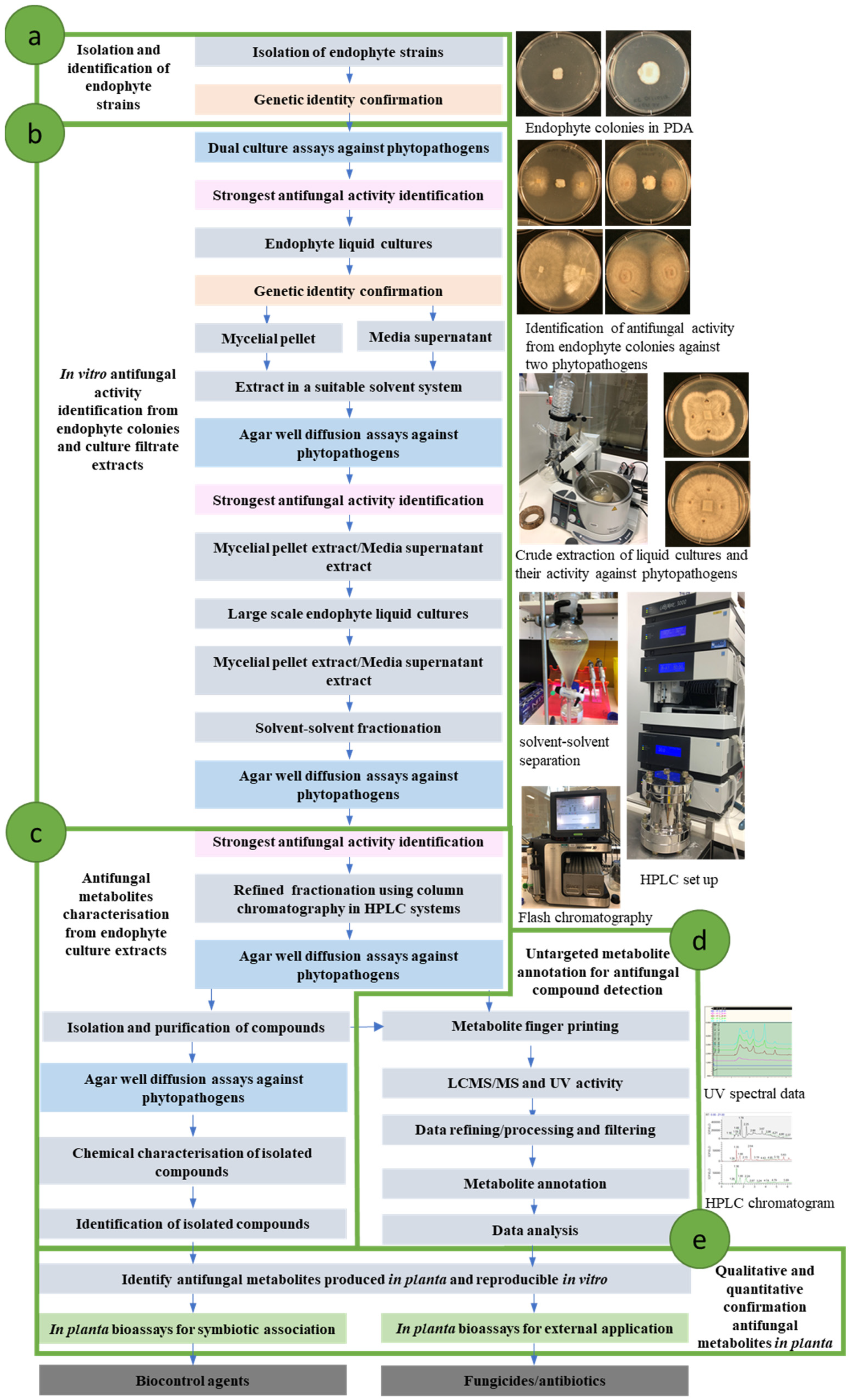
6. Bioprospecting Antifungal Metabolites from Epichloë Endophyte Strains
6.1. Epichloë Endophyte Strain Identification
6.2. Identification of Bioactive Strains Using In Vitro and in Planta Assays
6.3. Antifungal Metabolite Isolation and Characterisation
6.4. Untargeted Metabolite Annotation for Antifungal Compound Detection
6.5. Qualitative and Quantitative Confirmation of Antifungal Metabolites in Planta
7. Future Directions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tauer, L.W.; Mishra, A.K. Can the small dairy farm remain competitive in US agriculture? Food Policy 2006, 31, 458–468. [Google Scholar] [CrossRef] [Green Version]
- Alonso, M.E.; González-Montaña, J.R.; Lomillos, J.M. Consumers’ concerns and perceptions of farm animal welfare. Animals 2020, 10, 385. [Google Scholar] [CrossRef] [Green Version]
- FAO; IFAD; UNICEF; WFP; WHO. The State of Food Security and Nutrition in the World 2021; FAO: Rome, Italy, 2021. [Google Scholar]
- Vågsholm, I.; Arzoomand, N.S.; Boqvist, S. Food security, safety, and sustainabilitygetting the trade-offs right. Front. Sustain. Food Syst. 2020, 4, 16. [Google Scholar] [CrossRef]
- Charlton, J.; Stewart, A. Pasture species and cultivars used in New Zealand-a list. Proc. Conf. N. Z. Grassl. Assoc. 1999, 61, 147–166. [Google Scholar] [CrossRef]
- Reed, K. Species and Cultivars for Improved Pastures; Department of Primary Industries: Victoria, Australia, 2006; pp. 20–33.
- Jones, R.A. Virus diseases of pasture grasses in Australia: Incidences, losses, epidemiology, and management. Crop Pas. Sci. 2013, 64, 216–233. [Google Scholar] [CrossRef]
- Suzuki, N.; Rivero, R.M.; Shulaev, V.; Blumwald, E.; Mittler, R. Abiotic and biotic stress combinations. New Phytol. 2014, 203, 32–43. [Google Scholar] [CrossRef]
- Singh, L.P.; Gill, S.S.; Tuteja, N. Unraveling the role of fungal symbionts in plant abiotic stress tolerance. Plant Signal Behav. 2011, 6, 175–191. [Google Scholar] [CrossRef] [Green Version]
- Karpyn Esqueda, M.; Yen, A.L.; Rochfort, S.; Guthridge, K.M.; Powell, K.S.; Edwards, J.; Spangenberg, G.C. A review of perennial ryegrass endophytes and their potential use in the management of African black beetle in perennial grazing systems in Australia. Front. Plant Sci. 2017, 8, 3. [Google Scholar] [CrossRef] [Green Version]
- Xia, C.; Li, N.; Zhang, Y.; Li, C.; Zhang, X.; Nan, Z. Role of Epichloë endophytes in defense responses of cool-season grasses to pathogens: A review. Plant Dis. 2018, 102, 2061–2073. [Google Scholar] [CrossRef] [Green Version]
- Card, S.D.; Bastías, D.A.; Caradus, J.R. Antagonism to plant pathogens by Epichloë fungal endophytes—A review. Plants 2021, 10, 1997. [Google Scholar] [CrossRef]
- Scott, B. Epichloë endophytes: Fungal symbionts of grasses. Curr. Opin. Microbiol. 2001, 4, 393–398. [Google Scholar] [CrossRef]
- Young, C.A.; Tapper, B.A.; May, K.; Moon, C.D.; Schardl, C.L.; Scott, B. Indole-diterpene biosynthetic capability of Epichloë endophytes as predicted by ltm gene analysis. Appl. Environ. Microbiol. 2009, 75, 2200–2211. [Google Scholar] [CrossRef] [Green Version]
- Guerre, P. Ergot alkaloids produced by endophytic fungi of the genus Epichloë. Toxins 2015, 7, 773–790. [Google Scholar] [CrossRef] [Green Version]
- Leuchtmann, A. Taxonomy and diversity of Epichloë endophytes. In Clavicipitalean Fungi; CRC Press: Boca Raton, FL, USA, 2003; pp. 175–197. [Google Scholar]
- Kirby, E.J.M. Host-parasite relations in the choke disease of grasses. Trans. Br. Mycol. Soc. 1961, 44, 493–503. [Google Scholar] [CrossRef]
- Schardl, C.L.; Leuchtmann, A.; Spiering, M.J. Symbioses of grasses with seedborne fungal endophytes. Annu. Rev. Plant Biol. 2004, 55, 315–340. [Google Scholar] [CrossRef]
- Reddy, P.; Hemsworth, J.; Guthridge, K.M.; Vinh, A.; Vassiliadis, S.; Ezernieks, V.; Spangenberg, G.C.; Rochfort, S.J. Ergot alkaloid mycotoxins: Physiological effects, metabolism and distribution of the residual toxin in mice. Sci. Rep. 2020, 10, 9714. [Google Scholar] [CrossRef] [PubMed]
- Reddy, P.; Guthridge, K.; Vassiliadis, S.; Hemsworth, J.; Hettiarachchige, I.; Spangenberg, G.; Rochfort, S. Tremorgenic mycotoxins: Structure diversity and biological activity. Toxins 2019, 11, 302. [Google Scholar] [CrossRef] [Green Version]
- Tor-Agbidye, J. Correlation of endophyte toxins (ergovaline and lolitrem B) with clinical disease: Fescue foot and perennial ryegrass staggers. Vet. Hum. Tox. 1993, 43, 140–146. [Google Scholar]
- Fernando, K.; Reddy, P.; Vassiliadis, S.; Spangenberg, G.C.; Rochfort, S.J.; Guthridge, K.M. The known antimammalian and insecticidal alkaloids are not responsible for the antifungal activity of Epichloë endophytes. Plants 2021, 10, 2486. [Google Scholar] [CrossRef] [PubMed]
- Leuchtmann, A.; Schmidt, D.; Bush, L.P. Different levels of protective alkaloids in grasses with stroma-forming and seed-transmitted Epichloë/Neotyphodium endophytes. J. Chem. Ecol. 2000, 26, 1025–1036. [Google Scholar] [CrossRef]
- Schardl, C.L.; Grossman, R.B.; Nagabhyru, P.; Faulkner, J.R.; Mallik, U.P. Loline alkaloids: Currencies of mutualism. Phytochemistry 2007, 68, 980–996. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Zhang, X.; Li, C.; Christensen, M.J.; Nan, Z. Antifungal activity and phytochemical investigation of the asexual endophyte of Epichloë sp. from Festuca sinensis. Sci. China Life Sci. 2015, 58, 821–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ekanayake, P.N.; Kaur, J.; Tian, P.; Rochfort, S.J.; Guthridge, K.M.; Sawbridge, T.I.; Spangenberg, G.C.; Forster, J.W. Genomic and metabolic characterisation of alkaloid biosynthesis by asexual Epichloë fungal endophytes of tall fescue pasture grasses. Genome 2017, 60, 496–509. [Google Scholar] [CrossRef] [PubMed]
- Kaur, J.; Ekanayake, P.N.; Tian, P.; van Zijll de Jong, E.; Dobrowolski, M.P.; Rochfort, S.J.; Mann, R.C.; Smith, K.F.; Forster, J.W.; Guthridge, K.M.; et al. Discovery and characterisation of novel asexual Epichloë endophytes from perennial ryegrass (Lolium perenne L.). Crop Past. Sci. 2015, 66, 1058. [Google Scholar] [CrossRef]
- Hettiarachchige, I.; Ekanayake, P.; Mann, R.; Guthridge, K.; Spangenberg, G.; Forster, J. Phylogenomics of asexual Epichloë fungal endophytes forming associations with perennial ryegrass. BMC Evolut. Biol. 2015, 15, 72. [Google Scholar] [CrossRef] [Green Version]
- Caradus, J.; Chapman, D.; Cookson, T.; Cotching, B.; Deighton, M.; Donnelly, L.; Ferguson, J.; Finch, S.; Gard, S.; Hume, D. Epichloë endophytes–new perspectives on a key ingredient for resilient perennial grass pastures. NZGA Res. Pract. Ser. 2021, 17, 57–70. [Google Scholar] [CrossRef]
- Johnson, L.J.; Caradus, J.R. The science required to deliver Epichloë endophytes to commerce. In Endophytes for a Growing World; Hodkinson, T.R., Doohan, F.M., Saunders, M.J., Murphy, B.R., Eds.; Cambridge University Press: Cambridge, UK, 2019; pp. 343–370. [Google Scholar]
- Schardl, C.L.; Balestrini, R.; Florea, S.; Zhang, D.; Scott, B. Epichloë endophytes: Clavicipitaceous symbionts of grasses. In Plant Relationships; Springer: Berlin/Heidelberg, Germany, 2009; pp. 275–306. [Google Scholar]
- Ekanayake, P.; Rabinovich, M.; Guthridge, K.; Spangenberg, G.; Forster, J. Phylogenomics of fescue grass-derived fungal endophytes based on selected nuclear genes and the mitochondrial gene complement. BMC Evol. Biol. 2013, 13, 270. [Google Scholar] [CrossRef] [Green Version]
- Eady, C. The impact of alkaloid-producing Epichloë endophyte on forage ryegrass breeding: A New Zealand perspective. Toxins 2021, 13, 158. [Google Scholar] [CrossRef]
- Vassiliadis, S.; Elkins, A.C.; Reddy, P.; Guthridge, K.M.; Spangenberg, G.C.; Rochfort, S.J. A simple LC–MS method for the quantitation of alkaloids in endophyte-infected perennial ryegrass. Toxins 2019, 11, 649. [Google Scholar] [CrossRef] [Green Version]
- Reddy, P.; Deseo, M.A.; Ezernieks, V.; Guthridge, K.; Spangenberg, G.; Rochfort, S. Toxic indole diterpenes from endophyte-infected perennial ryegrass Lolium perenne L.: Isolation and stability. Toxins 2019, 11, 16. [Google Scholar] [CrossRef] [Green Version]
- Fernando, K.; Reddy, P.; Guthridge, K.M.; Spangenberg, G.C.; Rochfort, S.J. A Metabolomic Study of Epichloë Endophytes for Screening Antifungal Metabolites. Metabolites 2022, 12, 37. [Google Scholar] [CrossRef]
- Fernando, K.; Reddy, P.; Hettiarachchige, I.K.; Spangenberg, G.C.; Rochfort, S.J.; Guthridge, K.M. Novel antifungal activity of Lolium-associated Epichloë endophytes. Microorganisms 2020, 8, 955. [Google Scholar] [CrossRef]
- Wishart, D.S. Computational strategies for metabolite identification in metabolomics. Bioanalysis 2009, 1, 1579–1596. [Google Scholar] [CrossRef] [PubMed]
- Price, T. Ryegrass rust in Victoria. Plant Prot. Q. 1987, 2, 189. [Google Scholar]
- Mattner, S.; Parbery, D. Crown rust affects plant performance and interference ability of Italian ryegrass in the post-epidemic generation. Grass Forage Sci. 2007, 62, 437–444. [Google Scholar] [CrossRef]
- Potter, L. Effect of crown rust on regrowth, competitive ability and nutritional quality of perennial and Italian ryegrasses. Plant Pathol. 1987, 36, 455–461. [Google Scholar] [CrossRef]
- Plummer, R.; Hall, R.; Watt, T. The influence of crown rust (Puccinia coronata) on tiller production and survival of perennial ryegrass (Lolium perenne) plants in simulated swards. Grass Forage Sci. 1990, 45, 9–16. [Google Scholar] [CrossRef]
- Clarke, R. Rusts in Ryegrass. Available online: https://www.vgls.vic.gov.au/client/en_AU/search/asset/1281530/0 (accessed on 14 November 2021).
- Clarke, R.; Eagling, D. Effects of pathogens on perennial pasture grasses. N. Z. J. Agric. Res. 1994, 37, 319–327. [Google Scholar] [CrossRef] [Green Version]
- Aldaoud, R.; Anderson, M.; Reed, K.; Smith, K. Evidence of pathotypes among Australian isolates of crown rust infecting perennial ryegrass. Plant Breed. 2004, 123, 395–397. [Google Scholar] [CrossRef]
- Lancashire, J.A.; Latch, G.C.M. Some effects of crown rust (Puccinia coronata Corda) on the growth of two ryegrass varieties in New Zealand. N. Z. J. Agric. Res. 1966, 9, 628–640. [Google Scholar] [CrossRef]
- Di Menna, M.E.; Smith, B.L.; Miles, C.O. A history of facial eczema (pithomycotoxicosis) research. N. Z. J. Agric. Res. 2009, 52, 345–376. [Google Scholar] [CrossRef]
- Lam, A.; Lewis, G.C. Effects of nitrogen and potassium fertilizer application on Drechslera spp. and Puccinia coronata on perennial ryegrass (Lolium perenne) foliage. Plant Pathol. 1982, 31, 123–131. [Google Scholar] [CrossRef]
- Labruyere, R. Contamination of ryegrass seed with Drechslera species and its effect on disease incidence in the ensuing crop. Net. J. Plant Pathol. 1977, 83, 205–215. [Google Scholar] [CrossRef]
- Viji, G.; Wu, B.; Kang, S.; Uddin, W.; Huff, D. Pyricularia grisea causing gray leaf spot of perennial ryegrass turf: Population structure and host specificity. Plant Dis. 2001, 85, 817–826. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, N.; Hsiang, T. Snow Mold: The Battle under Snow between Fungal Pathogens and Their Plant Hosts; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Kolliker, R.; Kraehenbuehl, R.; Boller, B.; Widmer, F. Genetic diversity and pathogenicity of the grass pathogen Xanthomonas translucens pv. graminis. Syst. Appl. Microb. 2006, 29, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Bender, C.L.; Young, S.A.; Mitchell, R.E. Conservation of plasmid DNA sequences in coronatine-producing pathovars of Pseudomonas syringae. Appl. Environ. Microb. 1991, 57, 993–999. [Google Scholar] [CrossRef] [Green Version]
- Farquhar, M.; Winefield, C.; Eady, C. Dry matter yield and the prevalence of barley yellow dwarf and ryegrass mosaic viruses in old and young perennial ryegrass. J. N. Z. Grassl. 2017, 79, 165–171. [Google Scholar] [CrossRef]
- Fink-Gremmels, J. Mycotoxins in cattle feeds and carry-over to dairy milk: A review. Food Addit. Contam. 2008, 25, 172–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitlow, L.W.; Hagler, W. Mycotoxins in dairy cattle: Occurrence, toxicity, prevention and treatment. In Proceedings of the Southwest Nutrition Conference, Tempe, AZ, USA, 24–25 February 2005; pp. 124–138. [Google Scholar]
- Read, E.; Edwards, J.; Deseo, M.; Rawlin, G.; Rochfort, S. Current understanding of acute bovine liver disease in Australia. Toxins 2017, 9, 8. [Google Scholar] [CrossRef]
- Aslani, M.R.; Pascoe, I.; Kowalski, M.; Michalewicz, A.; Retallick, M.; Colegate, S. In vitro detection of hepatocytotoxic metabolites from Drechslera biseptata: A contributing factor to acute bovine liver disease? Aust. J. Exp. Agric. 2006, 46, 599–604. [Google Scholar] [CrossRef]
- Smith, K.; Simpson, R.; Oram, R.; Lowe, K.; Kelly, K.; Evans, P.; Humphreys, M. Seasonal variation in the herbage yield and nutritive value of perennial ryegrass (Lolium perenne L.) cultivars with high or normal herbage water-soluble carbohydrate concentrations grown in three contrasting Australian dairy environments. Aust. J. Exp. Agric. 1998, 38, 821–830. [Google Scholar] [CrossRef]
- Latch, G. Fungous diseases of ryegrasses in New Zealand: I. Foliage diseases. N. Z. J. Agric. Res. 1966, 9, 394–409. [Google Scholar] [CrossRef]
- Buckingham, S.; McCalman, H.; Powell, H. Pest and Disease Control in Grass and Forage Crops; Farming Connect: Milton Park, UK, 2013.
- Lucas, J. Advances in plant disease and pest management. J. Agric. Sci. 2011, 149, 91–114. [Google Scholar] [CrossRef]
- Uddin, W.; Viji, G.; Vincelli, P. Gray leaf spot (blast) of perennial ryegrass turf: An emerging problem for the turfgrass industry. Plant Dis. 2003, 87, 880–889. [Google Scholar] [CrossRef] [Green Version]
- Chynoweth, R.; Rolston, M.; Kelly, M.; Grbavac, N. Control of blind seed disease (Gloeotinia temulenta) in perennial ryegrass (Lolium perenne) seed crops and implications for endophyte transmission. Agron. N. Z. 2012, 42, 141–148. [Google Scholar]
- Hampton, J. Effect of nitrogen rate and time of application on seed yield in perennial ryegrass cv. grasslands Nui. N. Z. J. Exp. Agric. 1987, 15, 9–16. [Google Scholar] [CrossRef]
- Oruc, H.H. Fungicides and their effects on animals. In Fungicides; Carisse, O., Ed.; InTech: Rijeka, Croatia, 2010; pp. 349–362. [Google Scholar]
- Gupta, U.C.; Gupta, S.C. Trace element toxicity relationships to Crop. production and livestock and human health: Implications for management. Commun. Soil Sci. Plant Anal. 1998, 29, 1491–1522. [Google Scholar] [CrossRef]
- Harvey, I.; Fletcher, L.; Emms, L. Effects of several fungicides on the Lolium endophyte in ryegrass plants, seeds, and in culture. N. Z. J. Agric. Res. 1982, 25, 601–606. [Google Scholar] [CrossRef] [Green Version]
- Viji, G.; Uddin, W.; Romaine, C. Suppression of gray leaf spot (blast) of perennial ryegrass turf by Pseudomonas aeruginosa from spent mushroom substrate. Biol. Control 2003, 26, 233–243. [Google Scholar] [CrossRef]
- Kim, D.S.; Rae, C.Y.; Chun, S.J.; Kim, D.H.; Choi, S.W.; Choi, K.H. Paenibacillus elgii SD17 as a biocontrol agent against soil-borne turf diseases. Plant Pathol. J. 2005, 21, 328–333. [Google Scholar] [CrossRef] [Green Version]
- De Silva, N.I.; Brooks, S.; Lumyong, S.; Hyde, K.D. Use of endophytes as biocontrol agents. Fungal Biol. Rev. 2019, 33, 133–148. [Google Scholar] [CrossRef]
- Zhang, X.; Xia, C.; Li, C.; Nan, Z. Chemical composition and antifungal activity of the volatile oil from Epichloë gansuensis, endophyte-infected and non-infected Achnatherum inebrians. Sci. China Life Sci. 2015, 58, 512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, K.A.; Berry, D.; Feussner, K.; Eaton, C.J.; Ram, A.; Mesarich, C.H.; Solomon, P.; Feussner, I.; Scott, B. Lolium perenne apoplast metabolomics for identification of novel metabolites produced by the symbiotic fungus Epichloë festucae. New Phytol. 2020, 227, 559–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertrand, S.; Schumpp, O.; Bohni, N.; Bujard, A.; Azzollini, A.; Monod, M.; Gindro, K.; Wolfender, J.-L. Detection of metabolite induction in fungal co-cultures on solid media by high-throughput differential ultra-high pressure liquid chromatography–time-of-flight mass spectrometry fingerprinting. J. Chromatogr. A 2013, 1292, 219–228. [Google Scholar] [CrossRef]
- Ito, T.; Odake, T.; Katoh, H.; Yamaguchi, Y.; Aoki, M. High-throughput profiling of microbial extracts. J. Nat. Prod. 2011, 74, 983–988. [Google Scholar] [CrossRef]
- Reheul, D.; Ghesquiere, A. Breeding perennial ryegrass with better crown rust resistance. Plant Breed. 1996, 115, 465–469. [Google Scholar] [CrossRef]
- Bonos, S.A.; Kubik, C.; Clarke, B.B.; Meyer, W.A. Breeding perennial ryegrass for resistance to gray leaf spot. Crop Sci. 2004, 44, 575–580. [Google Scholar] [CrossRef]
- Han, Y.; Bonos, S.A.; Clarke, B.B.; Meyer, W.A. Inheritance of resistance to gray leaf spot disease in perennial ryegrass. Crop Sci. 2006, 46, 1143–1148. [Google Scholar] [CrossRef]
- Vargas, J.M. Management of Turfgrass Diseases, 2nd ed.; CRC Press: Boca Raton, FL, USA, 1994. [Google Scholar]
- Pfender, W. Interaction of fungicide physical modes of action and plant phenology in control of stem rust of perennial ryegrass grown for seed. Plant Dis. 2006, 90, 1225–1232. [Google Scholar] [CrossRef]
- Wilson, M.; Noble, M.; Gray, E.G. The blind seed disease of ryegrass and its causal fungus. Trans. R. Soc. Edinb. 1945, 61, 327–340. [Google Scholar] [CrossRef]
- Hofgaard, I.; Ergon, Å.; Wanner, L.; Tronsmo, A.M. The effect of chitosan and Bion on resistance to pink snow mould in perennial ryegrass and winter wheat. J. PhytoPathol. 2005, 153, 108–119. [Google Scholar] [CrossRef]
- Sneh, B.; Jabaji-Hare, S.; Neate, S.; Dijst, G. Rhizoctonia Species: Taxonomy, Molecular Biology, Ecology, Pathology and Disease Control; Springer Science & Business Media: Berlin/Heidelberg, Germany, 1996. [Google Scholar]
- Burpee, L. Rhizoctonia cerealis causes yellow patch of turfgrasses. Plant Dis. 1980, 64, 1114–1116. [Google Scholar] [CrossRef]
- Caesar, A.J. Identity, pathogenicity, and comparative virulence of Fusarium spp. related to stand declines of leafy spurge (Euphorbia esula) in the Northern Plains. Plant Dis. 1996, 80, 1395–1398. [Google Scholar] [CrossRef] [Green Version]
- Caesar, A.J.; Campobasso, G.; Terragitti, G. Effects of European and US strains of Fusarium spp. pathogenic to leafy spurge on North American grasses and cultivated species. Biol. Control 1999, 15, 130–136. [Google Scholar] [CrossRef] [Green Version]
- Egli, T.; Goto, M.; Schmidt, D. Bacterial wilt, a new forage grass disease. J. Phytopathol. 1975, 82, 111–121. [Google Scholar] [CrossRef]
- Schmidt, D.; Nuesch, B. Resistance to bacterial wilt (Xanthomonas graminis) increases yield and persistency of Lolium multiflorum. EPPO Bull. 1980, 10, 335–339. [Google Scholar] [CrossRef]
- Schmidt, D. Prevention of bacterial wilt of grasses by phylloplane bacteria. J. Phytopathol. 1988, 122, 253–260. [Google Scholar] [CrossRef]
- Jones, R. Virus diseases of Australian pastures. In Pasture and Forage Crop Pathology; Sukumar, C., Ed.; American Society of Agronomy, Crop Science Society of America, and Soil Science Society of America: Madison, WI, USA, 1996; pp. 303–322. [Google Scholar]
- Li, F.; Guo, Y.e.; Christensen, M.J.; Gao, P.; Li, Y.; Duan, T. An arbuscular mycorrhizal fungus and Epichloë festucae var. lolii reduce Bipolaris sorokiniana disease incidence and improve perennial ryegrass growth. Mycorrhiza 2018, 28, 159–169. [Google Scholar] [CrossRef]
- Christensen, M. Antifungal activity in grasses infected with Acremonium and Epichloë endophytes. Australas. Plant Pathol. 1996, 25, 186–191. [Google Scholar] [CrossRef]
- Christensen, M.J.; Latch, G.C.M.; Tapper, B.A. Variation within isolates of Acremonium endophytes from perennial ryegrasses. Mycol. Res. 1991, 95, 918–923. [Google Scholar] [CrossRef]
- Siegel, M.R.; Latch, G.C. Expression of antifungal activity in agar culture by isolates of grass endophytes. Mycologia 1991, 83, 529–537. [Google Scholar] [CrossRef]
- Tian, P.; Nan, Z.; Li, C.; Spangenberg, G. Effect of the endophyte Neotyphodium lolii on susceptibility and host physiological response of perennial ryegrass to fungal pathogens. Eur. J. Plant Pathol. 2008, 122, 593–602. [Google Scholar] [CrossRef]
- He, C.; Holme, J.; Anthony, J. SNP genotyping: The KASP assay. Methods Mol. Biol. 2014, 1145, 75–86. [Google Scholar] [PubMed]
- Niones, J.T.; Takemoto, D. An isolate of Epichloë festucae, an endophytic fungus of temperate grasses, has growth inhibitory activity against selected grass pathogens. J. Gen. Plant Pathol. 2014, 80, 337–347. [Google Scholar] [CrossRef]
- Stromstedt, A.A.; Felth, J.; Bohlin, L. Bioassays in natural product research—strategies and methods in the search for anti-inflammatory and antimicrobial activity. Phytochem. Anal. 2014, 25, 13–28. [Google Scholar] [CrossRef] [PubMed]
- Oldenburg, K.R.; Vo, K.T.; Ruhland, B.; Schatz, P.J.; Yuan, Z. A dual culture assay for detection of antimicrobial activity. J. Biomol. Screen. 1996, 1, 123–130. [Google Scholar] [CrossRef] [Green Version]
- Li, C.J.; Gao, J.H.; Nan, Z.B. Interactions of Neotyphodium gansuense, Achnatherum inebrians, and plant-pathogenic fungi. Mycol. Res. 2007, 111, 1220–1227. [Google Scholar] [CrossRef]
- Christensen, M.J.; Latch, G.C.M. Variation among isolates of Acremonium endophytes (A. coenophialum and possibly A. typhinum) from tall fescue (Festuca arundinacea). Mycol. Res. 1991, 95, 1123–1126. [Google Scholar] [CrossRef]
- Steinebrunner, F.; Schiestl, F.P.; Leuchtmann, A. Ecological role of volatiles produced by Epichloë: Differences in antifungal toxicity. FEMS Microbiol. Ecol. 2008, 64, 307–316. [Google Scholar] [CrossRef] [Green Version]
- Clarke, B.B.; White, J.F., Jr.; Hurley, R.H.; Torres, M.S.; Sun, S.; Huff, D.R. Endophyte-mediated suppression of dollar spot disease in fine fescues. Plant Dis. 2006, 90, 994–998. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Luo, Y.; Tian, P.; Peng, H.; Feng, J. Preliminary evaluation of the disease resistance of Festuca sinensis infected by Epichloë sinensis. J. Phytopathol. 2021, 169, 623–629. [Google Scholar] [CrossRef]
- Tian, Z.; Wang, R.; Ambrose, K.V.; Clarke, B.B.; Belanger, F.C. The Epichloë festucae antifungal protein has activity against the plant pathogen Sclerotinia homoeocarpa, the causal agent of dollar spot disease. Sci. Rep. 2017, 7, 5643. [Google Scholar] [CrossRef] [Green Version]
- Wiewióra, B.; Żurek, G.; Żurek, M. Endophyte-mediated disease resistance in wild populations of perennial ryegrass (Lolium perenne). Fungal Ecol. 2015, 15, 1–8. [Google Scholar] [CrossRef]
- Liu, L.; Guo, C.; Lv, H.; Gu, L.; Li, C. Effects of the Epichloë gansuensis endophyte on the disease resistance of drunken horse grass to powdery mildew. Acta Prataculturae Sin. 2015, 24, 65–71. [Google Scholar] [CrossRef]
- Sabzalian, M.R.; Mirlohi, A.; Sharifnabi, B. Reaction to powdery mildew fungus, Blumeria graminis in endophyte-infected and endophyte-free tall and meadow fescues. Australas. Plant Pathol. 2012, 41, 565–572. [Google Scholar] [CrossRef]
- Bonos, S.A.; Wilson, M.M.; Meyer, W.A.; Reed Funk, C. Suppression of red thread in fine fescues through endophyte-mediated resistance. Appl. Turfgrass Sci. 2005, 2, 1–7. [Google Scholar] [CrossRef]
- Li, F.; Duan, T.; Li, Y. Effects of the fungal endophyte Epichloë festucae var. lolii on growth and physiological responses of perennial ryegrass cv. fairway to combined drought and pathogen stresses. Microorganisms 2020, 8, 1917. [Google Scholar] [CrossRef]
- White, J.F.; Cole, G.T. Endophyte-host associations in forage grasses. IV. The endophyte of Festuca versuta. Mycologia 1986, 78, 102–107. [Google Scholar] [CrossRef]
- Yue, Q.; Miller, C.J.; White, J.F.; Richardson, M.D. Isolation and characterization of fungal inhibitors from Epichloë festucae. J. Agric. Food Chem. 2000, 48, 4687–4692. [Google Scholar] [CrossRef]
- Holzmann-Wirth, A.; Dapprich, P.; Eierdanz, S.; Heerz, D.; Paul, V. Anti-fungal substances extracted from Neotyphodium endophytes. In Proceedings of the 3rd International Conference on Harmful and Beneficial Microorganisms in Grassland, Pasture and Turf, Paderborn, Germany, 26–29 September 2000; pp. 65–69. [Google Scholar]
- Yoshihara, T.; Togiya, S.; Koshino, H.; Sakamura, S.; Shimanuki, T.; Sato, T.; Tajimi, A. Three fungitoxic cyclopentanoid sesquiterpenes from stromata of Epichloë Typhina. Tetrahedron Lett. 1985, 26, 5551–5554. [Google Scholar] [CrossRef]
- Koshino, H.; Togiya, S.; Yoshihara, T.; Sakamura, S.; Shimanuki, T.; Sato, T.; Tajimi, A. Four fungitoxic C-18 hydroxy unsaturated fatty acids from stromata of Epichloë Typhina. Tetrahedron Lett. 1987, 28, 73–76. [Google Scholar] [CrossRef]
- Purev, E.; Kondo, T.; Takemoto, D.; Niones, J.T.; Ojika, M. Identification of ε-poly-l-lysine as an antimicrobial product from an Epichloë endophyte and isolation of fungal ε-PL synthetase gene. Molecules 2020, 25, 1032. [Google Scholar] [CrossRef] [Green Version]
- Kusari, S.; Hertweck, C.; Spiteller, M. Chemical ecology of endophytic fungi: Origins of secondary metabolites. Chem. Biol. 2012, 19, 792–798. [Google Scholar] [CrossRef] [Green Version]
- Hettiarachchige, I.K.; Jagt, C.J.V.; Mann, R.C.; Sawbridge, T.I.; Spangenberg, G.C.; Guthridge, K.M. Global changes in asexual Epichloë transcriptomes during the early stages, from seed to seedling, of symbiotum establishment. Microorganisms 2021, 9, 991. [Google Scholar] [CrossRef]
- Winter, D.J.; Ganley, A.R.; Young, C.A.; Liachko, I.; Schardl, C.L.; Dupont, P.-Y.; Berry, D.; Ram, A.; Scott, B.; Cox, M.P. Repeat elements organise 3D genome structure and mediate transcription in the filamentous fungus Epichloë festucae. PLoS Genet. 2018, 14, e1007467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rochfort, S. Metabolomics reviewed: A new “omics” platform technology for systems biology and implications for natural products research. J. Nat. Prod. 2005, 68, 1813–1820. [Google Scholar] [CrossRef] [PubMed]
- Mashego, M.R.; Rumbold, K.; De Mey, M.; Vandamme, E.; Soetaert, W.; Heijnen, J.J. Microbial metabolomics: Past, present and future methodologies. Biotechnol. Lett. 2007, 29, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Pinu, F.; Villas-Boas, S.; Aggio, R. Analysis of intracellular metabolites from microorganisms: Quenching and extraction protocols. Metabolites 2017, 7, 53. [Google Scholar] [CrossRef] [Green Version]
- Pinu, F.; Villas-Boas, S. Extracellular microbial metabolomics: The state of the art. Metabolites 2017, 7, 43. [Google Scholar] [CrossRef]
- Behrends, V.; Ebbels, T.M.; Williams, H.D.; Bundy, J.G. Time-resolved metabolic footprinting for nonlinear modeling of bacterial substrate utilization. Appl. Environ. Microb. 2009, 75, 2453–2463. [Google Scholar] [CrossRef] [Green Version]
- Chumnanpuen, P.; Hansen, M.A.E.; Smedsgaard, J.; Nielsen, J. Dynamic metabolic footprinting reveals the key components of metabolic network in yeast Saccharomyces cerevisiae. Int. J. Genom. 2014, 2014, 894296. [Google Scholar] [CrossRef] [Green Version]
- Hajjaj, H.; Blanc, P.; Goma, G.; Francois, J. Sampling techniques and comparative extraction procedures for quantitative determination of intra-and extracellular metabolites in filamentous fungi. FEMS Microbiol. Lett. 1998, 164, 195–200. [Google Scholar] [CrossRef]
- Rasmussen, S.; Parsons, A.J.; Popay, A.; Xue, H.; Newman, J.A. Plant-endophyte-herbivore interactions: More than just alkaloids? Plant Signal Behav. 2008, 3, 974–977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasmussen, S.; Parsons, A.J.; Fraser, K.; Xue, H.; Newman, J.A. Metabolic profiles of Lolium perenne are differentially affected by nitrogen supply, carbohydrate content, and fungal endophyte infection. Plant Physiol. 2008, 146, 1440–1453. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, S.; Parsons, A.J.; Newman, J.A. Metabolomics analysis of the Lolium perenne–Neotyphodium lolii symbiosis: More than just alkaloids? Phytochem. Rev. 2009, 8, 535–550. [Google Scholar] [CrossRef] [Green Version]
- Patchett, A.; Newman, J.A. Comparison of plant metabolites in root exudates of Lolium perenne infected with different strains of the fungal endophyte Epichloë festucae var. lolii. J. Fungi 2021, 7, 148. [Google Scholar] [CrossRef] [PubMed]
- Geddes-McAlister, J.; Sukumaran, A.; Patchett, A.; Hager, H.A.; Dale, J.; Roloson, J.L.; Prudhomme, N.; Bolton, K.; Muselius, B.; Powers, J. Examining the impacts of CO2 concentration and genetic compatibility on perennial ryegrass—Epichloë festucae var. lolii interactions. J. Fungi 2020, 6, 360. [Google Scholar] [CrossRef]
- Hiroyuki, K.; Satoshi, T.; Shun-ichi, T.; Yoshihara, T.; Sakamura, S.; Shimanuki, T.; Sato, T.; Tajimi, A. New fungitoxic sesquiterpenoids, chokols A-G, from stromata of Epichloë typhina and the absolute configuration of chokol E. Agric. Biol. Chem. 1989, 53, 789–796. [Google Scholar] [CrossRef]
- Koshino, H.; Yoshihara, T.; Okuno, M.; Sakamura, S.; Tajimi, A.; Shimanuki, T. Gamahonolides A, B, and gamahorin, novel antifungal compounds from stromata of Epichloë typhina on Phleum pratense. Biosci. Biotech. Biochem. 1992, 56, 1096–1099. [Google Scholar] [CrossRef]
- Koshino, H.; Terada, S.-I.; Yoshihara, T.; Sakamura, S.; Shimanuki, T.; Sato, T.; Tajimi, A. Three phenolic acid derivatives from stromata of Epichloë typhina on Phleum pratense. Phytochemistry 1988, 27, 1333–1338. [Google Scholar] [CrossRef]
- Seto, Y.; Takahashi, K.; Matsuura, H.; Kogami, Y.; Yada, H.; Yoshihara, T.; Nabeta, K. Novel cyclic peptide, epichlicin, from the endophytic fungus, Epichloe Typhina. Biosci. Biotech. Biochem. 2007, 71, 1470–1475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Q.Y.; Nan, Z.B.; Gao, K.; Song, H.; Tian, P.; Zhang, X.X.; Li, C.J.; Xu, W.B.; Li, X.Z. Antifungal, phytotoxic, and cytotoxic activities of metabolites from Epichloë bromicola, a fungus obtained from Elymus tangutorum grass. J. Agric. Food Chem. 2015, 63, 8787–8792. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Country | PBR Grant Date | Endophyte Strain (Market Name) | Epichloë Species | Host Common Name | Known Alkaloid Profile 2 | Applicant |
|---|---|---|---|---|---|---|
| New Zealand | 23 April 1996 (expired) | AR1 | E. festucae var. lolii (LpTG-1) | Perennial ryegrass | P | Grasslanz Technology Ltd. (GTL; Palmerston North, New Zealand) |
| European Union | 20 October 2003 | |||||
| Australia | 26 October 2004 | |||||
| New Zealand | 21 April 2015 | AR1006 | E. uncinata | Meadow fescue | L | GTL |
| New Zealand | 5 October 2016 | AR1017 | E. uncinata | Meadow fescue | L | GTL |
| New Zealand | 29 November 2018 | AR127 | E. festucae var. lolii (LpTG-1) | Perennial ryegrass | GTL | |
| New Zealand | 25 July 2008 | AR37 | E. festucae var. lolii (LpTG-1) | Perennial ryegrass | J | GTL |
| Australia | 30 March 2010 | |||||
| Uruguay | 20 November 2011 | Fischer Fleurquin Gustavo (FFG; Montevideo, Uruguay) | ||||
| New Zealand | 23 April 1996 (expired) | AR501 | E. coenophiala (FaTG-1) | Tall fescue | LP | GTL |
| European Union | 20 October 2003 | |||||
| New Zealand | 1 February 1999 (expired) | AR542 (MaxP® New Zealand, Australia; MaxQ® USA) | E. coenophiala (FaTG-1) | Tall fescue | LP | GTL |
| Australia | 26 October 2004 | |||||
| Uruguay | 6 December 2004 | FFG | ||||
| New Zealand | 25 July 2008 | AR584 (MaxP® New Zealand, Australia; MaxQII™ USA) | E. coenophiala (FaTG-1) | Tall fescue | LP | GTL |
| Australia | 29 September 2010 | |||||
| Uruguay | 2 August 2013 | FFG | ||||
| Argentina | 4 September 2014 | Gentos SA (Buenos Aires, Argentina) | ||||
| New Zealand | 12 May 2010 | AR601 (Avanex®) | E. coenophiala (FaTG-1) | Tall fescue | EL | GTL |
| Australia | 19 August 2013 | |||||
| European Union | 22 May 2017 | |||||
| New Zealand | 12 May 2010 | AR604 | E. coenophiala (FaTG-1) | Tall fescue | EL | GTL |
| European Union | 22 May 2017 | |||||
| New Zealand | 28 August 2014 | AR95 (Avanex®) | E. festucae var. lolii (LpTG-1) | Perennial ryegrass | E | GTL |
| European Union | 22 May 2017 | |||||
| Australia | 3 April 2018 | |||||
| Uruguay | 21 November 2014 | FFG | ||||
| New Zealand | 17 January 2019 | CM142 | E. festucae var. lolii (LpTG-1) | Perennial ryegrass | J | Cropmark Seeds Ltd. (CSL; Rolleston, New Zealand) |
| Australia | 17 August 2020 | Cropmark Seeds Australia Pty Ltd. (CSA; South Melbourne, Australia) | ||||
| New Zealand | 27 August 2014 | E815 (Edge) | E. festucae var. lolii (LpTG-1) | Perennial ryegrass | LtmEP | DLF Seeds A/S (DLF; Roskilde, Denmark) |
| Australia | 23 October 2017 | DLF | ||||
| New Zealand | 23 June 2010 | Happe | E. siegelii | L | DLF | |
| Australia | 23 October 2017 | DLF | ||||
| New Zealand | 25 July 2008 | Nea 2 (NEA/NEA2/NEA4) 3 | E. festucae var. lolii (LpTG-1) | Perennial ryegrass | LtmEP | Barenbrug New Zealand Ltd. (BBNZ; Christchurch, New Zealand) |
| New Zealand | 30 June 2009 | Nea 3 (NEA4 3) | E. festucae var. lolii (LpTG-1) | Perennial ryegrass | EP | BBNZ |
| New Zealand | 25 July 2008 | Nea 6 (NEA2 3) | E. festucae var. lolii (LpTG-1) | Perennial ryegrass | EP | BBNZ |
| New Zealand | 29 August 2014 | Nea 10 | E. festucae var. lolii (LpTG-1) | Perennial ryegrass | EP | BBNZ |
| New Zealand | 13 August 2014 | Nea 11 | E. festucae var. lolii (LpTG-1) | Perennial ryegrass | EP | BBNZ |
| New Zealand | 19 March 2021 | Nea 12 | Epichloë sp. (LpTG-3) | Perennial ryegrass | J | Agriculture Victoria Services Pty Ltd. (Bundoora, Australia) |
| New Zealand | 29 August 2014 | Nea 21 | Epichloë sp. (FaTG-3) | Tall fescue | LP | BBNZ |
| New Zealand | 29 August 2014 | Nea 23 | Epichloë sp. (FaTG-3) | Tall fescue | LP | BBNZ |
| New Zealand | 10 July 2019 | Nea 47 (NEA2 3) | E. festucae var. lolii (LpTG-1) | Perennial ryegrass | EP | BBNZ |
| New Zealand | 18 August 2014 | PTK647 (Protek®) | E. coenophiala (FaTG-1) | Tall fescue | EL | DLF |
| Australia | 23 October 2017 | DLF | ||||
| New Zealand | 28 August 2014 | U12 | E. uncinata | Meadow fescue | L | CSL |
| Australia | 12 August 2021 | CSA | ||||
| New Zealand | 2 September 2016 | U13 | E. uncinata | Meadow fescue | L | CSL |
| New Zealand | 25 July 2008 | U2 (GrubOUT®) | E. uncinata | Meadow fescue | L | CSL |
| Australia | 30 January 2014 | CSA | ||||
| Argentina | 11 March 2015 | Gentos SA | ||||
| European Union | 22 May 2017 | CSL | ||||
| New Zealand | 14 October 2008 | UNC1 | E. uncinata | Meadow fescue | L | CSL |
| Australia | Not applicable 4 | AR5 (Endo5) | E. festucae var. lolii (LpTG-1) | Perennial ryegrass | EP | GTL |
| USA | Not applicable 4 | E34® | E. coenophiala (FaTG-1) | Tall fescue | LP | Barenbrug USA (Tangent, OR, USA) |
| USA | Not applicable 5 | KY31 (Kentucky31) | E. coenophiala (FaTG-1) | Tall fescue | EPL | Not applicable |
| worldwide | Not applicable 5 | SE (Standard endophyte) | E. festucae var. lolii (LpTG-1) | Perennial ryegrass | LtmEP | Not applicable |
| Disease/ Common Name | Causative Organism | Symptoms | Damage/Loss | Control Measures | References |
|---|---|---|---|---|---|
| Crown rust | Puccinia coronata | Reddish brown spores on leaf | Dry matter yield loss 30–40% and stock thrift | Fungicide; Judicious grazing management; Resistant varieties. | [41,45,76] |
| Grey leaf spot | Pyricularia grisea | Small water-soaked lesions on leaf blades gradually turning to dark necrotic spots and to grey spots | Up to 90% pasture loss | Fungicide; Controlled release of N fertiliser; Biocontrol, Resistant varieties. | [50,63,77,78] |
| Brown blight and net blotch | Drechslera sp. | Net lesions with small dark brown bars amphigenous lesions with dark brown margins, light brown center | Dry matter and herbage loss | Fungicide; Managed grazing before it spreads. | [49,60,79] |
| Stem end rust | Puccinia graminis | Reddish brown spores on sheath and stem | Seed yield loss, Dry matter loss | Fungicide; Judicious grazing management; Resistant varieties. | [49,80] |
| Blind seed disease | Gloeotinia temulenta | Fungal mycelia on seeds under microscopic observation | Seed yield loss, reduce seed germination 50–90% | Fungicide; Increased rate of N application. | [64,65,81] |
| Snow mould | Microdochium nivale | Dark brown lesions and pink sporodochia rows parallel to veins | Seedling damage leading to yield loss, seed loss and yield loss | Fungicide; Biological control; Compost application. | [51,60,82] |
| Yellow patch | Ceratobasidium cereale | Root pathogen | Yield loss | Fungicide | [83,84] |
| seedling pathogen and leaf spot | Fusarium solani | Wilting of seedlings and necrotic lesions in mature plants | Yield loss, dry matter loss | Fungicide | [85,86] |
| Bacterial wilt | Xanthomonas translucens | Water-soaked lesions and turning to bluish purple colour | Forage yield loss 20–40% | Biological control; Resistant varieties. | [52,87,88,89] |
| Ryegrass mosaic virus | RGMV | Light green-yellow streaky mosaic or brown necrosis on leaves | Dry matter yield loss 21–30% | Resistant varieties; Mixed pasture species. | [7,90] |
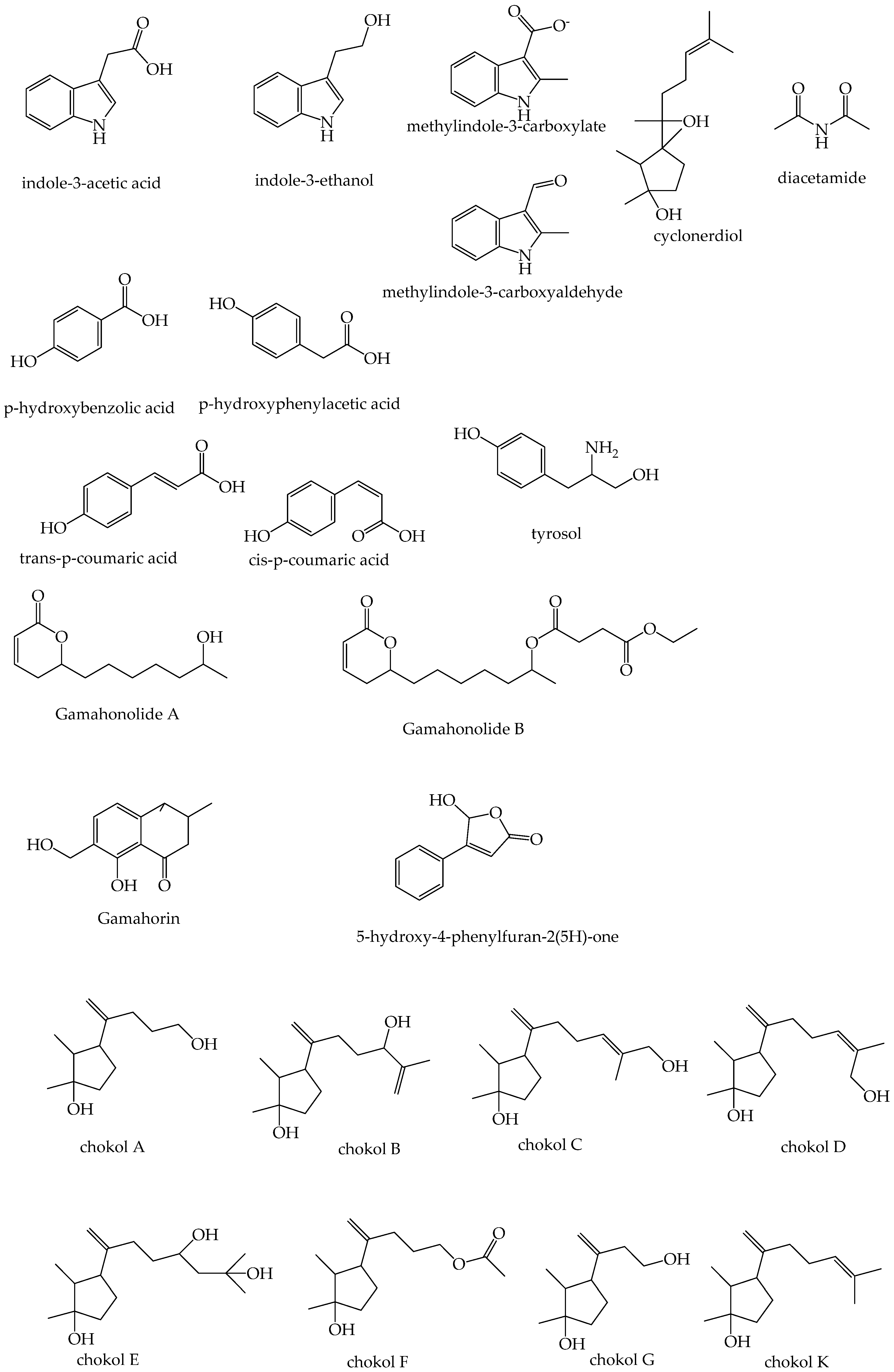
| Fungitoxic Compound | Chemical Characteristics | Chemical Formula | m/z or Molecular Weight | Epichloë sp. | Host Grass | Source Material | Tested Pathogens | Reference |
|---|---|---|---|---|---|---|---|---|
| Indole-3-acetic acid | IAA derivative | C10H9NO2 | 176.0667 | E. festucae | Festuca rubra | purified mycelia | Lactisaria fusiformis, Magnaporthe poae, Rhizoctonia solani | [112] |
| Indole-3-ethanol | IAA derivative | C10H11NO | 162.0874 | E. festucae | Festuca rubra | purified mycelia | L. fusiformis, M. poae, R. solani | [112] |
| Methylindole-3-carboxylate | IAA derivative | C10H9NO2 | 175.0594 | E. festucae | Festuca rubra | purified mycelia | L. fusiformis, M. poae, R. solani | [112] |
| Indole-3-carboxaldehyde | IAA derivative | C9H7NO | 159.0684 | E. festucae | Festuca rubra | purified mycelia | L. fusiformis, M. poae, R. solani | [112] |
| Cyclonerodiol | sesquiterpinoid | C15H28O2 | 165.0507 | E. festucae | Festuca rubra | purified mycelia | L. fusiformis, M. poae, R. solani | [112] |
| Chokol A Chokol B Chokol C Chokol D Chokol E Chokol F Chokol G | sesquiterpinoid | C12H22O2 C15H26O2 C15H26O2 C15H26O2 C15H28O3 C14H24O3 C11H20O2 | 199.1693 239.2006 239.2006 239.2006 271.2228 241.1798 185.1536 | E. typhina | Phleum pratense | Epichloë infected plant material (choke) | Cladosporium herbarum, Cladosporium phlei | [114,132] |
| Chokol K | sesquiterpinoid | C15H26O | 222.1984 1 [M-H2O]− = 204 | E. sylvatica E. clarkii | Brachypodium sylvaticum Holcus lanatus | Unfertilized stromata and unfertilized stromata head space | Stagonospora nodorum, Mycosphaerella graminicola | [102] |
| N,N-diacetamide | diactamide | C4H7NO2 | 102.0610 | E. festucae | Festuca rubra | Epichloë infected plant material (leaves) | L. fusiformis, M. poae, R. solani | [112] |
| Gamahonolide A Gamahonolide B Gamahorin | gamahonolide | C12H2 3 C18H28O6 C12H14O4 | 213.1502 341.1952 222.0881 | E. typhina | Phleum pratense | Epichloë infected plant material (choke) | [133] | |
| 5-hydroxy-4-phenyl-2(5H)-furanone | C12H10O3 | 177.0546 | E. typhina | Phleum pratense | Epichloë infected plant material (choke) | [133] | ||
| Trans-p-coumaric acid Cis-p-coumaric acid p-hydroxybenzoic acid p-hydroxyphenylacetic acid Tyrosol | phenolic acid derivatives | C9H8O3 C9H8O3 C7H6O3 C8H8O3 C8H10O2 | 164.1580 164.1580 139.1220 153.0507 168.0980 | E. typhina | Phleum pratense | Epichloë infected plant material (choke) | C. phlei | [134] |
| Epichlicin | Cyclic peptide | C48H74N12O14 | 3 [M+Na+17]+ = 1082 | E. typhina | Phleum pretense | purified mycelia | C. phlei | [135] |
| Fatty acids | C-18 and C-19 fatty acid | C18H32O3 C19H34O3 Cl9H36O3 Cl9H36O3 | 297 311 312 312 | E. typhina | Phleum pratense | Epichloë infected plant material (choke) | C. herbarum, C. phlei | [115] |
| Efe-AfpA | protein | 55 amino acids | 6278 Da | E. festucae | Festuca rubra subsp. rubra | Epichloë infected plant material (tiller) | Sclerotinia homoeocarpa | [105,136] |
| Cyclosporin T | peptides | C61H109N11O12 (11 amino acids) | 1188.6 g/mol (2 MW) | E. bromicola | Elymus tangutorum | purified mycelia | Alternaria alternata, Bipolaris sorokiniana, Fusarium avenaceum Curvularia lunata | [136] |
| ε-poly-L-lysines | peptides | 28–34 lysine sub-units | E. festucae | Festuca pulchella | purified mycelia | Drechslera erythrospila Phytophthora capsici | [116] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernando, K.; Reddy, P.; Spangenberg, G.C.; Rochfort, S.J.; Guthridge, K.M. Metabolic Potential of Epichloë Endophytes for Host Grass Fungal Disease Resistance. Microorganisms 2022, 10, 64. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10010064
Fernando K, Reddy P, Spangenberg GC, Rochfort SJ, Guthridge KM. Metabolic Potential of Epichloë Endophytes for Host Grass Fungal Disease Resistance. Microorganisms. 2022; 10(1):64. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10010064
Chicago/Turabian StyleFernando, Krishni, Priyanka Reddy, German C. Spangenberg, Simone J. Rochfort, and Kathryn M. Guthridge. 2022. "Metabolic Potential of Epichloë Endophytes for Host Grass Fungal Disease Resistance" Microorganisms 10, no. 1: 64. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10010064