
Comparative Surfaceome Analysis of Clonal Histomonas meleagridis Strains with Different Pathogenicity Reveals Strain-Dependent Profiles
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
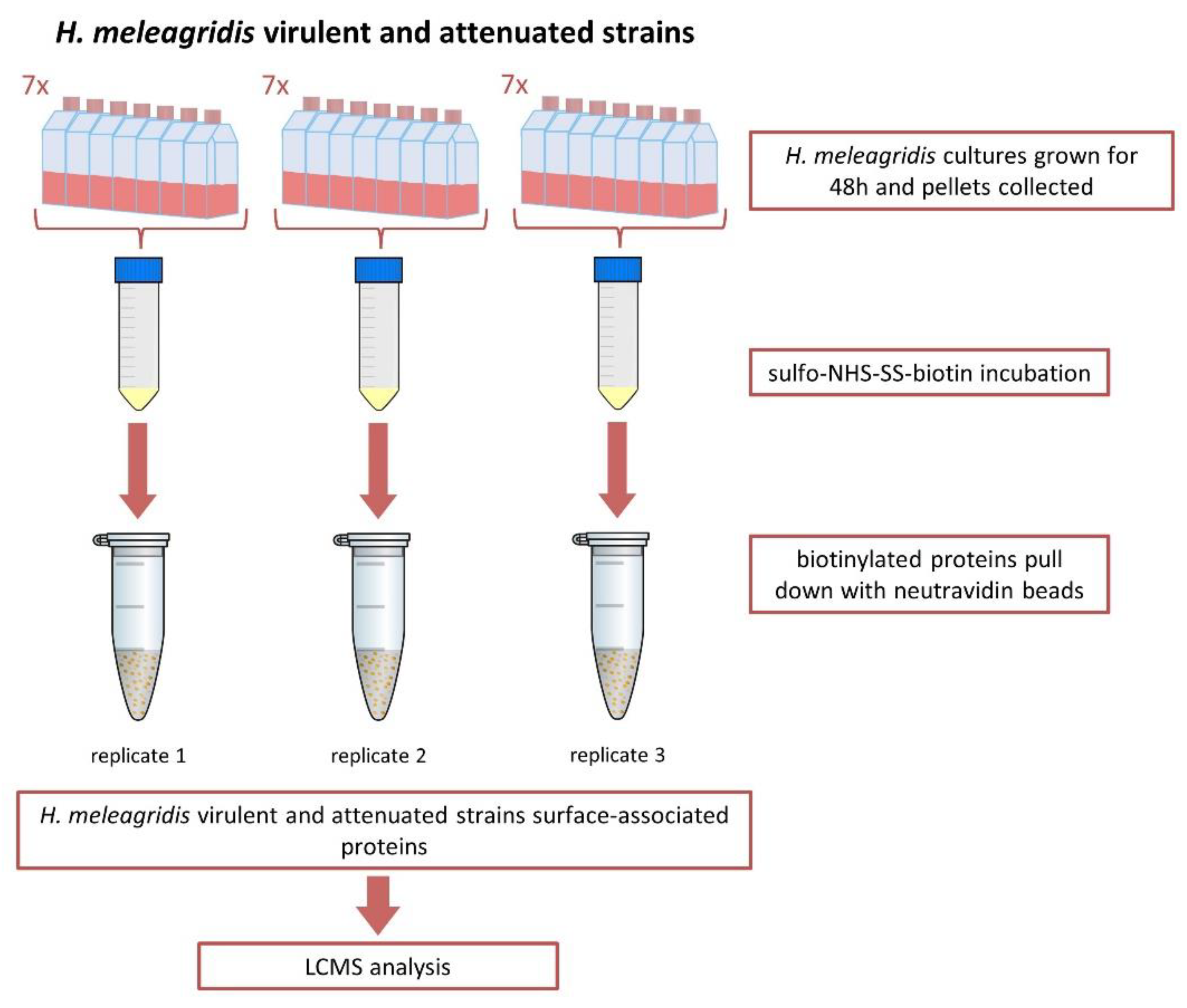
2.1. Protozoan Cultures
2.2. Biotinylation of Surface-Associated Proteins
2.3. Assessing Membrane Permeabilization after Biotinylation
2.4. Membrane Protein Enrichment and Purification of Biotinylated Proteins
2.5. One-Dimensional SDS-PAGE (Sodium Dodecyl Sulfate–Polyacrylamide Gel Electrophoresis)
2.6. Sample Preparation and nanoHPLC-Orbitrap MS/MS Analysis
2.7. H. meleagridis Proteome Database
2.8. Identification and Quantification of Surface-Associated Proteins
2.9. Re-Analysis of H. meleagridis Proteome and Exoproteome Data
2.10. In Silico Analysis
2.11. RNA Extraction and Quantitative Reverse Transcriptase Polymerase Chain Reaction (RT-qPCR) Analysis
3. Results
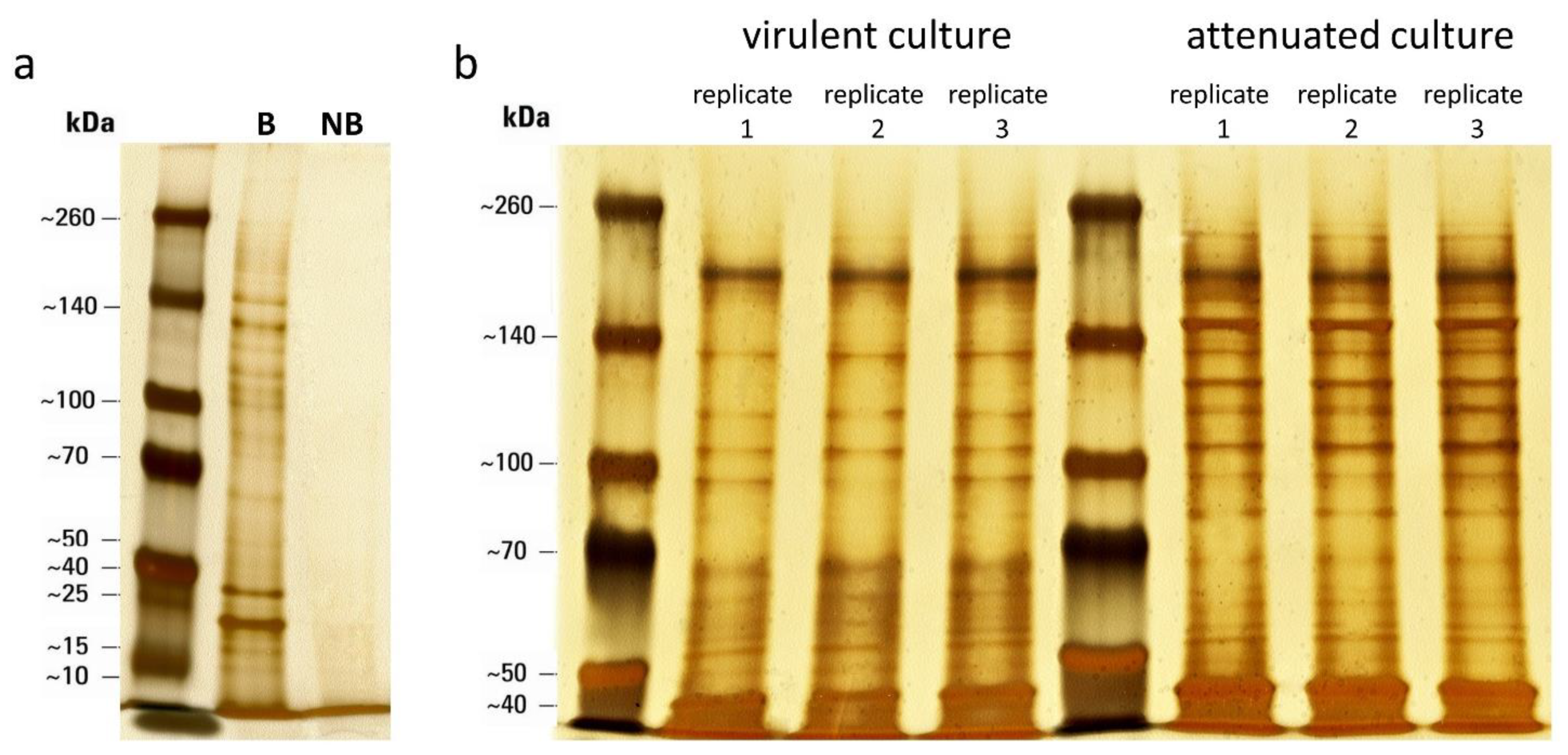
3.1. Selective Biotinylation of Surface-Associated Proteins
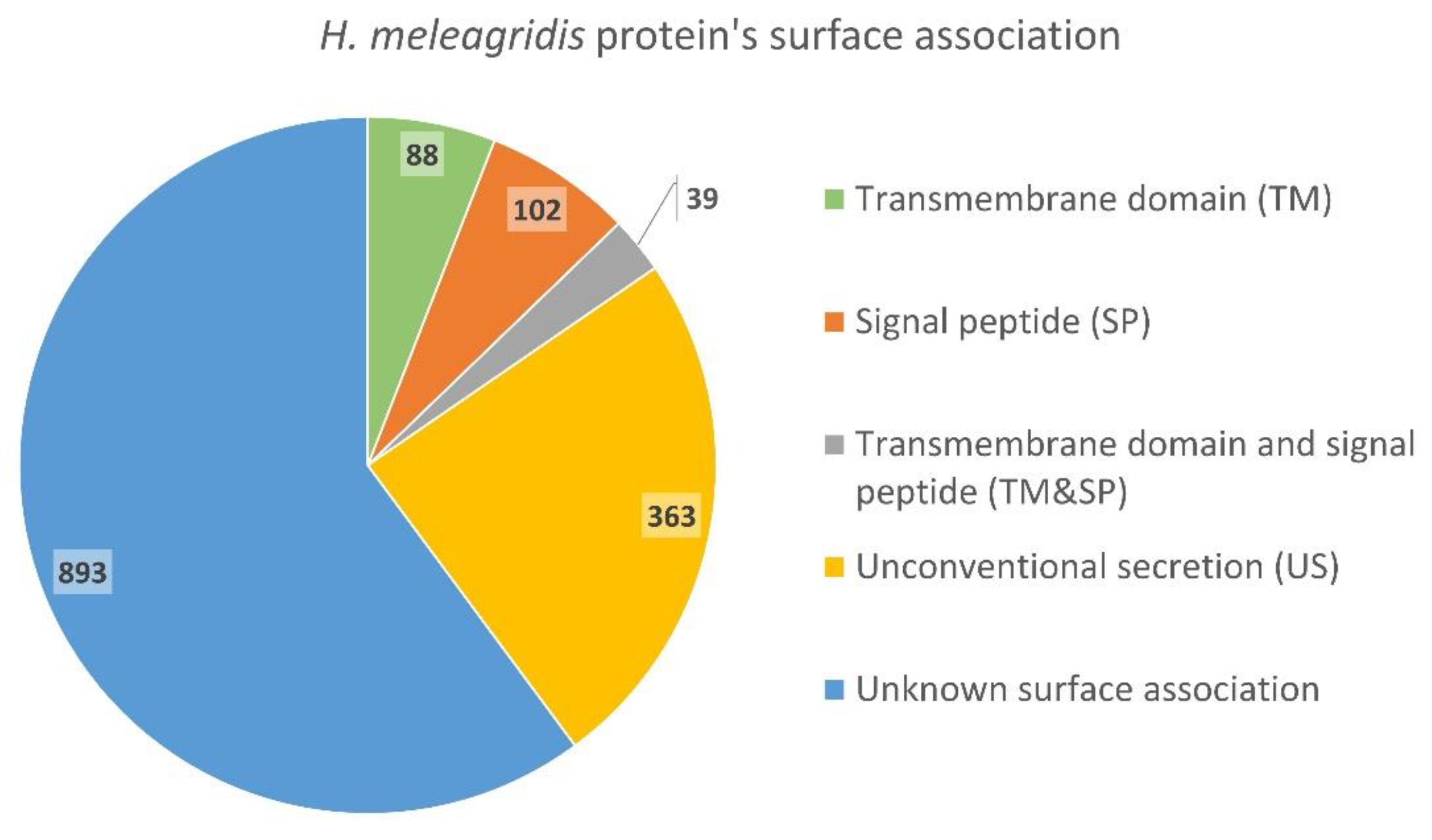
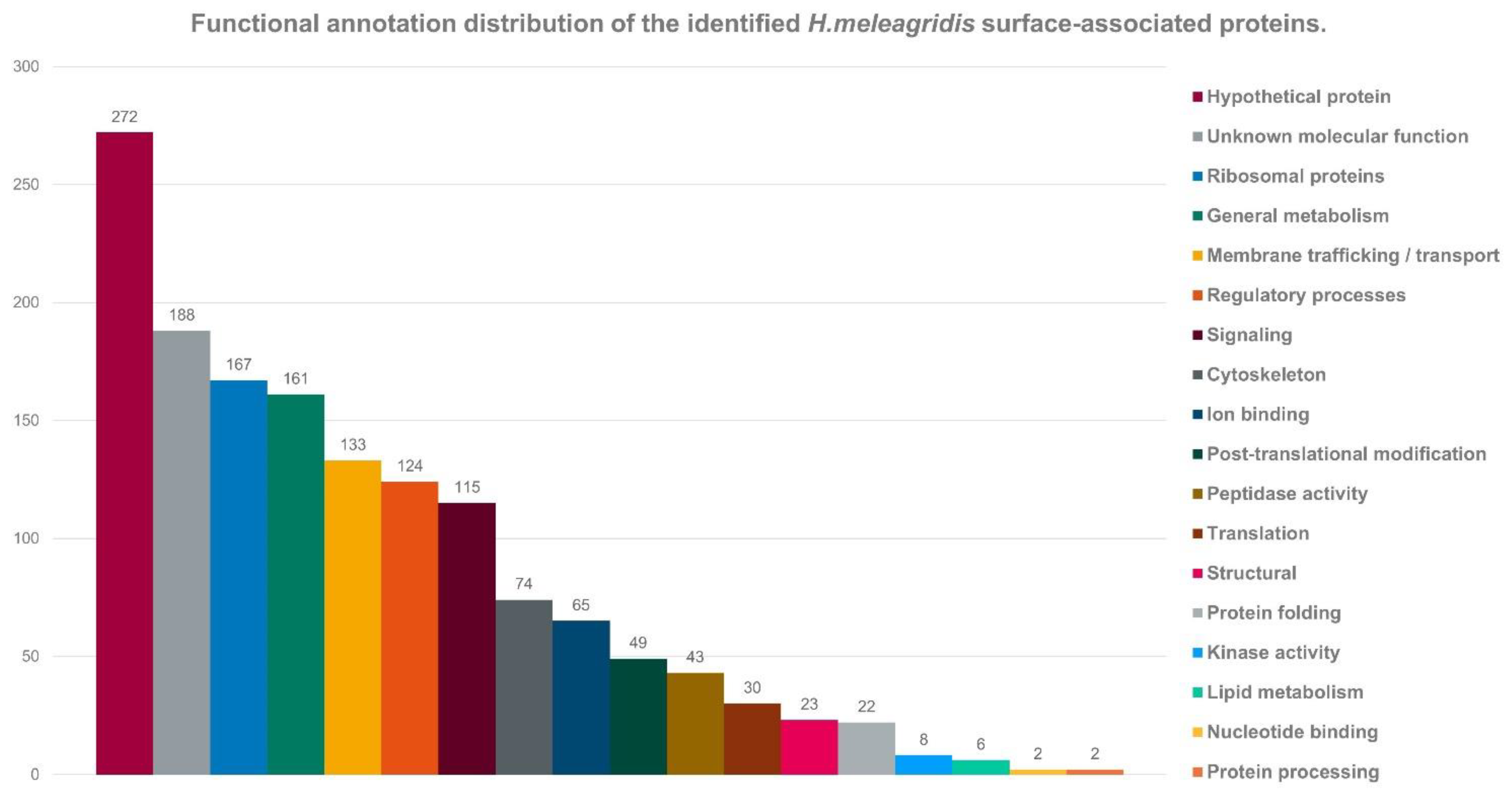
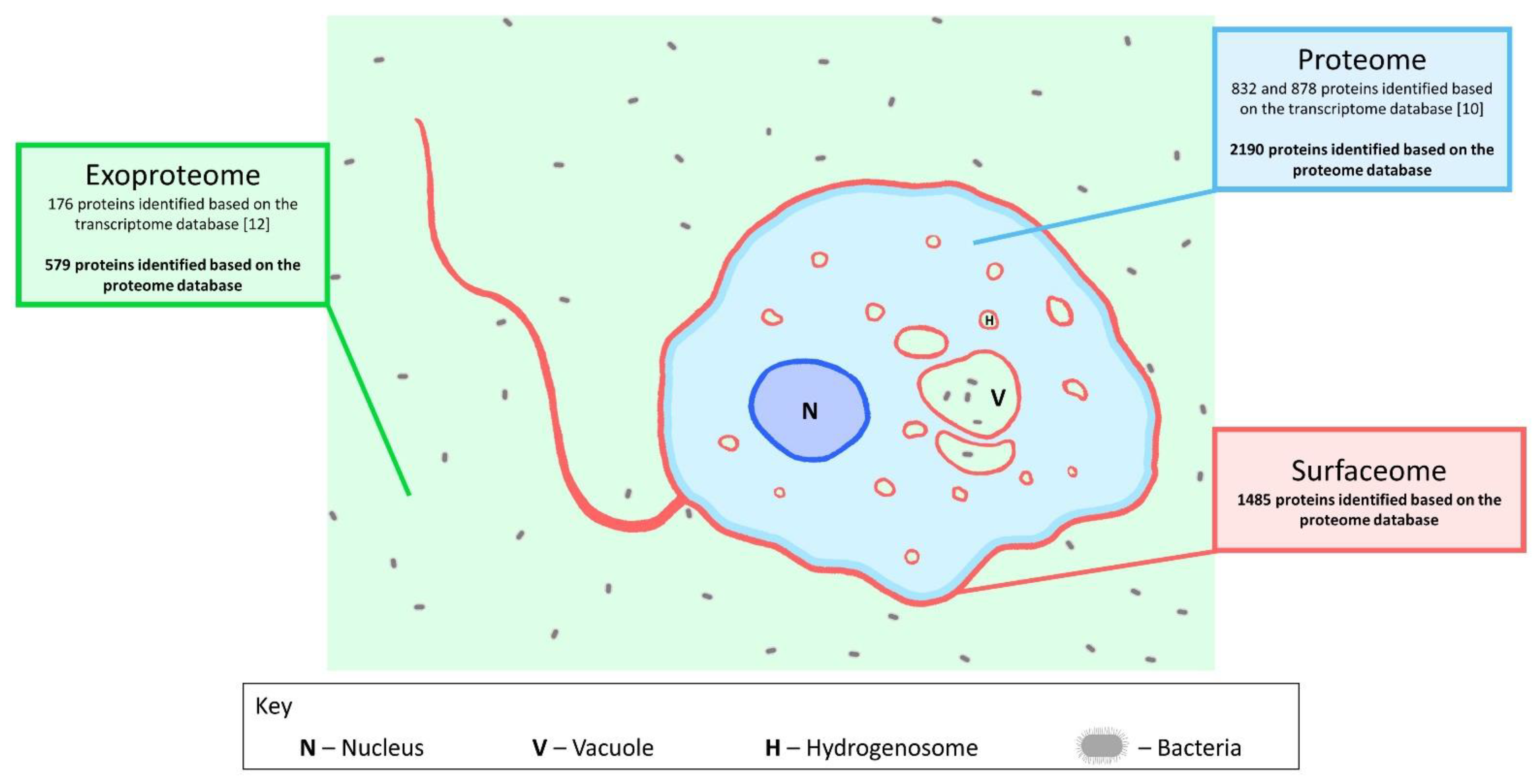
3.2. Identification and Quantification of Surface-Associated Proteins
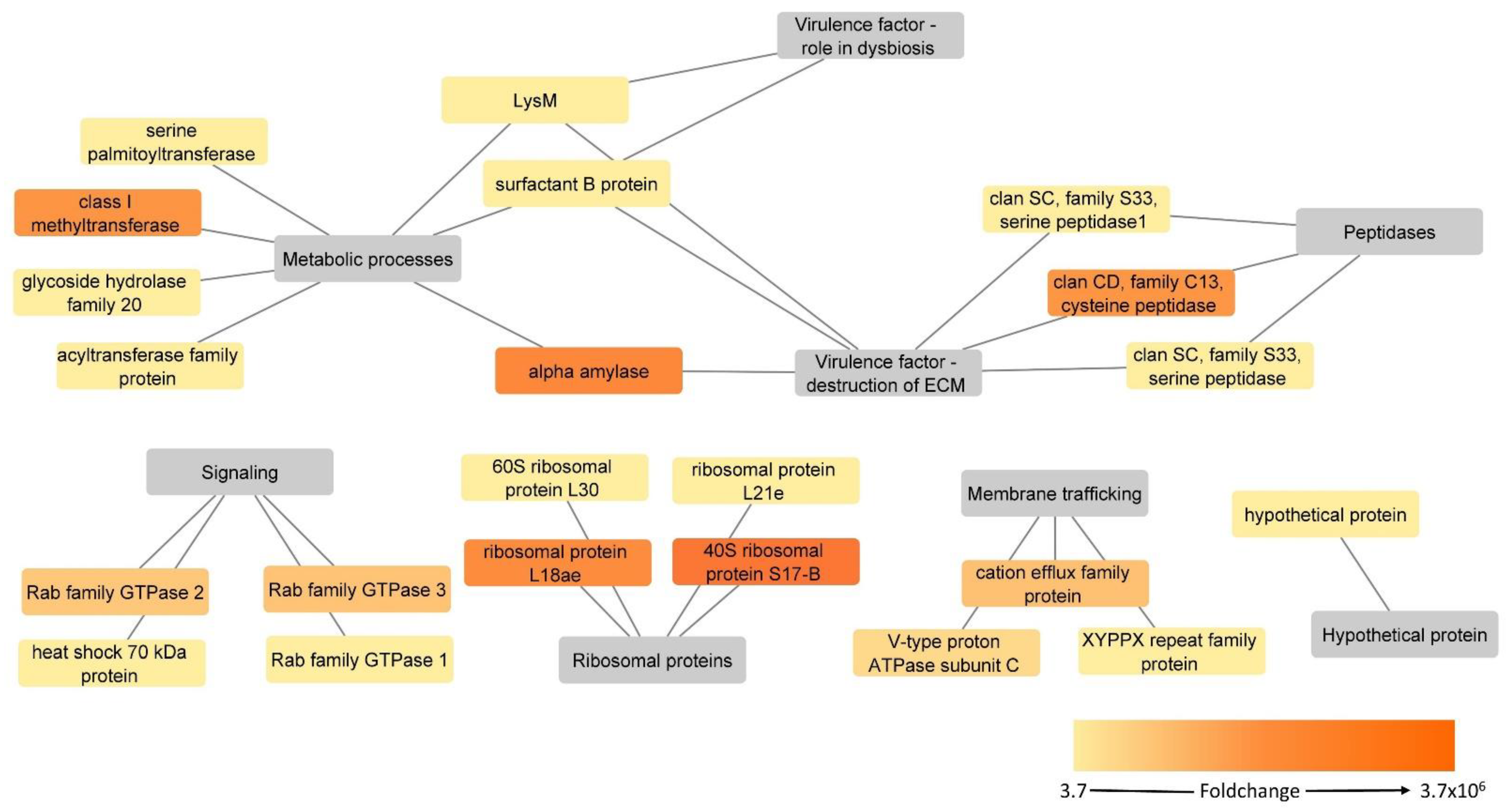
3.3. Proteins Upregulated in the H. meleagridis Virulent Strain
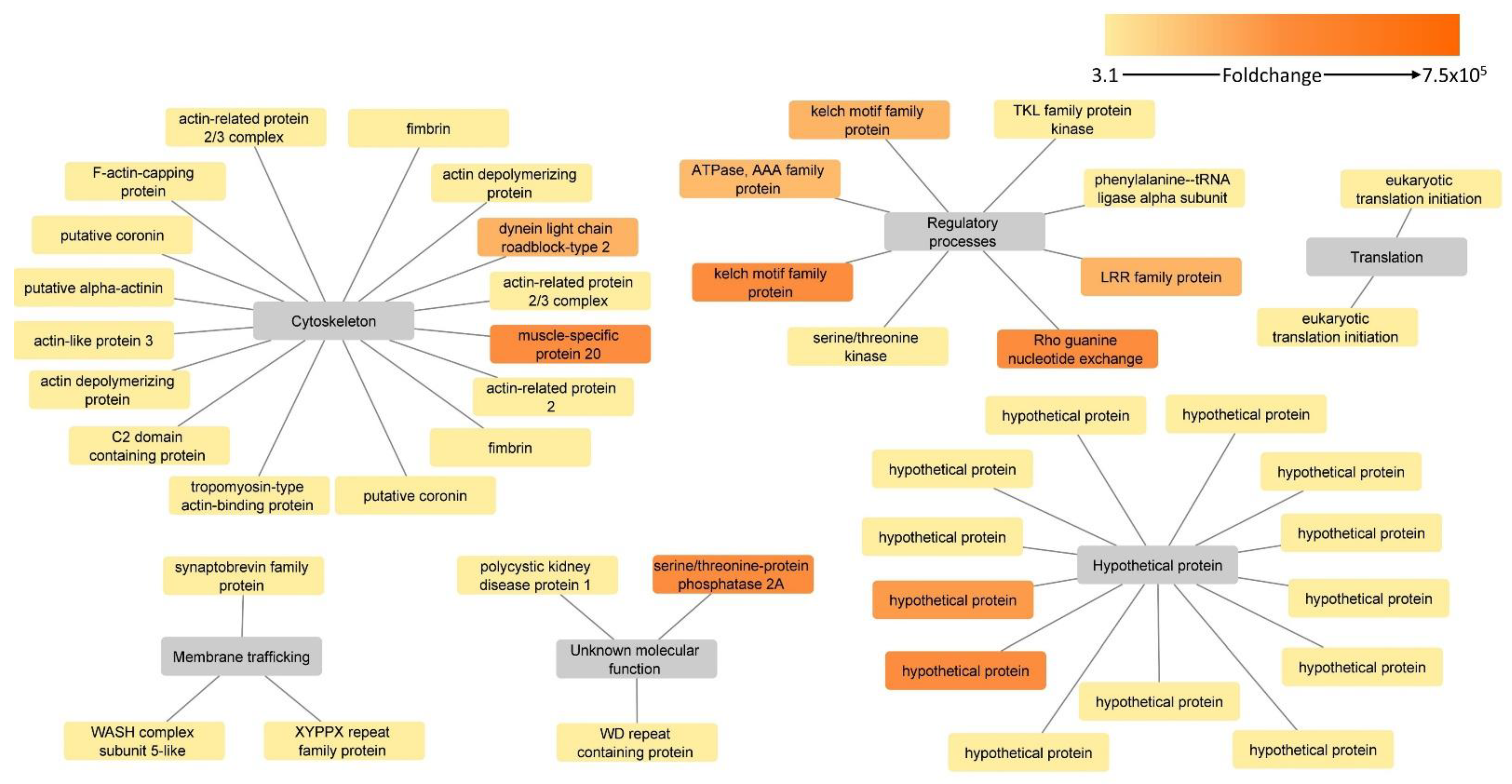
3.4. Proteins Upregulated in the H. meleagridis Attenuated Strain
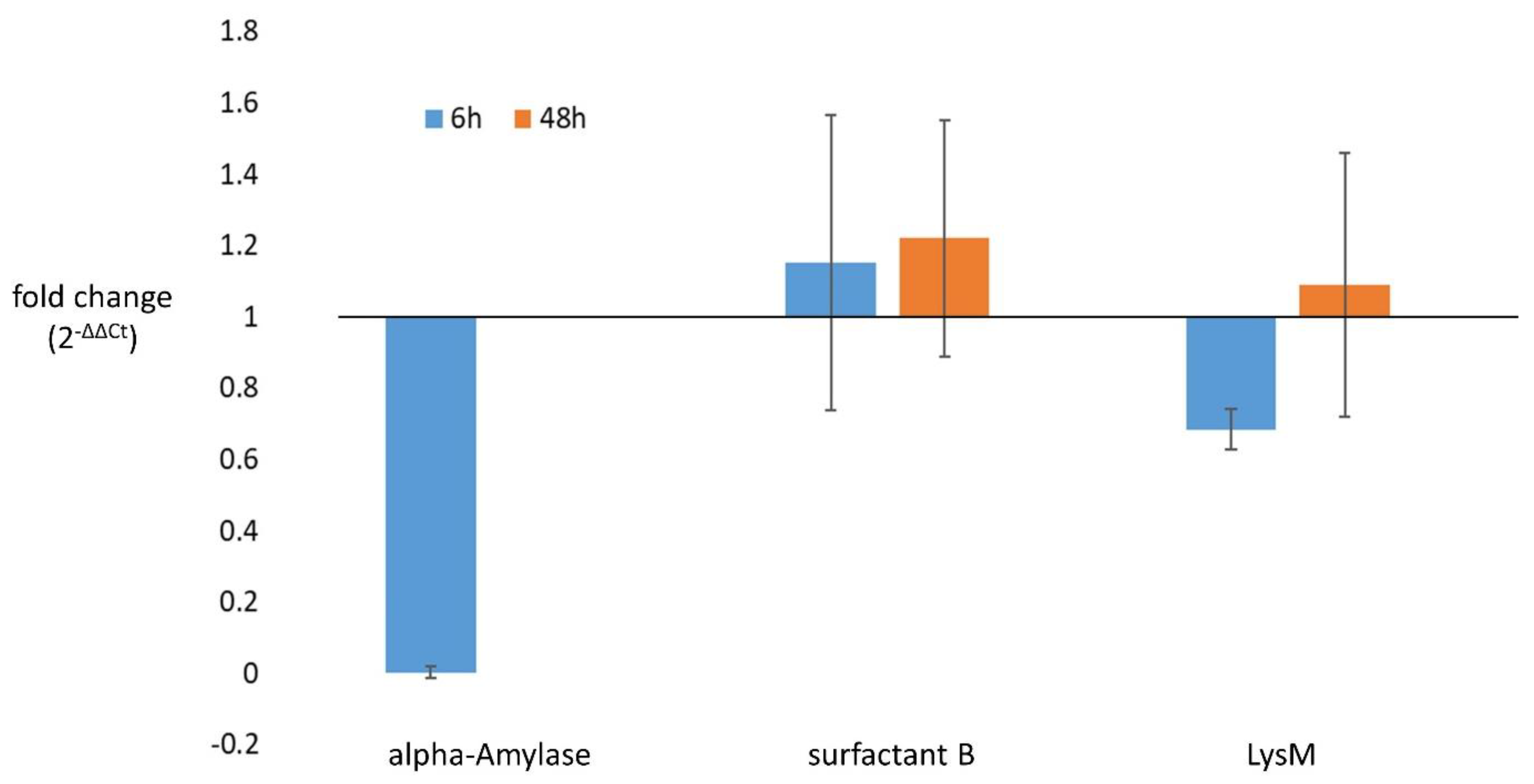
3.5. Confirmation of Differential Gene Expression in Selected Candidates
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cepicka, I.; Hampl, V.; Kulda, J. Critical Taxonomic Revision of Parabasalids with Description of One New Genus and Three New Species. Protist 2010, 161, 400–433. [Google Scholar] [CrossRef]
- Tyzzer, E.E. The flagellate character and reclassification of theparasite producing “blackhead” in turkeys-Histomonas meleagridis (Smith). J. Parasitol. 1920, 6, 124–131. [Google Scholar] [CrossRef]
- McDougald, L.R. Blackhead Disease (Histomoniasis) in Poultry: A Critical Review. Avian Dis. 2005, 49, 462–476. [Google Scholar] [CrossRef]
- Hess, M.; Liebhart, D.; Bilic, I.; Ganas, P. Histomonas meleagridis-New Insights into an Old Pathogen. Vet. Parasitol. 2015, 208, 67–76. [Google Scholar] [CrossRef]
- Liebhart, D.; Ganas, P.; Sulejmanovic, T.; Hess, M. Histomonosis in Poultry: Previous and Current Strategies for Prevention and Therapy*. Avian Pathol. 2017, 46, 1–18. [Google Scholar] [CrossRef]
- Liebhart, D.; Hess, M. Histomonosis (Blackhead Disease): A Re-Emerging Disease in Turkeys and Chickens. Avian Pathol. 2019, 49, 1–4. [Google Scholar] [CrossRef]
- Bilic, I.; Hess, M. Interplay between Histomonas meleagridis and Bacteria: Mutualistic or Predator–Prey? Trends Parasitol. 2020, 36, 232–235. [Google Scholar] [CrossRef]
- Pham, A.D.N.; Mast, J.; Magez, S.; Goddeeris, B.M.; Carpentier, S.C. The Enrichment of Histomonas meleagridis and Its Pathogen-Specific Protein Analysis: A First Step to Shed Light on Its Virulence. Avian Dis. 2016, 60, 628–636. [Google Scholar] [CrossRef]
- Monoyios, A.; Patzl, M.; Schlosser, S.; Hess, M.; Bilic, I. Unravelling the Differences: Comparative Proteomic Analysis of a Clonal Virulent and an Attenuated Histomonas meleagridis Strain. Int. J. Parasitol. 2017, 48, 145–157. [Google Scholar] [CrossRef]
- Monoyios, A.; Hummel, K.; Nöbauer, K.; Patzl, M.; Schlosser, S.; Hess, M.; Bilic, I. An Alliance of Gel-Based and Gel-Free Proteomic Techniques Displays Substantial Insight Into the Proteome of a Virulent and an Attenuated Histomonas meleagridis Strain. Front. Cell. Infect. Microbiol. 2018, 8, 407. [Google Scholar] [CrossRef] [Green Version]
- Mazumdar, R.; Endler, L.; Monoyios, A.; Hess, M.; Bilic, I. Establishment of a de Novo Reference Transcriptome of Histomonas meleagridis Reveals Basic Insights About Biological Functions and Potential Pathogenic Mechanisms of the Parasite. Protist 2017, 168, 663–685. [Google Scholar] [CrossRef]
- Mazumdar, R.; Nöbauer, K.; Hummel, K.; Hess, M.; Bilic, I. Molecular Characterization of Histomonas meleagridis Exoproteome with Emphasis on Protease Secretion and Parasite-Bacteria Interaction. PLoS ONE 2019, 14, 1–23. [Google Scholar] [CrossRef]
- Palmieri, N.; Ramires, M.; Hess, M.; Bilic, I. Complete Genomes of the Eukaryotic Poultry Parasite Histomonas meleagridis: Linking Sequence Analysis with Virulence/Attenuation. BMC Genom. 2021, 22, 1–18. [Google Scholar] [CrossRef]
- Holder, A.A. Proteins on the Surface of the Malaria Parasite and Cell Invasion. Parasitology 1994, 108, S5–S18. [Google Scholar] [CrossRef]
- Pickering, A.C.; Fitzgerald, J.R. The Role of Gram-Positive Surface Proteins in Bacterial Niche- and Host-Specialization. Front. Microbiol. 2020, 11, 1–9. [Google Scholar] [CrossRef]
- Esbelin, J.; Santos, T.; Ribière, C.; Desvaux, M.; Viala, D.; Chambon, C.; Hébraud, M. Comparison of Three Methods for Cell Surface Proteome Extraction of Listeria monocytogenes Biofilms. OMICS J. Integr. Biol. 2018, 22, 779–787. [Google Scholar] [CrossRef]
- Hess, M.; Kolbe, T.; Grabensteiner, E.; Prosl, H. Clonal Cultures of Histomonas meleagridis, Tetratrichomonas gallinarum and a Blastocystis sp. Established through Micromanipulation. Parasitology 2006, 133, 547–554. [Google Scholar] [CrossRef]
- Ganas, P.; Liebhart, D.; Glösmann, M.; Hess, C.; Hess, M. Escherichia Coli Strongly Supports the Growth of Histomonas meleagridis, in a Monoxenic Culture, without Influence on Its Pathogenicity. Int. J. Parasitol. 2012, 42, 893–901. [Google Scholar] [CrossRef]
- Blum, H.; Beier, H.; Gross, H.J. Improved Silver Staining of Plant Proteins, RNA and DNA in Polyacrylamide Gels. Electrophoresis. 1987, 8, 93–99. [Google Scholar] [CrossRef]
- Wiśniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal Sample Preparation Method for Proteome Analysis. Nat. Methods 2009, 6, 359–362. [Google Scholar] [CrossRef]
- Wiśniewski, J.R. Quantitative Evaluation of Filter Aided Sample Preparation (FASP) and Multienzyme Digestion FASP Protocols. Anal. Chem. 2016, 88, 5438–5443. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, A.M.; Sotillo, J.; Schlosser, S.; Hummel, K.; Miller, I. Towards Understanding Non-Infectious Growth-Rate Retardation in Growing Pigs. Proteomes 2019, 7, 31. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Smits, A.H.; Van Tilburg, G.B.A.; Ovaa, H.; Huber, W.; Vermeulen, M. Proteome-Wide Identification of Ubiquitin Interactions Using UbIA-MS. Nat. Protoc. 2018, 13, 530–550. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2-ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE Guidelines: Minimum Information for Publication of Quantitative Real-Time PCR Experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Elschenbroich, S.; Kim, Y.; Medin, J.A.; Kislinger, T. Isolation of Cell Surface Proteins for Mass Spectrometry-Based Proteomics. Expert Rev. Proteom. 2010, 7, 141–154. [Google Scholar] [CrossRef] [PubMed]
- Marttila, A.T.; Laitinen, O.H.; Airenne, K.J.; Kulik, T.; Bayer, E.A.; Wilchek, M.; Kulomaa, M.S. Recombinant NeutraLite Avidin: A Non-Glycosylated, Acidic Mutant of Chicken Avidin That Exhibits High Affinity for Biotin and Low Non-Specific Binding Properties. FEBS Lett. 2000, 467, 31–36. [Google Scholar] [CrossRef]
- Osborn, M.J.; Gander, J.E.; Parisi, E.; Carson, J. Mechanism of Assembly of the Outer Membrane of Salmonella Typhimurium. J. Biol. Chem. 1972, 247, 3962–3972. [Google Scholar] [CrossRef]
- McNamara, M.; Tzeng, S.C.; Maier, C.; Zhang, L.; Bermudez, L.E. Surface Proteome of “Mycobacterium avium subsp. hominissuis” during the Early Stages of Macrophage Infection. Infect. Immun. 2012, 80, 1868–1880. [Google Scholar] [CrossRef]
- Zhou, X.; Liao, W.J.; Liao, J.M.; Liao, P.; Lu, H. Ribosomal Proteins: Functions beyond the Ribosome. J. Mol. Cell Biol. 2015, 7, 92–104. [Google Scholar] [CrossRef] [Green Version]
- Nievas, Y.R.; Coceres, V.M.; Midlej, V.; de Souza, W.; Benchimol, M.; Pereira-Neves, A.; Vashisht, A.A.; Wohlschlegel, J.A.; Johnson, P.J.; de Miguel, N. Membrane-Shed Vesicles from the Parasite Trichomonas vaginalis: Characterization and Their Association with Cell Interaction. Cell. Mol. Life Sci. 2018, 75, 2211–2226. [Google Scholar] [CrossRef] [PubMed]
- De Miguel, N.; Lustig, G.; Twu, O.; Chattopadhyay, A.; Wohlschlegel, J.A.; Johnson, P.J. Proteome Analysis of the Surface of Trichomonas vaginalis Reveals Novel Proteins and Strain-Dependent Differential Expression. Mol. Cell. Proteom. 2010, 9, 1554–1566. [Google Scholar] [CrossRef] [PubMed]
- Delic, M.; Valli, M.; Graf, A.B.; Pfeffer, M.; Mattanovich, D.; Gasser, B. The Secretory Pathway: Exploring Yeast Diversity. FEMS Microbiol. Rev. 2013, 37, 872–914. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Poschmann, G.; Waldera-Lupa, D.; Rafiee, N.; Kollmann, M.; Stühler, K. OutCyte: A Novel Tool for Predicting Unconventional Protein Secretion. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef]
- Davids, B.J.; Liu, C.M.; Hanson, E.M.; Le, C.H.Y.; Ang, J.; Hanevik, K.; Fischer, M.; Radunovic, M.; Langeland, N.; Ferella, M.; et al. Identification of Conserved Candidate Vaccine Antigens in the Surface Proteome of Giardia lamblia. Infect. Immun. 2019, 87, e00219-19. [Google Scholar] [CrossRef]
- Biller, L.; Matthiesen, J.; Kühne, V.; Lotter, H.; Handal, G.; Nozaki, T.; Saito-Nakano, Y.; Schümann, M.; Roeder, T.; Tannich, E.; et al. The Cell Surface Proteome of Entamoeba histolytica. Mol. Cell. Proteom. 2014, 13, 132–144. [Google Scholar] [CrossRef]
- Giuliani, F.; Grieve, A.; Rabouille, C. Unconventional Secretion: A Stress on GRASP. Curr. Opin. Cell Biol. 2011, 23, 498–504. [Google Scholar] [CrossRef]
- Field, M.C.; Ali, B.R.S.; Field, H. GTPases in Protozoan Parasites: Tools for Cell Biology and Chemotherapy. Parasitol. Today 1999, 15, 365–371. [Google Scholar] [CrossRef]
- Zhen, Y.; Stenmark, H. Cellular Functions of Rab GTPases at a Glance. J. Cell Sci. 2015, 128, 3171–3176. [Google Scholar] [CrossRef]
- Prashar, A.; Schnettger, L.; Bernard, E.M.; Gutierrez, M.G. Rab GTPases in Immunity and Inflammation. Front. Cell. Infect. Microbiol. 2017, 7, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Bosch, D.E.; Siderovski, D.P. G Protein Signaling in the Parasite Entamoeba histolytica. Exp. Mol. Med. 2013, 45, e15-12. [Google Scholar] [CrossRef] [PubMed]
- Verma, K.; Srivastava, V.K.; Datta, S. Rab GTPases Take Centre Stage in Understanding Entamoeba histolytica Biology. Small GTPases 2020, 11, 320–333. [Google Scholar] [CrossRef] [PubMed]
- Lorentz, A.; Baumann, A.; Vitte, J.; Blank, U. The SNARE Machinery in Mast Cell Secretion. Front. Immunol. 2012, 3, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Gerst, J.E. SNAREs and SNARE Regulators in Membrane Fusion and Exocytosis. Cell. Mol. Life Sci. 1999, 55, 707–734. [Google Scholar] [CrossRef]
- Dando, P.M.; Fortunato, M.; Smith, L.; Knight, C.G.; McKendrick, J.E.; Barrett, A.J. Pig Kidney Legumain: An Asparaginyl Endopeptidase with Restricted Specificity. Biochem. J. 1999, 339, 743–749. [Google Scholar] [CrossRef]
- Arroyo, R.; Alderete, J.F. Trichomonas vaginalis Surface Proteinase Activity Is Necessary for Parasite Adherence to Epithelial Cells. Infect. Immun. 1989, 57, 2991–2997. [Google Scholar] [CrossRef]
- Rendón-Gandarilla, F.J.; Ramón-Luing, L.A.; Jaime, O.-L.; Ivone Rosa, d.A.; Benchimol, M.; Arroyo, R. The TvLEGU-1, a Legumain-Like Cysteine Proteinase, Plays a Key Role in Trichomonas vaginalis Cytoadherence. Biomed Res. Int. 2013, 15, 958–968. [Google Scholar] [CrossRef]
- Kim, H.K.; Ha, Y.R.; Yu, H.S.; Kong, H.H.; Chung, D. Il Purification and Characterization of a 33 KDa Serine Protease from Acanthamoeba lugdunensis KA/E2 Isolated from a Korean Keratitis Patient. Korean J. Parasitol. 2003, 41, 189–196. [Google Scholar] [CrossRef]
- Conseil, V.; Soête, M.; Dubremetz, J.F. Serine Protease Inhibitors Block Invasion of Host Cells by Toxoplasma gondii. Antimicrob. Agents Chemother. 1999, 43, 1358–1361. [Google Scholar] [CrossRef]
- Miller, S.A.; Binder, E.M.; Blackman, M.J.; Carruthers, V.B.; Kim, K. A Conserved Subtilisin-like Protein TgSUB1 in Microneme Organelles of Toxoplasma gondii. J. Biol. Chem. 2001, 276, 45341–45348. [Google Scholar] [CrossRef] [Green Version]
- Withers-Martinez, C.; Jean, L.; Blackman, M.J. Subtilisin-like Protease of the Malaria Parasite. Mol. Microbiol. 2004, 53, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Romano, P.; Hernández, R.; Arroyo, R.; Alderete, J.F.; López-Villaseñor, I. Identification and Characterization of a Surface-Associated, Subtilisin-like Serine Protease in Trichomonas vaginalis. Parasitology 2010, 137, 1621–1635. [Google Scholar] [CrossRef] [PubMed]
- Clark, C.G.; Diamond, L.S. Methods for Cultivation of Luminal Parasitic Protists of Clinical Importance. Clin. Microbiol. Rev. 2002, 15, 329–341. [Google Scholar] [CrossRef]
- Thibeaux, R.; Weber, C.; Hon, C.C.; Dillies, M.A.; Avé, P.; Coppée, J.Y.; Labruyère, E.; Guillén, N. Identification of the Virulence Landscape Essential for Entamoeba histolytica Invasion of the Human Colon. PLoS Pathog. 2013, 9, 1–19. [Google Scholar] [CrossRef]
- Frantz, C.; Stewart, K.M.; Weaver, V.M. The Extracellular Matrix at a Glance. J. Cell Sci. 2010, 123, 4195–4200. [Google Scholar] [CrossRef] [PubMed]
- Moncada, D.; Keller, K.; Chadee, K. Entamoeba histolytica Cysteine Proteinases Disrupt the Polymeric Structure of Colonic Mucin and Alter Its Protective Function. Infect. Immun. 2003, 71, 838–844. [Google Scholar] [CrossRef]
- Moncada, D.; Keller, K.; Chadee, K. Entamoeba histolytica-Secreted Products Degrade Colonic Mucin Oligosaccharides. Infect. Immun. 2005, 73, 3790–3793. [Google Scholar] [CrossRef] [PubMed]
- Tovy, A.; Hertz, R.; Siman-Tov, R.; Syan, S.; Faust, D.; Guillen, N.; Ankri, S. Glucose Starvation Boosts Entamoeba histolytica Virulence. PLoS Negl. Trop. Dis. 2011, 5, e1247. [Google Scholar] [CrossRef]
- Mesnage, S.; Dellarole, M.; Baxter, N.J.; Rouget, J.B.; Dimitrov, J.D.; Wang, N.; Fujimoto, Y.; Hounslow, A.M.; Lacroix-Desmazes, S.; Fukase, K.; et al. Molecular Basis for Bacterial Peptidoglycan Recognition by LysM Domains. Nat. Commun. 2014, 5, 4269. [Google Scholar] [CrossRef]
- Hirschhausen, N.; Schlesier, T.; Peters, G.; Heilmann, C. Characterization of the Modular Design of the Autolysin/Adhesin Aaa from Staphylococcus aureus. PLoS ONE 2012, 7, e40353. [Google Scholar] [CrossRef]
- Abdelhamid, M.K.; Quijada, N.M.; Dzieciol, M.; Hatfaludi, T.; Bilic, I.; Selberherr, E.; Liebhart, D.; Hess, C.; Hess, M.; Paudel, S. Co-Infection of Chicken Layers With Histomonas meleagridis and Avian Pathogenic Escherichia coli Is Associated With Dysbiosis, Cecal Colonization and Translocation of the Bacteria From the Gut Lumen. Front. Microbiol. 2020, 11, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Abdelhamid, M.K.; Rychlik, I.; Hess, C.; Hatfaludi, T.; Crhanova, M.; Karasova, D.; Lagler, J.; Liebhart, D.; Hess, M.; Paudel, S. Typhlitis Induced by Histomonas meleagridis Affects Relative but Not the Absolute Escherichia coli Counts and Invasion in the Gut in Turkeys. Vet. Res. 2021, 52, 92. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Y.; Saier, M.H. The Amoebapore Superfamily. Biochim. Biophys. Acta Rev. Biomembr. 2000, 1469, 87–99. [Google Scholar] [CrossRef]
- Bruhn, H. A Short Guided Tour through Functional and Structural Features of Saposin-like Proteins. Biochem. J. 2005, 389, 249–257. [Google Scholar] [CrossRef]
- Bujanover, S.; Katz, U.; Bracha, R.; Mirelman, D. A Virulence Attenuated Amoebapore-Less Mutant of Entamoeba histolytica and Its Interaction with Host Cells. Int. J. Parasitol. 2003, 33, 1655–1663. [Google Scholar] [CrossRef]
- Bracha, R.; Nuchamowitz, Y.; Mirelman, D. Transcriptional Silencing of an Amoebapore Gene in Entamoeba histolytica: Molecular Analysis and Effect on Pathogenicity. Eukaryot. Cell 2003, 2, 295–305. [Google Scholar] [CrossRef]
- Ralston, K.S.; Petri, W.A. Tissue Destruction and Invasion by Entamoeba histolytica. Trends Parasitol. 2011, 27, 254–263. [Google Scholar] [CrossRef]
- Hawgood, S.; Derrick, M.; Poulain, F. Structure and Properties of Surfactant Protein B. Biochim. Biophys. Acta Mol. Basis Dis. 1998, 1408, 150–160. [Google Scholar] [CrossRef]
- Hirt, R.P.; de Miguel, N.; Nakjang, S.; Dessi, D.; Liu, Y.C.; Diaz, N.; Rappelli, P.; Acosta-Serrano, A.; Fiori, P.L.; Mottram, J.C. Trichomonas vaginalis Pathobiology. New Insights from the Genome Sequence. Adv. Parasitol. 2011, 77, 87–140. [Google Scholar] [CrossRef]
- Gao, J.; Nakamura, F. Actin-Associated Proteins and Small Molecules Targeting the Actin Cytoskeleton. Int. J. Mol. Sci. 2022, 23, 2118. [Google Scholar] [CrossRef]
- Lappalainen, P. Actin-Binding Proteins: The Long Road to Understanding the Dynamic Landscape of Cellular Actin Networks. Mol. Biol. Cell 2016, 27, 2519–2522. [Google Scholar] [CrossRef] [PubMed]
- Gruber, J.; Ganas, P.; Hess, M. Long-Term in Vitro Cultivation of Histomonas meleagridis Coincides with the Dominance of a Very Distinct Phenotype of the Parasite Exhibiting Increased Tenacity and Improved Cell Yields. Parasitology 2017, 144, 1253–1263. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.L.; Long, P.L.; Millard, B.J.; Bradley, J. The Fine Structure and Method of Feeding of the Tissue Parasitizing Stages of Histomonas meleagridis. Parasitology 1969, 59, 171–184. [Google Scholar] [CrossRef] [PubMed]
- Hussain, I.; Jaskulska, B.; Hess, M.; Bilic, I. Detection and Quantification of Histomonas meleagridis by Real-Time PCR Targeting Single Copy Genes. Vet. Parasitol. 2015, 212, 382–388. [Google Scholar] [CrossRef] [PubMed]
- Perez-Riverol, Y.; Bai, J.; Bandla, C.; García-Seisdedos, D.; Hewapathirana, S.; Kamatchinathan, S.; Kundu, D.J.; Prakash, A.; Frericks-Zipper, A.; Eisenacher, M.; et al. The PRIDE Database Resources in 2022: A Hub for Mass Spectrometry-Based Proteomics Evidences. Nucleic Acids Res. 2022, 50, D543–D552. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession | Protein Name | MW (kDa) | Unique Peptides | Tryptic Peptides | Fold Change | # of TM Domains | Signal-Peptide Prediction | Non-Classical Secretion | Re-Analysis of Proteome LCMS [10] | Re-Analysis of Exoproteome LCMS [12] |
|---|---|---|---|---|---|---|---|---|---|---|
| Peptidases | ||||||||||
| KAH0796674 | Clan SC, family S33, methylesterase-like serine peptidase | 39.3 | 8 | 8 | 4.4 | 0 | No | Secreted | Yes—not upregulated | No |
| KAH0805360 | Clan CD, family C13, asparaginyl endopeptidase-like cysteine peptidase | 44.8 | 2 | 2 | ON/OFF * | 0 | No | Secreted | - | No |
| KAH0803400 | Clan SC, family S33, methylesterase-like serine peptidase.1 | 38.9 | 6 | 6 | 7.3 | 0 | No | No | Yes—upregulated (2.12-fold) | No |
| Metabolic processes | ||||||||||
| KAH0798244 | Class I SAM-dependent methyltransferase | 30.4 | 2 | 2 | ON/OFF * | 0 | No | No | No | No |
| KAH0797675 | Alpha-amylase, catalytic domain-containing protein | 59.6 | 1 | 10 | ON/OFF * | 1 | No | Secreted | No | No |
| KAH0804379 | Serine palmitoyltransferase | 52.7 | 2 | 3 | 8.6 | 0 | No | Secreted | Yes—not upregulated | No |
| KAH0804812 | LysM peptidoglycan binding domain-containing protein.1 | 32.2 | 3 | 7 | 3.7 | 0 | Yes | No | Yes—upregulated (5.38-fold) | Yes |
| KAH0787061 | Surfactant B protein | 44.1 | 2 | 10 | 9.0 | 0 | No | No | Yes—not upregulated | Yes |
| KAH0800457 | Glycoside hydrolase family 20 | 12.3 | 2 | 5 | 47.5 | 0 | No | Secreted | Yes—upregulated (35.5-fold) | No |
| KAH0799877 | Acyltransferase family protein | 36.2 | 2 | 2 | 7.7 | 3 | No | No | No | No |
| Membrane trafficking | ||||||||||
| KAH0804263 | Cation efflux family protein | 52.5 | 2 | 2 | ON/OFF * | 6 | No | No | No | No |
| KAH0802276 | XYPPX repeat family protein/C2 domain-containing protein | 32.2 | 1 | 2 | 4.4 | 0 | No | No | Yes—upregulated (3.07-fold) | No |
| KAH0801182 | V-type proton ATPase subunit C | 47.3 | 1 | 3 | ON/OFF * | 0 | No | No | Yes—not upregulated | No |
| Ribosomal proteins | ||||||||||
| KAH0796569 | Ribosomal protein L21e.1 | 18.6 | 1 | 5 | 4.7 | 0 | No | Secreted | Yes—not upregulated | No |
| KAH0796694 | 40S ribosomal protein S17-B | 14.8 | 1 | 7 | ON/OFF * | 0 | No | Secreted | Yes—not upregulated | No |
| KAH0802566 | 60S ribosomal protein L30 | 12.3 | 1 | 5 | 10.7 | 0 | No | No | Yes—not upregulated | No |
| KAH0798245 | Ribosomal protein L18ae | 20 | 1 | 11 | ON/OFF * | 0 | No | No | No | Yes |
| Signaling | ||||||||||
| KAH0796629 | Rab family GTPase | 19.4 | 1 | 2 | ON/OFF * | 0 | No | Secreted | No | No |
| KAH0806080 | Ras family GTPase | 21.7 | 7 | 7 | 216.8 | 0 | No | No | Yes—not upregulated | No |
| KAH0802584 | Heat shock 70 kDa protein | 72.4 | 5 | 6 | 101.8 | 0 | Yes | No | Yes—not upregulated | Yes |
| KAH0798120 | Rab family GTPase | 24.3 | 1 | 2 | ON/OFF * | 0 | No | No | No | No |
| Hypothetical proteins | ||||||||||
| KAH0799077 | Hypothetical protein/Formin | 36.9 | 1 | 2 | 4.4 | 0 | No | No | No | No |
| Accession | Protein name | MW (kDa) | Unique Peptides | Tryptic Peptides | Fold Change | # of TM Domains | Signal-Peptide Prediction | Non-Classical Secretion | Re-Analysis of Proteome LCMS [10] | Re-Analysis of Exoproteome LCMS [12] |
|---|---|---|---|---|---|---|---|---|---|---|
| Cytoskeleton | ||||||||||
| KAH0806015 | Actin-related protein 2 | 44.5 | 11 | 11 | 3.3 | 0 | No | No | Yes—not upregulated | No |
| KAH0803799 | Fimbrin | 68.5 | 1 | 16 | 7.8 | 1 | No | No | No | No |
| KAH0803847 | Actin depolymerizing protein | 35.7 | 5 | 7 | 8.6 | 0 | No | No | Yes—not upregulated | No |
| KAH0804054 | Dynein light chain roadblock-type 2 | 11.2 | 2 | 2 | ON/OFF * | 0 | No | No | No | No |
| KAH0807157 | Putative alpha-actinin | 130.1 | 86 | 86 | 42.8 | 0 | No | No | Yes—not upregulated | No |
| KAH0807177 | Actin-like protein 3 | 47.3 | 2 | 14 | 3.4 | 0 | No | Secreted | No | No |
| KAH0803330 | F-actin-capping protein subunit beta | 30.6 | 1 | 10 | 6.1 | 0 | No | Secreted | No | No |
| KAH0801303 | Cofilin/tropomyosin-type actin-binding protein | 16.1 | 4 | 4 | 5.1 | 0 | No | No | No | No |
| KAH0800820 | C2 domain-containing protein/CH-domain-containing protein | 46.6 | 10 | 10 | 3.9 | 0 | No | No | No | No |
| KAH0799687 | Putative coronin | 94.8 | 4 | 32 | 21.6 | 0 | No | No | No | No |
| KAH0806391 | Putative coronin | 91.3 | 2 | 30 | 8.0 | 0 | No | No | No | No |
| KAH0799604 | Actin depolymerizing protein | 35.8 | 6 | 8 | 7.1 | 0 | No | No | No | No |
| KAH0798726 | Muscle-specific protein 20 | 47.9 | 3 | 3 | ON/OFF * | 0 | No | No | No | No |
| KAH0797693 | Fimbrin | 70.5 | 1 | 16 | 7.2 | 0 | No | No | No | No |
| KAH0797549 | Actin-related protein 2/3 complex, subunit 1 | 40 | 2 | 17 | 4.8 | 0 | No | No | No | No |
| KAH0797350 | Actin-related protein 2/3 complex subunit 2 | 34.3 | 7 | 12 | 3.4 | 0 | No | Secreted | No | No |
| Hypothetical proteins | ||||||||||
| KAH0806065 | Hypothetical protein.5 | 14.2 | 2 | 2 | ON/OFF | 0 | No | No | No | No |
| KAH0806131 | Hypothetical protein.157 | 36.9 | 2 | 3 | 8.1 | 0 | No | No | Yes—not upregulated | No |
| KAH0805781 | Hypothetical protein | 55.8 | 3 | 3 | 8.3 | 0 | No | No | Yes—not upregulated | No |
| KAH0805381 | Hypothetical protein.62 | 64.3 | 11 | 11 | 3.8 | 0 | No | No | Yes—not upregulated | No |
| KAH0804660 | Hypothetical protein.92 | 25.6 | 6 | 6 | 7.2 | 0 | No | No | Yes—not upregulated | No |
| KAH0807132 | Hypothetical protein.68 | 62.3 | 9 | 9 | 4.8 | 0 | No | No | Yes—not upregulated | No |
| KAH0802306 | Hypothetical protein.128 | 88.2 | 3 | 3 | 4.6 | 0 | No | No | No | No |
| KAH0800233 | Hypothetical protein.81 | 23.3 | 8 | 8 | 3.2 | 0 | No | No | No | No |
| KAH0798642 | Hypothetical protein.60 | 116.8 | 9 | 9 | 3.3 | 0 | No | Secreted | No | No |
| KAH0806186 | Hypothetical protein.3 | 82.3 | 2 | 2 | ON/OFF * | 0 | No | Secreted | No | No |
| KAH0798386 | Hypothetical protein.111 | 26.3 | 6 | 6 | 4.8 | 0 | No | No | Yes—not upregulated | Yes |
| KAH0798396 | Hypothetical protein.88 | 41.5 | 2 | 9 | 17.3 | 0 | No | No | Yes—not upregulated | No |
| KAH0798145 | Hypothetical protein.153 | 91.8 | 4 | 4 | 3.1 | 0 | No | No | Yes—not upregulated | No |
| Regulatory processes | ||||||||||
| KAH0796192 | Protein serine/threonine kinase, putative | 124.9 | 8 | 8 | 10.3 | 0 | No | Secreted | Yes—not upregulated | Yes |
| KAH0796421 | Leucine Rich Repeat family protein | 83.9 | 2 | 2 | ON/OFF * | 0 | No | No | No | No |
| KAH0804216 | Kelch motif family protein | 199.7 | 2 | 2 | ON/OFF * | 0 | No | No | Yes—not upregulated | No |
| KAH0804546 | Kelch motif family protein | 137.4 | 4 | 4 | ON/OFF * | 0 | No | No | Yes—not upregulated | No |
| KAH0806868 | TKL family protein kinase | 135.2 | 2 | 2 | 5.8 | 0 | No | No | Yes—not upregulated | No |
| KAH0802085 | Rho guanine nucleotide exchange factor 39 | 33.6 | 2 | 2 | ON/OFF * | 0 | No | No | No | No |
| KAH0806401 | ATPase, AAA family protein | 97.5 | 2 | 2 | ON/OFF * | 0 | No | No | Yes—not upregulated | No |
| KAH0798081 | Phenylalanine–tRNA ligase alpha subunit | 61.3 | 2 | 2 | 25.3 | 0 | No | Secreted | Yes—not upregulated | No |
| Membrane trafficking/transport | ||||||||||
| KAH0796205 | Synaptobrevin family protein | 25 | 1 | 4 | 5.4 | 0 | No | No | Yes—not upregulated | Yes |
| KAH0802328 | WASH complex subunit 5-like | 131.5 | 1 | 3 | 3.5 | 0 | No | No | Yes—not upregulated | Yes |
| KAH0797426 | XYPPX repeat family protein | 35.8 | 2 | 2 | 11.0 | 0 | No | No | Yes—not upregulated | No |
| Translation | ||||||||||
| KAH0796670 | Eukaryotic translation initiation factor 3 subunit C isoform X1 | 80.9 | 12 | 14 | 3.2 | 0 | No | No | Yes—not upregulated | No |
| KAH0805651 | Eukaryotic translation initiation factor 3 subunit 8 N-terminus-domain-containing protein | 81.7 | 9 | 11 | 3.2 | 0 | No | No | Yes—not upregulated | Yes |
| Unknown molecular function | ||||||||||
| KAH0796283 | Serine/threonine-protein phosphatase 2A 65 kDa regulatory subunit A alpha isoform/HEAT repeat family protein | 41.4 | 1 | 4 | ON/OFF * | 0 | No | No | Yes—not upregulated | Yes |
| KAH0796931 | WD repeat-containing protein 5B isoform X2 | 39.3 | 2 | 2 | 7.9 | 0 | No | No | No | Yes |
| KAH0799325 | Polycystic kidney disease protein 1-like 3 | 30.4 | 2 | 2 | 8.9 | 0 | No | No | No | No |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramires, M.d.J.; Hummel, K.; Hatfaludi, T.; Riedl, P.; Hess, M.; Bilic, I. Comparative Surfaceome Analysis of Clonal Histomonas meleagridis Strains with Different Pathogenicity Reveals Strain-Dependent Profiles. Microorganisms 2022, 10, 1884. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10101884
Ramires MdJ, Hummel K, Hatfaludi T, Riedl P, Hess M, Bilic I. Comparative Surfaceome Analysis of Clonal Histomonas meleagridis Strains with Different Pathogenicity Reveals Strain-Dependent Profiles. Microorganisms. 2022; 10(10):1884. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10101884
Chicago/Turabian StyleRamires, Marcelo de Jesus, Karin Hummel, Tamas Hatfaludi, Petra Riedl, Michael Hess, and Ivana Bilic. 2022. "Comparative Surfaceome Analysis of Clonal Histomonas meleagridis Strains with Different Pathogenicity Reveals Strain-Dependent Profiles" Microorganisms 10, no. 10: 1884. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10101884