
Divergent Changes in Bacterial Functionality as Affected by Root-Zone Ecological Restoration in an Aged Peach Orchard

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Soil Property Determination
2.3. DNA Extraction, PCR Amplification and High-Throughput Sequencing
2.4. Data Analysis
3. Results
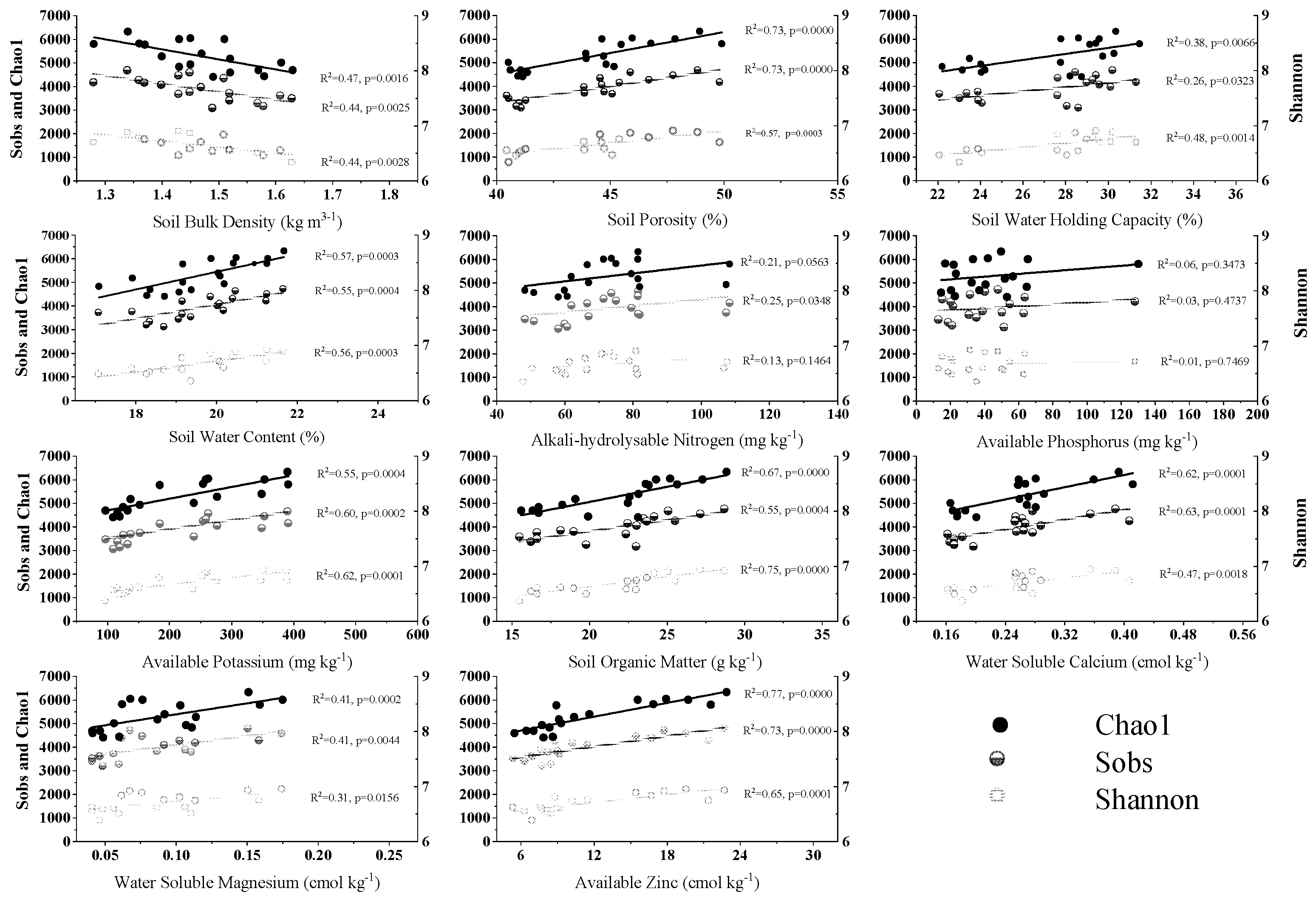
3.1. Soil Properties Affected by RERP
3.2. Alpha Diversity of Soil Bacterial Community Affected by RERP
3.3. Beta Diversity of Soil Bacterial Community Affected by RERP
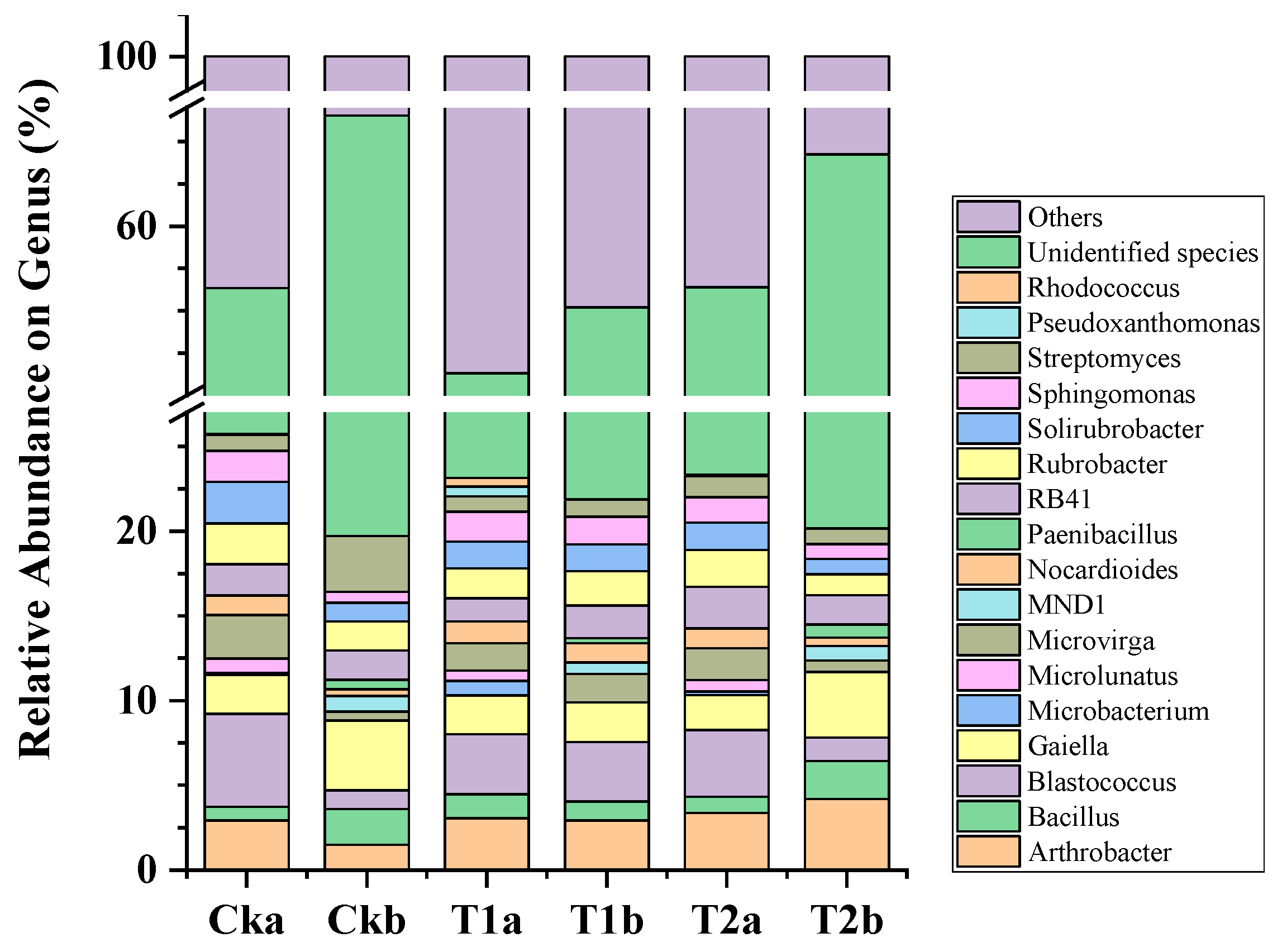
3.4. Soil Bacterial Functionality
4. Discussion
4.1. Soil Bacterial Diversity as Affected by RERP
4.2. The structure Soil Bacterial Functionality as Affected by RERP
4.3. Soil Bacterial Functionality as Affected by Soil Physicochemical Properties
4.4. Soil Bacterial Functionality as Affected by Alpha and Beta Diversities
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wang, H.; Yang, Z.; Cong, P.; Cao, Y.; Cheng, C.; Liu, F.; Zhou, Z.; Xu, G.; Wang, W.; Hu, C.; et al. Fruit industry mission and 14th five-year key tasks in China. China Fruits 2022, 4, 1–4. [Google Scholar]
- Shu, H. Study on sustainable development strategy of fruit industry in China. Dec. Fruits 2012, 44, 1–4. [Google Scholar]
- Bai, Y.; Xuan, J.; Li, L. Present situation and countermeasures of old orchard reconstruction in China. North. Fruits 2019, 3, 47–49. [Google Scholar]
- Shu, H.; Chen, X. The current task of the development of fruits industry in China. China Fruits 2018, 2, 1–3. [Google Scholar]
- Chen, Y.; Wen, X.; Sun, Y.; Zhang, J.; Wu, W.; Liao, Y. Mulching practices altered soil bacterial community structure and improved orchard productivity and apple quality after five growing seasons. Sci. Hortic. 2014, 172, 248–257. [Google Scholar] [CrossRef]
- Liang, B.; Ma, C.; Fan, L.; Wang, Y.; Yuan, Y. Soil amendment alters soil physicochemical properties and bacterial community structure of a replanted apple orchard. Microbiol. Res. 2018, 216, 1–11. [Google Scholar] [CrossRef]
- Wang, L.; Li, J.; Yang, F.; E, Y.; Yuan, J.; Raza, W.; Huang, Q.; Shen, Q. Long term application of bioorganic fertilizer improved soil biochemical properties and microbial communities of an apple orchard soil. Front. Microbiol. 2016, 7, 1893. [Google Scholar] [CrossRef]
- Wang, L.; Li, J.; Yang, F.; E, Y.; Raza, W.; Huang, Q.; Shen, Q. Application of bioorganic fertilizer significantly increased apple yields and shaped bacterial community structure in orchard soil. Microb. Ecol. 2017, 27, 404–416. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, L.; Yuan, Y.; Xu, J.; Tu, C.; Fisk, C.; Zhang, W.; Chen, X.; Ritchie, D.; Hu, S. Irrigation and weed control alter soil microbiology and nutrient availability in North Carolina Sandhill peach orchards. Sci. Total Environ. 2018, 615, 517–525. [Google Scholar] [CrossRef]
- Yang, H.; Shu, H. Studies on Apple Roots; Science Press: Beijing, China, 2007; pp. 103–104. [Google Scholar]
- Bai, J.; Li, N.; Fu, C.; Zhang, Z.; Wang, Y.; Shu, H. Effect of Micro-environment Regulation Mode Around Root Zone on the Soil Property and Root Growth of Apple Trees. Acta Hortic. Sinica 2016, 43, 829–840. [Google Scholar]
- Qin, L.; Wei, Q.; Li, J.; Wang, X.; Zou, Y.; Gao, Z.; Liu, J. Effects of soil amended model in root-zone on growth of grape roots. Chin. Agric. Sci. Bull. 2005, 21, 270–272. [Google Scholar]
- Qin, S.; Zhang, Y.; Xuan, J.; Lv, D. Mountain apple orchards straw mulching combined with acupuncture point and water storage technology. North. Fruits 2015, 5, 29–30. [Google Scholar]
- Zhang, W.; Chen, Y.; Li, X.; Li, Y.; Tian, X. Effects of balanced dressing of N, P and K fertilizers by injecting fertilizer solution into root zone on apple yield and quality in Weibei Dryland. J. Northwest A F Univ.-Nat. Sci. Ed. 2018, 46, 59–68. [Google Scholar]
- Peck, G.M.; Merwin, I.A.; Thies, J.E.; Schindelbeck, R.R.; Brown, G.B. Soil properties change during the transition to integrated and organic apple production in a New York orchard. Appl. Soil Ecol. 2011, 48, 18–30. [Google Scholar] [CrossRef]
- Yim, B.; Smalla, K.; Winkelmann, T. Evaluation of apple replant problems based on different soil disinfection treatments—Links to soil microbial community structure? Plant Soil 2013, 366, 617–631. [Google Scholar] [CrossRef]
- Dangi, S.R.; Zhang, H.; Wang, D.; Gerik, J.; Hanson, B.D. Soil Microbial Community Composition in a Peach Orchard Under Different Irrigation Methods and Postharvest Deficit Irrigation. Soil Sci. 2016, 181, 208–215. [Google Scholar] [CrossRef]
- Zhang, H.; Cao, Y.; Lyu, J. Decomposition of different crop straws and variation in straw-associated microbial communities in a peach orchard, China. J. Arid Land 2021, 13, 152–164. [Google Scholar] [CrossRef]
- Yang, J.; Ruegger, P.M.; McKenry, M.V.; Becker, J.O.; Borneman, J. Correlations between root-associated microorganisms and peach replant disease symptoms in a California soil. PLoS ONE 2012, 7, e46420. [Google Scholar] [CrossRef] [Green Version]
- Antwis, R.E.; Griffiths, S.H.; Harrison, X.A.; Brailsford, F.L.; De Menezes, A.; Devaynes, A.; Forbes, K.M.; Fry, E.L.; De Menezes, A.; Devaynes, A.; et al. Fifty important research questions in microbial ecology. FEMS Microbiol. Ecol. 2017, 93, fix044. [Google Scholar] [CrossRef]
- Bahram, M.; Hildebrand, F.; Forslund, S.K.; Anderson, J.L.; Soudzilovskaia, N.A.; Bodegom, P.M.; Bengtssom-Palme, J.; Anslan, S.; Coelho, L.P.; Harend, H.; et al. Structure and function of the global topsoil microbiome. Nature 2018, 560, 233–237. [Google Scholar] [CrossRef] [Green Version]
- Delgado-Baquerizo, M.; Maestre, F.T.; Reich, P.B.; Jeffries, T.C.; Gaitan, J.J.; Encinar, D.; Berdugo, M.; Campbell, C.D.; Singh, B.K. Microbial diversity drives multifunctionality in terrestrial ecosystems. Nat. Commun. 2016, 7, 10541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, X.; Zhou, Y.; Xia, Y.; Chen, L.; Li, T.; Zhao, H. Effects of Paclobutrazol on Soil Bacterial Diversity in Mango Orchard and PICRUSt-based Predicted Metagenomic Analysis. Chin. J. Trop. Crops 2019, 40, 807–814. [Google Scholar]
- Walter, K.; Don, A.; Tiemeyer, B.; Freibauer, A. Determining soil bulk density for carbon stock calculations: A systematic method comparison. Soil Sci. Soc. Am. J. 2016, 80, 579–591. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Qin, S.; Lv, D.; Wang, B. Isolation of dominant Actinomycetes from cherry orchard soil and their functions in biocontrol and plant promotion. Sci. Agric. Sin. 2013, 46, 1221–1229. [Google Scholar]
- Lv, Y.; Qin, S. Soil Testing and Fertilization; China Agriculture Press: Beijing, China, 2002; pp. 33–37. [Google Scholar]
- GB 7172-87; Method for the Determination of Soil Water Content. National Bureau of Standards of the People’s Republic of China: Beijing, China, 1987.
- NY/T 85-1988; Method for Determination of Soil Organic Matter. Ministry of Agriculture of the People’s Republic of China: Beijing, China, 1988.
- Lu, R. Soil Agricultural Chemistry Analysis Method; China Agricultural Scientech Press: Beijing, China, 1999; pp. 150–152. [Google Scholar]
- Bao, S. Soil Agrochemical Analysis, 3rd ed.; China Agriculture Press: Beijing, China, 2000. [Google Scholar]
- NY/T 3242-2018; Method for the Determination of Water-Soluble Calcium and Water-Soluble Magnesium in Soil. Ministry of Agriculture and Rural Affaires of the People’s Republic of China: Beijing, China, 2018.
- NY/T 890-2004; Determination of Available Zinc, Manganese, Iron, Copper in Soil-Extraction with Buffered DTPA Solution. Ministry of Agriculture of the People’s Republic of China: Beijing, China, 2004.
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microb. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Ding, J.; Zhu, Y.; He, J.; Hu, H. Soil bacterial taxonomic diversity is critical to maintaining the plant productivity. Environ. Int. 2020, 140, 105766. [Google Scholar] [CrossRef]
- Louca, S.; Jacques, S.M.S.; Pires, A.P.F.; Leal, J.S.; Srivastava, D.S.; Parfrey, L.W.; Farjalla, V.F.; Doebeli, M. High taxonomic variability despite stable functional structure across microbial communities. Nat. Ecol. Evol. 2017, 1, 0015. [Google Scholar] [CrossRef]
- Ramos, M.F.; Almeida, W.R.S.; Amaral, R.L.; Suzuki, L.E.A.S. Degree of compactness and soil quality of peach orchards with different production ages. Soil Till. Res. 2022, 219, 105324. [Google Scholar] [CrossRef]
- Beth, F.T.; Brockett, C.E.; Grayston, S.J. Soil moisture is the major factor influencing microbial community structure and enzyme activities across seven biogeoclimatic zones in western Canada. Soil Biol. Biochem. 2012, 44, 9–20. [Google Scholar]
- Budge, K.; Leifeld, J.; Hiltbrunner, E.; Fuhrer, J. Litter quality and pH are strong drivers of carbon turnover and distribution in alpine grassland soils. Biogeosci. Discuss. 2010, 7, 6207–6242. [Google Scholar]
- Luo, G.; Rensing, C.; Chen, H.; Liu, M.; Wang, M.; Guo, S.; Ling, N.; Shen, Q.; Briones, M. Deciphering the associations between soil microbial diversity and ecosystem multifunctionality driven by long-term fertilization management. Funct. Ecol. 2018, 32, 1103–1116. [Google Scholar] [CrossRef]
- Lun, A.; Zhao, M.; Li, X.; Sun, G. Different Urban Forest Tree Species Affect the Assembly of the Soil Bacterial and Fungal Community. Microb. Ecol. 2022, 83, 447–458. [Google Scholar]
- Sun, Y.; Ma, J.; Fu, G.; Zhang, P.; Ma, R.; Sun, J.; Dong, K. Effects of soil conditioner and growing grass on yield and soil chemical properties in orchard acidic soils. Soils Fert. Sci. China 2021, 2, 61–68. [Google Scholar]
- Liu, X.; Wei, T. High-throughput Sequencing Analysis of Soil Bacterial Community in the Grain for Green Project Areas of the Loess Plateau. Environ. Sci. 2021, 42, 4489–4499. [Google Scholar]
- Wang, Y.; Liu, L.; Yang, J.; Duan, Y.; Luo, Y.; Taherzadeh, M.J.; Li, Y.; Li, H.; Awasthi, M.K.; Zhao, Z. The diversity of microbial community and function varied in response to different agricultural residues composting. Sci. Total Environ. 2020, 715, 136983. [Google Scholar] [CrossRef]
- Chen, L.; Brookes, P.C.; Xu, J.; Zhang, J.; Zhang, C.; Zhou, X.; Luo, Y. Structural and functional differentiation of the root-associated bacterial microbiomes of perennial ryegrass. Soil Biol. Biochem. 2016, 98, 1–10. [Google Scholar] [CrossRef]
- Duan, Y.; Awasthi, S.K.; Liu, T.; Verma, S.; Wang, Q.; Chen, H.; Awasthi, M.K. Positive impact of biochar alone and combined with bacterial consortium amendment on improvement of bacterial community during cow manure composting. Bioresour. Technol. 2019, 280, 79–87. [Google Scholar] [CrossRef]
- Wu, R.; Cheng, X.; Han, H. The Effect of Forest Thinning on Soil Microbial Community Structure and Function. Forests 2019, 10, 352. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.; Du, Y.; Gao, G.; Zhang, Y.; Cao, H.; Zhu, B.; Yang, S.; Zhang, J.; Qiu, Y.; Liu, H. Soil bacterial community structure and functional prediction of Pinus sylvestrisvar mongolica plantation in the HulunBuir Sandy Land. Acta Ecol. Sin. 2021, 41, 4131–4139. [Google Scholar]
- Ma, X.; Luo, Z.; Zhang, Y.; Liu, J.; Niu, Y.; Cai, L. Distribution characteristics and ecological function predictions of soil bacterial communities in rainfed alfalfa fields on the Loess Plateau. Acta Pratac. Sin. 2021, 30, 54–67. [Google Scholar]
- Zhang, J.; Xu, M. Characteristic of soil bacterial community diversity among different vegetation types in the Loss hilly region. J. Ecol. Rur. Environ. 2022, 38, 225–235. [Google Scholar]
- Yan, Y.; Kuramae, E.E.; Hollander, M.; Klinkhamer, P.; Veen, J. Functional traits dominate the diversity-related selection of bacterial communities in the rhizosphere. ISME J. 2017, 11, 56–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falkowski, P.G.; Fenchel, T.; Delong, E.F. The microbial engines that drive Earth’s biogeochemical cycles. Science 2008, 320, 1034–1039. [Google Scholar] [CrossRef] [Green Version]
- Xun, W.; Li, W.; Xiong, W.; Ren, Y.; Liu, Y.; Miao, Y.; Xu, Z.; Zhang, N.; Shen, Q.; Zhang, R. Diversity-triggered deterministic bacterial assembly constrains community functions. Nat. Commun. 2019, 10, 3833. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y. Spatial and Temporal Distribution of Soil Microorganisms in the Rhizospherer of Ulmus Pumila Var. Sabulosa within Otindag Sandy Land. Ph.D. Thesis, Inner Mongolia Agricultural University, Hohhot, China, 2019. [Google Scholar]
- Cole, J.C.; Smith, M.W.; Penn, C.J.; Cheary, B.S.; Conaghan, K.J. Nitrogen, phosphorus, calcium, and magnesium applied individually or as a slow release or controlled release fertilizer increase growth and yield and affect macronutrient and micronutrient concentration and content of field-grown tomato plants. Sci. Hortic. 2016, 211, 420–430. [Google Scholar] [CrossRef] [Green Version]
- Amiri, M.E.; Fallahi, E.; Golchin, A. Influence of Foliar and Ground Fertilization on Yield, Fruit Quality, and Soil, Leaf, and Fruit Mineral Nutrients in Apple. J. Plant Nutr. 2008, 31, 515–525. [Google Scholar] [CrossRef]
- Moyin-Jesu, E.I. Impact of Different Organic Fertilizers Application on Soil Fertility Improvement, Growth and Fruit Yield Parameters of Pineapple (Ananas comosus L). J. Exp. Agric. Int. 2018, 23, 1–11. [Google Scholar] [CrossRef]
- Zhang, S.; Yang, W.; Muneer, M.A.; Ji, Z.; Tong, L.; Zhang, X.; Li, X.; Wang, W.; Zhang, F.; Wu, L. Integrated use of lime with Mg fertilizer significantly improves the pomelo yield, quality, economic returns and soil physicochemical properties under acidic soil of southern China. Sci. Hortic. 2021, 290, 110502. [Google Scholar] [CrossRef]
- Lewis, D.E.; White, J.R.; Wafula, D.; Athar, R.; Dickerson, T.; Williams, H.N.; Chauhan, A. Soil Functional Diversity Analysis of a Bauxite-Mined Restoration Chronosequence. Microb. Ecol. 2010, 59, 710–723. [Google Scholar] [CrossRef]
- Malchair, S.; De Boeck, H.J.; Lemmens, C.M.H.M.; Ceulemans, R.; Merckx, R.; Nijs, I.; Carnol, M. Diversity-function relationship of ammonia-oxidizing bacteria in soils among functional groups of grassland species under climate warming. Appl. Soil Ecol. 2010, 44, 15–23. [Google Scholar] [CrossRef]
- Bai, R.; Wang, J.; Deng, Y.; He, J.; Feng, K.; Zhang, L. Microbial community and functional structure significantly varied among distinct types of paddy soils but responded differently along gradients of soil depth layers. Front. Microbiol. 2017, 8, 945. [Google Scholar] [CrossRef]
- Chen, H.; Ma, K.; Huang, Y.; Yao, Z.; Chu, C. Stable soil microbial functional structure responding to biodiversity loss based on metagenomic evidences. Front. Microbiol. 2021, 12, 716764. [Google Scholar] [CrossRef]
- Fernandez, A.L.; Sheaffer, C.C.; Wyse, D.L.; Staley, C.; Gould, T.J.; Sadowsky, M.J. Associations between soil bacterial community structure and nutrient cycling functions in long-term organic farm soils following cover crop and organic fertilizer amendment. Sci. Total Environ. 2016, 566, 949–959. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Fang, B.; Gao, L.; Li, L.; Wang, S.; Jiang, H.; Li, W. Community structure and ecological functions of soil microorganisms in the degraded area of Barkol Lake. Acta Microbiol. Sin. 2022, 62, 2053–2073. [Google Scholar]
- Fredrickson, J.K.; Balkwill, D.; Drake, G.R.; Romine, M.; Ringelberg, D.; White, D.C. Aromatic-degrading sphingomonas isolates from the deep subsurface. Appl. Environ. Microbiol. 1995, 61, 1917–1922. [Google Scholar] [CrossRef] [Green Version]
- Sul, W.J.; Asuming-Brempong, S.; Wang, Q.; Tourlousse, D.M.; Penton, C.R.; Deng, Y.; Rodrigues, J.L.; Adiku, S.G.; Jones, J.W.; Zhou, J.; et al. Tropical agricultural land management influences on soil microbial communities through its effect on soil organic carbon. Soil Biol. Biochem. 2013, 65, 33–38. [Google Scholar] [CrossRef] [Green Version]
- Kamau, D.M.; Spiertz, J.H.J.; Oenema, O.; Owuor, P.O. Productivity and nitrogen use of tea plantations in relation to age and genotype. Field Crop Res. 2008, 108, 60–70. [Google Scholar] [CrossRef]
- Nemergut, D.R.; Cleveland, C.C.; Wieder, W.R.; Washenberger, C.L.; Townsend, A.R. Plot-scale manipulations of organic matter inputs to soils correlate with shifts in microbial community composition in a lowland tropical rain forest. Soil Biol. Biochem. 2010, 42, 2153–2160. [Google Scholar] [CrossRef]
- Bansal, K.; Kumar, S.; Patil, P.; Sharma, S.; Patil, P. Genomic data resource of type strains of genus Pseudoxanthomonas. Data Brief 2022, 42, 108145. [Google Scholar] [CrossRef] [PubMed]
- Smit, E.; Leeflang, P.; Gommans, S.; Broek, J.V.D.; Mil, S.V.; Wernars, K. Diversity and seasonal fluctuations of the dominant members of the bacterial soil community in a wheat field as determined by cultivation and molecular methods. Appl. Environ. Microb. 2001, 67, 2284–2291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torsvik, V.; Øvreås, L. Microbial diversity and function in soil: From genes to ecosystems. Curr. Opin. Microbiol. 2002, 5, 240–245. [Google Scholar] [CrossRef]
- Waschulin, V.; Borsetto, C.; James, R.; Newsham, K.K.; Donadio, S.; Corre, C.; Wellington, E. Biosynthetic potential of uncultured Antarctic soil bacteria revealed through long read metagenomic sequencing. ISME J. 2021, 16, 101–111. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Depth | Alkali-Hydrolysable N | Available P | Available K | Total N | Total P | Total K | SOM | WCa | WMg |
|---|---|---|---|---|---|---|---|---|---|
| cm | mg kg−1 | % | g kg−1 | cmol kg−1 | |||||
| 0–20 | 105 | 81.9 | 457.67 | 0.17 | 0.1 | 2.03 | 22.8 | 0.31 | 0.12 |
| 20–40 | 90.47 | 19.77 | 133.33 | 0.11 | 0.06 | 1.98 | 18.9 | 0.24 | 0.06 |
| Treatment | SBD | SP | SOM | SFC | SWC | Alkali-Hydrolysable N | Available P | Available K | WCa | WMg | AZn |
|---|---|---|---|---|---|---|---|---|---|---|---|
| g cm3−1 | % | g kg−1 | % | mg kg−1 | cmol kg−1 | ||||||
| CKa | 1.47 a,b | 44.64 b | 18.02 d | 23.24 c | 18.40 c | 81.69 a | 52.41 a | 138.05 c | 0.27 b | 0.10 b | 8.40 d |
| CKb | 1.57 a | 41.01 c | 16.21 d | 23.81 c | 18.94 b,c | 53.04 c | 23.97 c | 115.02 c | 0.18 c | 0.04 d | 6.31 e |
| T1a | 1.35 c | 48.89 a | 27.25 a | 30.46 a | 21.40 a | 81.43 a | 41.39 a,b | 378.04 a | 0.39 a | 0.16 a | 21.40 a |
| T1b | 1.44 b,c | 45.80 b | 24.37 b | 28.60 b | 20.26 a,b | 73.33 b | 29.38 b,c | 258.21 b | 0.27 b | 0.07 c | 16.80 b |
| T2a | 1.41 b,c | 44.70 b | 23.18 b,c | 29.71 a | 19.77 b,c | 69.35 b | 33.94 b,c | 269.78 b | 0.27 b | 0.10 b | 10.30 c |
| T2b | 1.56 a | 40.87 c | 21.83 c | 28.24 b | 18.72 c | 61.90 b,c | 27.58 b,c | 155.91 c | 0.18 c | 0.06 c,d | 8.60 c,d |
| Treatment | Diversity | Richness | |||
|---|---|---|---|---|---|
| Sobs | Shannon | Simpson | ACE | Chao1 | |
| Cka | 3698 c | 6.52 b | 0.0067 a | 4922 c | 4982 c |
| CKb | 3375 d | 6.46 b | 0.0038 c | 4953 c | 4655 c |
| T1a | 4423 a | 6.82 a | 0.0043 b,c | 6058 a | 6045 a |
| T1b | 4385 a | 6.82 a | 0.0042 b,c | 5935 a,b | 5956 a |
| T2a | 4044 b | 6.71 a | 0.0049 b,c | 5459 b,c | 5477 b |
| T2b | 3263 d | 6.50 b | 0.0053 a,b | 5106 c | 4620 c |
| Treatment | Metabolism | Genetic Information Processing | Cellular Processes | Environmental Information Processing | Human Diseases | Organismal Systems | Unclassified |
|---|---|---|---|---|---|---|---|
| Cka | 52.951 a | 15.292 | 3.639 | 14.249 | 0.767 a,b | 0.824 a | 12.278 |
| CKb | 52.457 c | 15.759 | 3.601 | 14.420 | 0.710 c | 0.797 b,c | 12.255 |
| T1a | 52.674 b | 15.379 | 3.534 | 14.410 | 0.798 a | 0.820 a | 12.384 |
| T1b | 52.769 a,b | 15.498 | 3.613 | 14.162 | 0.780 a | 0.819 a | 12.357 |
| T2a | 52.766 a,b | 15.453 | 3.641 | 14.148 | 0.782 a | 0.816 a,b | 12.394 |
| T2b | 52.350 c | 15.768 | 3.620 | 14.418 | 0.727 b,c | 0.789 c | 12.329 |
| Treatment | Amino Acid Metabolism | Translation | Transcription | Metabolism | Metabolism of Other Amino Acids | Infectious Diseases | Neurodegenerative Diseases | Signaling Molecules and Interaction | Cancers | Metabolic Diseases | Cardiovascular Diseases | Circulatory System | Excretory System |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cka | 11.254 a | 4.080 b | 2.405 c | 2.352 a,b | 1.975 a | 0.314 a,b | 0.204 a,b | 0.201 a | 0.106 a | 0.082 b | 0.012 a,b | 0.021 a,b | 0.0445 a,b |
| CKb | 11.091 c | 4.257 a | 2.530 a | 2.289 c | 1.878 b | 0.304 b | 0.176 c | 0.190 b,c | 0.086 b | 0.085 a | 0.007 c | 0.017 c | 0.0453 a |
| T1a | 11.223 a | 4.088 b | 2.433 b,c | 2.367 a | 1.966 a | 0.321 a | 0.219 a | 0.196 a,b | 0.113 a | 0.083 a,b | 0.013 a | 0.022 a | 0.0436 b |
| T1b | 11.201 a,b | 4.139 a,b | 2.444 b,c | 2.353 a,b | 1.945 a,b | 0.321 a | 0.206 a,b | 0.200 a | 0.106 a | 0.083 a,b | 0.011 a,b | 0.021 a | 0.0440 b |
| T2a | 11.194 a,b | 4.124 b | 2.439 b,c | 2.364 a | 1.952 a | 0.324 a | 0.209 a,b | 0.202 a | 0.106 a | 0.082 b | 0.011 a,b | 0.022 a | 0.0436 b |
| T2b | 11.119 b,c | 4.261 a | 2.497 a,b | 2.308 b,c | 1.884 b | 0.309 b | 0.185 b,c | 0.190 c | 0.0907 b | 0.084 a | 0.008 b,c | 0.018 b,c | 0.0452 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, N.; Zhang, W.; Liao, S.; Li, H. Divergent Changes in Bacterial Functionality as Affected by Root-Zone Ecological Restoration in an Aged Peach Orchard. Microorganisms 2022, 10, 2127. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10112127
Sun N, Zhang W, Liao S, Li H. Divergent Changes in Bacterial Functionality as Affected by Root-Zone Ecological Restoration in an Aged Peach Orchard. Microorganisms. 2022; 10(11):2127. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10112127
Chicago/Turabian StyleSun, Na, Weiwei Zhang, Shangqiang Liao, and Hong Li. 2022. "Divergent Changes in Bacterial Functionality as Affected by Root-Zone Ecological Restoration in an Aged Peach Orchard" Microorganisms 10, no. 11: 2127. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10112127