
Rhizobium pusense-Mediated Selenium Nanoparticles–Antibiotics Combinations against Acanthamoeba sp.
 ,
,  and
and
Abstract
:1. Introduction
2. Material and Methods
2.1. Materials
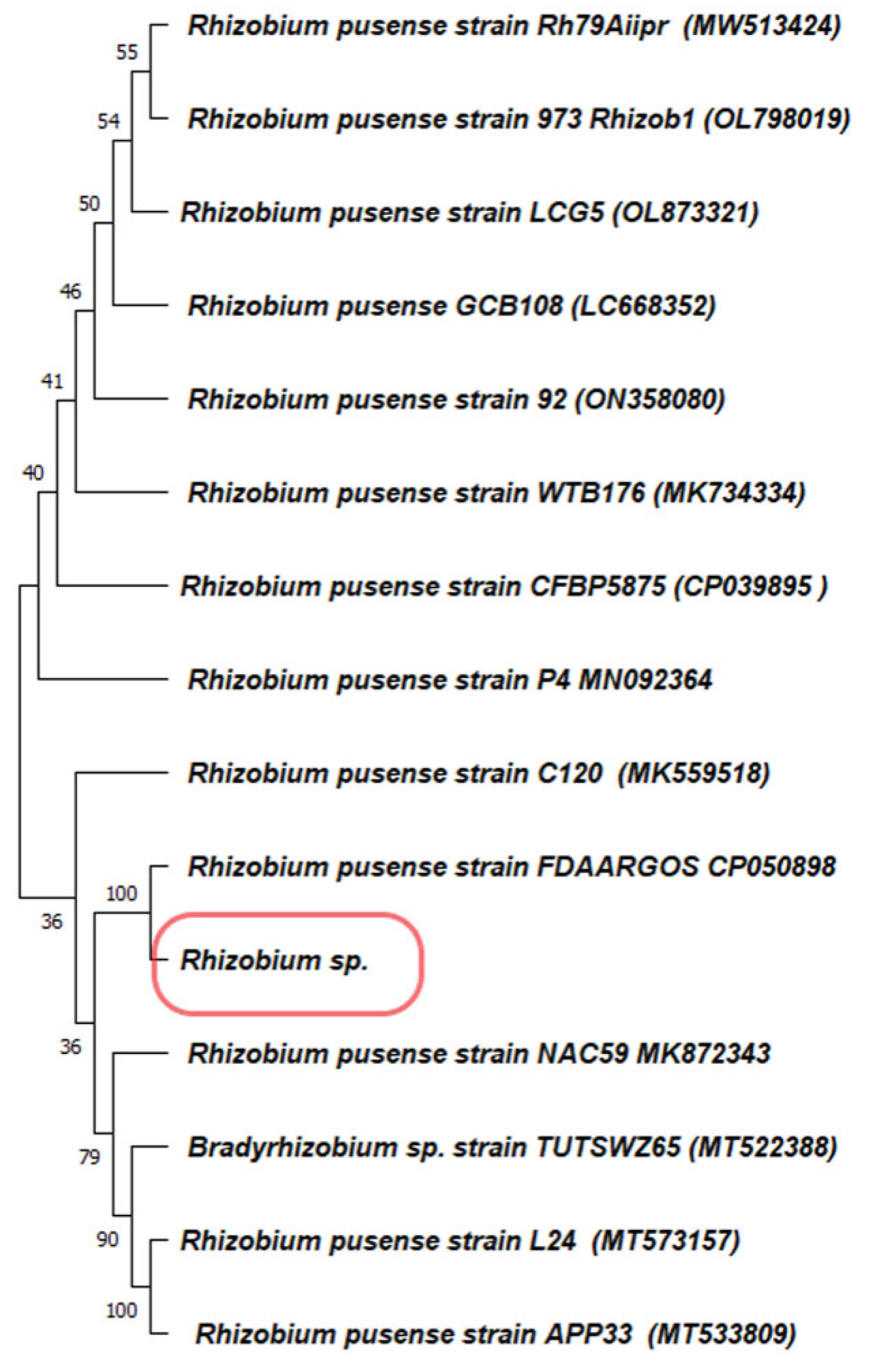
2.2. Sample Collection
2.3. Synthesis of Selenium Nanoparticles (SeNPs)
2.4. Characterization of Selenium Nanoparticles
2.5. Amoebicidal Effect
2.5.1. Test Organisms
2.5.2. Axenization of Acanthamoeba Cysts
2.5.3. Amoebicidal Assay
2.5.4. Combination Study of SeNPs (Se) and Ciprofloxacin (Cipro)
- Cipro-Se: The sub-lethal concentration, i.e., the LC30 of SeNPs, was combined with increasing ciprofloxacin concentrations, i.e., SeNPs were fixed with varying amounts of ciprofloxacin concentration;
- Se-Cipro: The sub-lethal concentration of ciprofloxacin was kept constant, which was calculated previously, and the concentration of SeNPs varied from 1.56 to 50 ppm. Both the combinations were exposed to 50 trophozoites in triplicate for 24 h. The LC50 value for both combinations was determined by Trypan blue exclusion method.
2.5.5. Mode of Action of Ciprofloxacin and SeNPs Combinations
2.5.6. Preparations of Amoeba Cell Lysates
2.5.7. Protease Inhibitor Assay (Trypsin)
2.5.8. Encystment Assay
2.5.9. Excystation Assay
2.5.10. Toxicity Assay of SeNPs–Cipro Conjugates
2.5.11. Statistical Test
3. Results
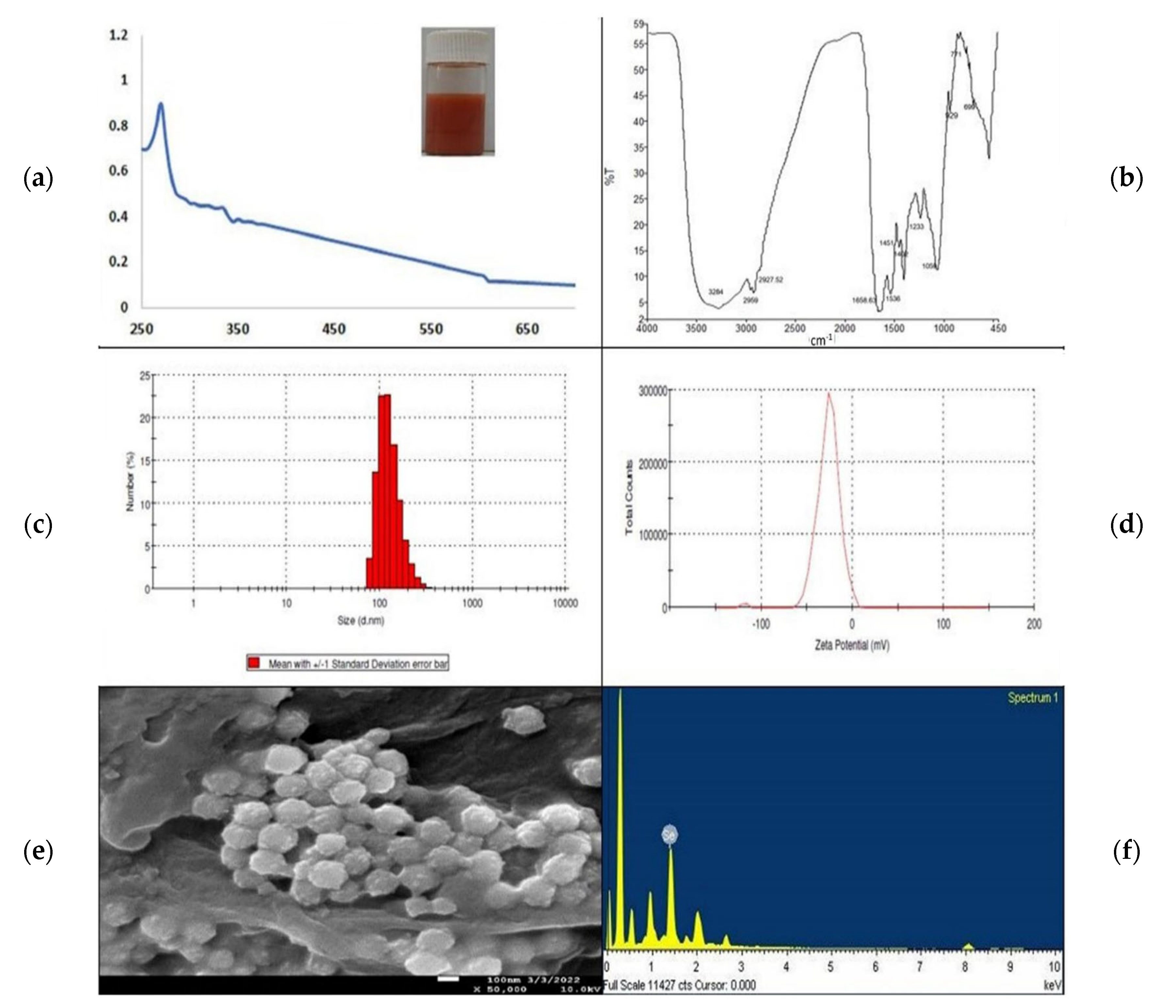
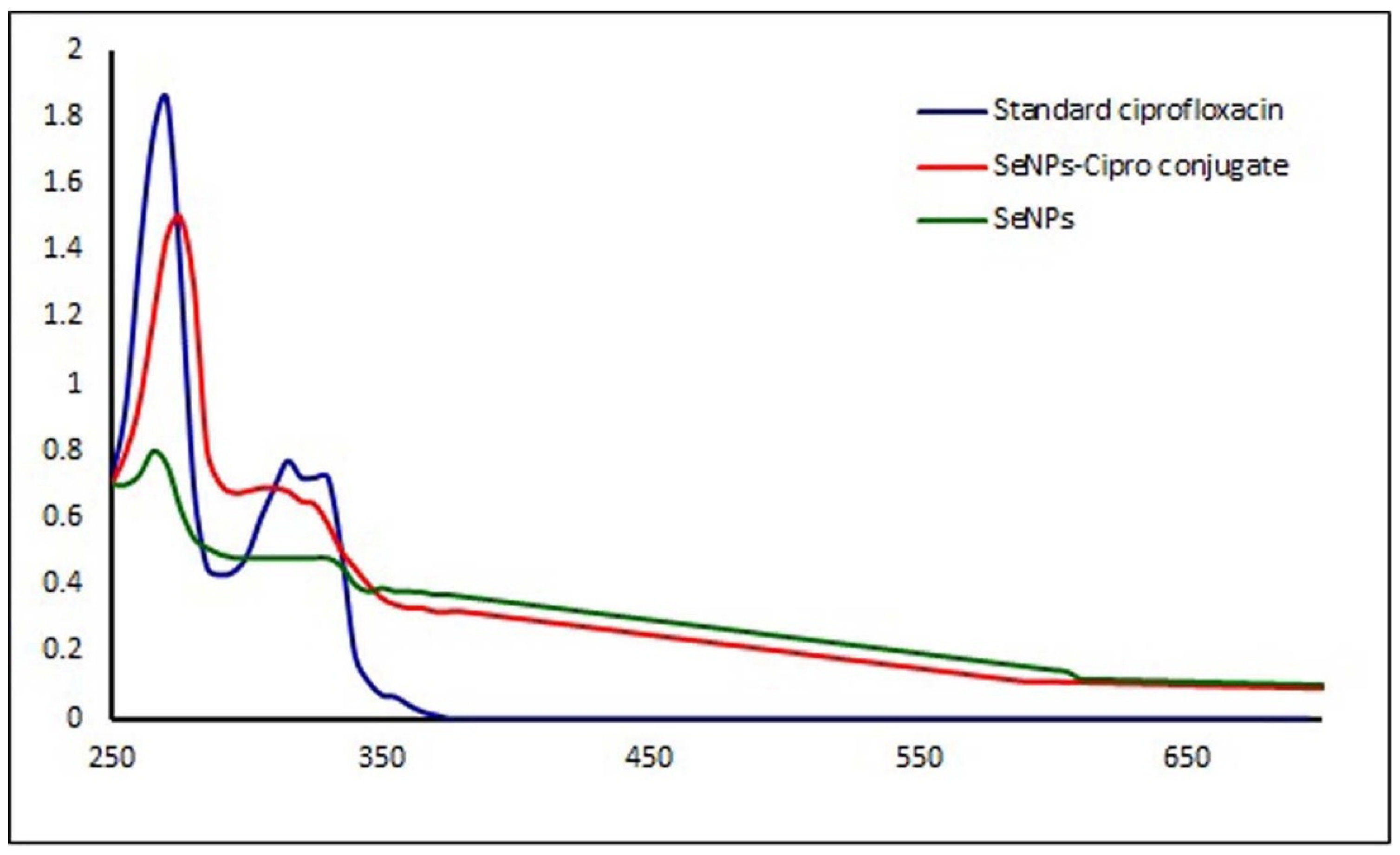
3.1. Synthesis and Characterization of Selenium Nanoparticles
3.2. Anti-Acanthamoeba Activity
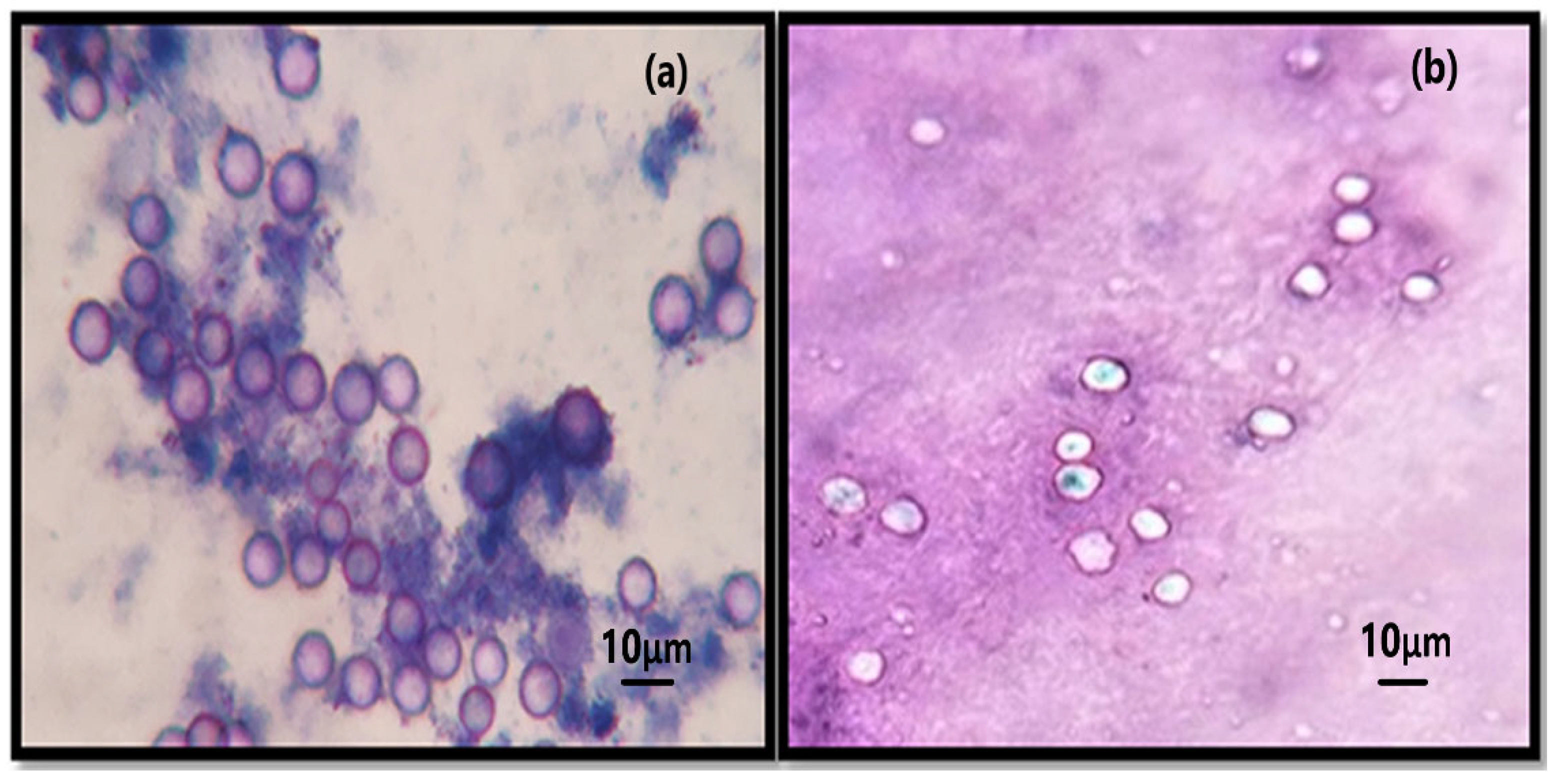
3.3. The Cysticidal Effect of SeNPs and SeNP–Cipro (Se–Cipro)
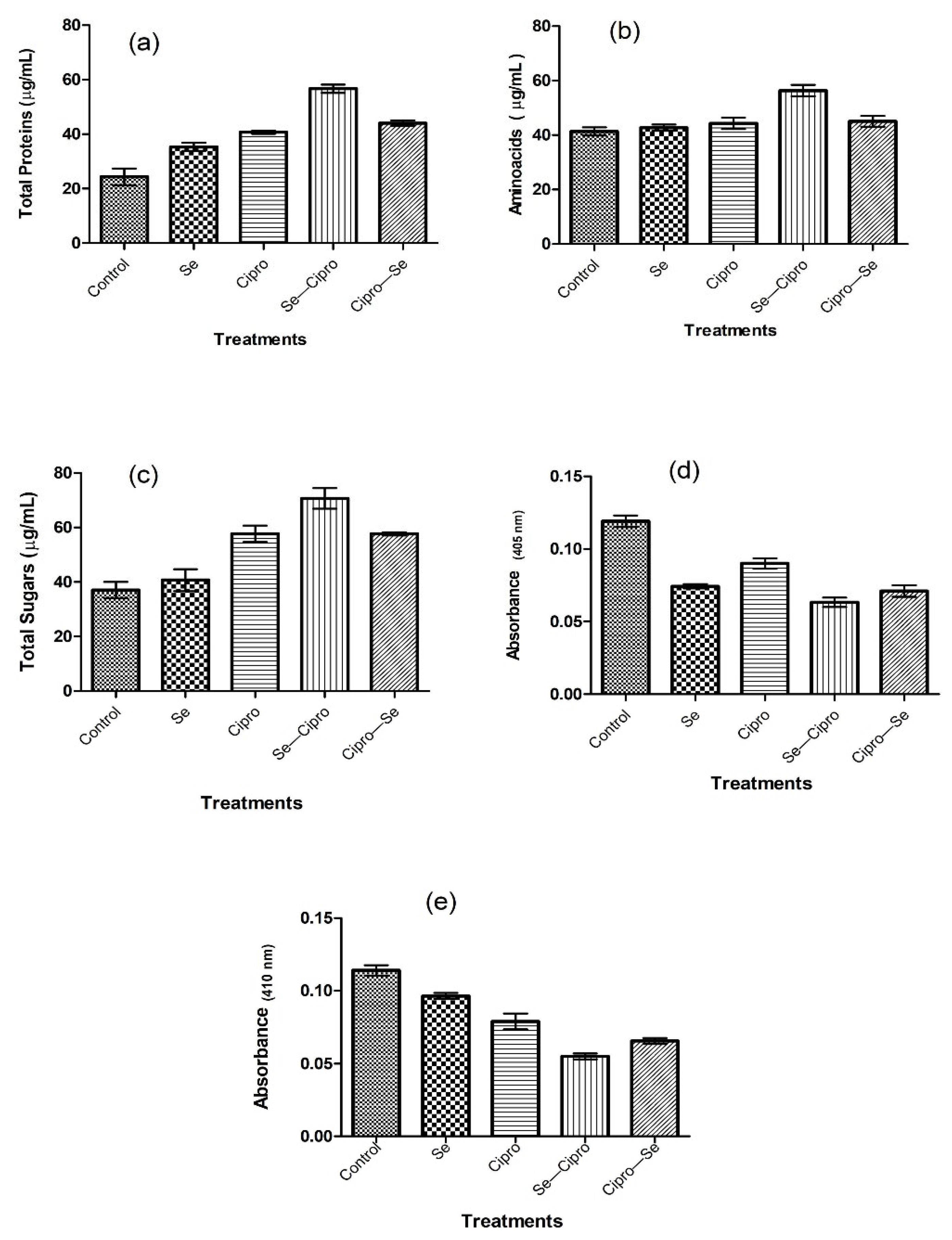
3.4. Mechanism of Anti-Acanthamoeba Activity of SeNPs
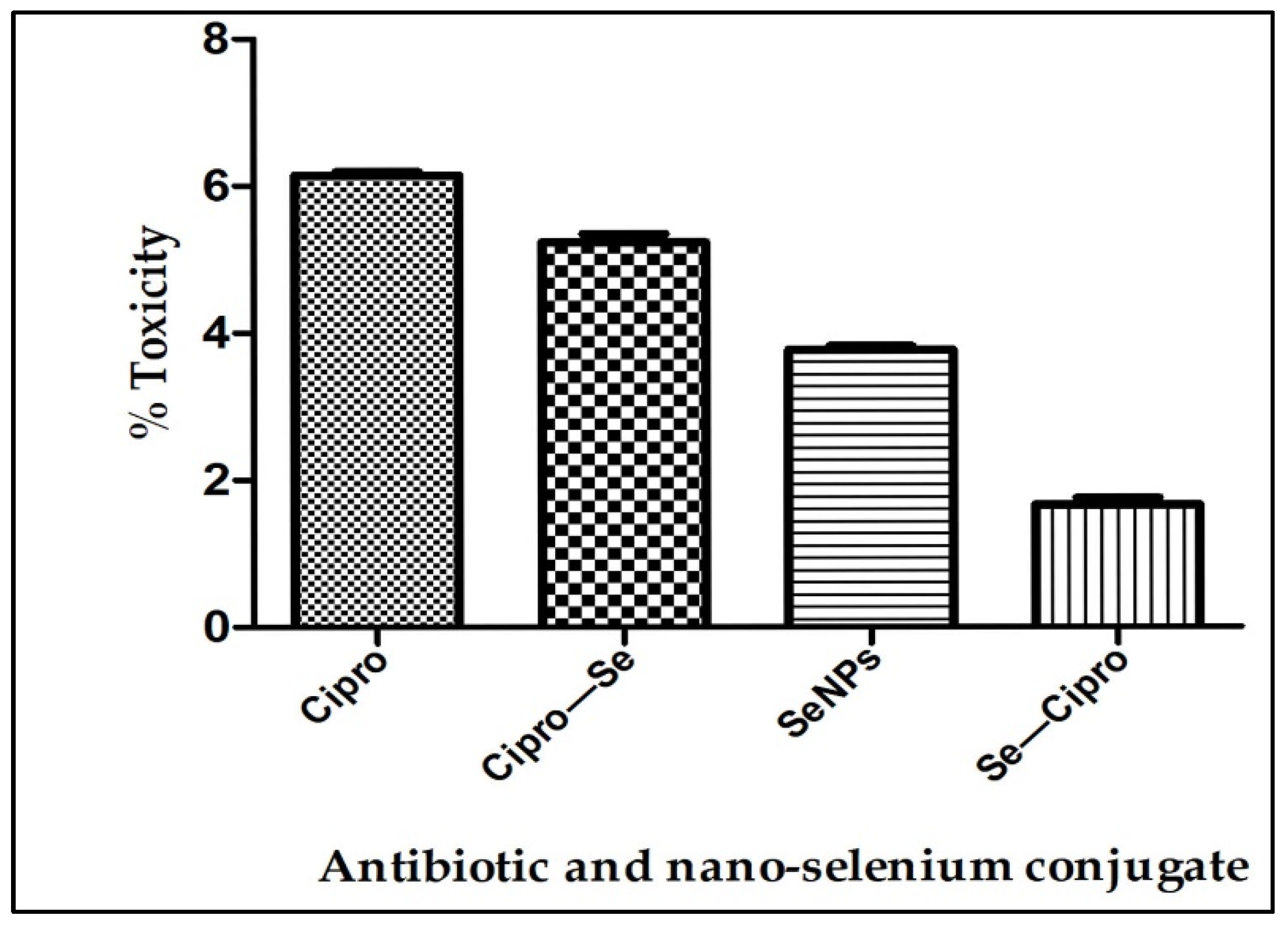
3.5. Toxicity Testing
4. Discussion
5. Conclusions
6. Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Trabelsi, H.; Dendana, F.; Sellami, A.; Sellami, H.; Cheikhrouhou, F.; Neji, S.; Makni, F.; Ayadi, A. Pathogenic free-living amoebae: Epidemiology and clinical review. Pathol. Biol. 2012, 60, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Marciano-Cabral, F.; Cabral, G. Acanthamoeba spp. as agents of disease in humans. Clin. Microbiol. Rev. 2003, 16, 273–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siddiqui, R.; Khan, N.A. Biology and pathogenesis of Acanthamoeba. Parasites Vectors 2012, 5, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gautom, R.K.; Fritsche, T.R.; Lindquist, T.D.; Tasman, W.; Jaeger, E.A. Acanthamoeba keratitis. Duane’s Found. Clin. Ophthalmol. 1997, 2, 1–15. [Google Scholar]
- El-Sayed, N.M.; Ismail, K.A.; Ahmed, S.A.E.G.; Hetta, M.H. In vitro amoebicidal activity of ethanol extracts of Arachis hypogaea L., Curcuma longa L. and Pancratium maritimum L. on Acanthamoeba castellanii cysts. Parasitol. Res. 2012, 110, 1985–1992. [Google Scholar] [CrossRef]
- Khan, N.A. Pathogenesis of Acanthamoeba infections. Microb. Pathog. 2003, 34, 277–285. [Google Scholar] [CrossRef]
- Martín-Navarro, C.M.; Lorenzo-Morales, J.; Cabrera-Serra, M.G.; Rancel, F.; Coronado-Alvarez, N.M.; Pinero, J.E.; Valladares, B. The potential pathogenicity of chlorhexidine-sensitive Acanthamoeba strains isolated from contact lens cases from asymptomatic individuals in Tenerife, Canary Islands, Spain. J. Med. Microbiol. 2008, 57, 1399–1404. [Google Scholar] [CrossRef]
- Schuster, F.L.; Visvesvara, G.S. Opportunistic amoebae: Challenges in prophylaxis and treatment. Drug Resist. Updat. 2004, 7, 41–51. [Google Scholar] [CrossRef]
- Leitsch, D.; Köhsler, M.; Marchetti-Deschmann, M.; Deutsch, A.; Allmaier, G.; Duchêne, M.; Walochnik, J. Major role for cysteine proteases during the early phase of Acanthamoeba castellanii encystment. Eukaryot. Cell. 2010, 9, 611–618. [Google Scholar] [CrossRef] [Green Version]
- Borase, H.P.; Patil, C.D.; Sauter, I.P.; Rott, M.B.; Patil, S.V. Amoebicidal activity of phytosynthesized silver nanoparticles and their in vitro cytotoxicity to human cells. FEMS Microbiollett. 2013, 345, 127–131. [Google Scholar] [CrossRef] [Green Version]
- Anwar, A.; Mungroo, M.R.; Anwar, A.; Sullivan, W.J.; Khan, N.A.; Siddiqui, R. Repositioning of guanabenz in conjugation with gold and silver nanoparticles against pathogenic amoebae Acanthamoeba castellanii and Naegleriafowleri. ACS Infect. Dis. 2019, 5, 2039–2046. [Google Scholar] [CrossRef] [PubMed]
- Hendiger, E.B.; Padzik, M.; Żochowska, A.; Baltaza, W.; Olędzka, G.; Zyskowska, D.; Bluszcz, J.; Jarzynka, S.; Chomicz, L.; Grodzik, M.; et al. Tannic acid-modified silver nanoparticles enhance the anti-Acanthamoeba activity of three multipurpose contact lens solutions without increasing their cytotoxicity. Parasites Vectors 2020, 13, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Hendiger, E.B.; Padzik, M.; Sifaoui, I.; Reyes-Batlle, M.; López-Arencibia, A.; Zyskowska, D.; Grodzik, M.; Pietruczuk-Padzik, A.; Hendiger, J.; Olędzka, G.; et al. Silver Nanoparticles Conjugated with Contact Lens Solutions May Reduce the Risk of Acanthamoeba Keratitis. Pathogens 2021, 10, 583. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.; Li, Y.; Tjong, S.C. Bactericidal and cytotoxic properties of silver nanoparticles. Int. J. Mol. Sci. 2019, 20, 449. [Google Scholar] [CrossRef] [Green Version]
- Ferdous, Z.; Nemmar, A. Health impact of silver nanoparticles: A review of the bio distribution and toxicity following various routes of exposure. Int. J. Mol. Sci. 2020, 21, 2375. [Google Scholar] [CrossRef] [Green Version]
- Ortillés, A.; Belloc, J.; Rubio, E.; Fernández, M.T.; Benito, M.; Cristóbal, J.A.; Calvo, B.; Goñi, P.P. In-vitro development of an effective treatment for Acanthamoeba keratitis. Int. J. Antimicrob. Agents 2017, 50, 325–333. [Google Scholar] [CrossRef]
- Eszenyi, P.; Sztrik, A.; Babka, B.; Prokisch, J. Elemental, nano-sized (100–500 nm) selenium production by probiotic lactic acid bacteria. Int. J. Biosci. Biochem. Bioinform. 2011, 1, 148. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Guo, Y.; Qiao, L.; Ma, L.; Cheng, Y.; Roman, A. Biogenic synthesis of novel functionalized selenium nanoparticles by Lactobacillus casei ATCC 393 and its protective effects on intestinal barrier dysfunction caused by enterotoxigenic Escherichia coli K88. Front. Microbiol. 2018, 9, 1129. [Google Scholar] [CrossRef] [Green Version]
- Maiyo, F.; Singh, M. Selenium nanoparticles: Potential in cancer gene and drug delivery. Nanomedicine 2017, 12, 1075–1089. [Google Scholar] [CrossRef]
- Khurana, A.; Tekula, S.; Saifi, M.A.; Venkatesh, P.; Godugu, C. Therapeutic applications of selenium nanoparticles. Biomed. Pharmacother. 2019, 111, 802–812. [Google Scholar] [CrossRef]
- Basaglia, M.; Toffanin, A.; Baldan, E.; Bottegal, M.; Shapleigh, J.P.; Casella, S. Selenite-reducing capacity of the copper-containing nitrite reductase of Rhizobium sullae. FEMS Microbiollett. 2007, 269, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Al Thaher, Y.; Chandrasekaran, B.; Panchu, S.J. The Importance of Nano-materials Characterization Techniques. In Integrative Nanomedicine for New Therapies; Engineering Materials; Krishnan, A., Chuturgoon, A., Eds.; Springer: Cham, Switzerland, 2020; pp. 19–37. [Google Scholar]
- Makdoumi, K.; Bäckman, A.; Mortensen, J.; Magnuson, A.; Crafoord, S. Comparison of UVA-and UVA/riboflavin-induced growth inhibition of Acanthamoeba castellanii. Graefes. Arch. Clin. Exp. Ophthalmol. 2013, 251, 509–514. [Google Scholar] [CrossRef] [PubMed]
- Lakhundi, S.; Khan, N.A.; Siddiqui, R. Inefficacy of marketed contact lens disinfection solutions against keratitis-causing Acanthamoeba castellanii belonging to the T4 genotype. Exp. Parasitol. 2014, 141, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Anwar, A.; Siddiqui, R.; Hussain, M.A.; Ahmed, D.; Shah, M.R.; Khan, N.A. Silver nanoparticle conjugation affects anti-acanthamoebic activities of amphotericin B, nystatin, and fluconazole. Parasitol. Res. 2018, 117, 265–271. [Google Scholar] [CrossRef]
- Anwar, A.; Ting, E.L.S.; Anwar, A.; ulAin, N.; Faizi, S.; Shah, M.R.; Khan, N.A.; Siddiqui, R. Antiamoebic activity of plant-based natural products and their conjugated silver nanoparticles against Acanthamoeba castellanii (ATCC 50492). AMB Express 2020, 10, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Dudley, R.; Jarroll, E.L.; Khan, N.A. Carbohydrate analysis of Acanthamoeba castellanii. Exp. Parasitol. 2009, 122, 338–343. [Google Scholar] [CrossRef]
- Vijayakumar, S.; Ganesan, S. In vitro cytotoxicity assay on gold nanoparticles with different stabilizing agents. J. Nanomater. 2012, 2012, 14. [Google Scholar] [CrossRef] [Green Version]
- Asghari-Paskiabi, F.; Imani, M.; Razzaghi-Abyaneh, M.; Rafii-Tabar, H. Fusariumoxysporum a bio-factory for nano selenium compounds: Synthesis and characterization. Sci. Iran 2018, 25, 1857–1863. [Google Scholar]
- Menon, S.; Agarwal, H.; Shanmugam, V.K. Catalytical degradation of industrial dyes using biosynthesized selenium nanoparticles and evaluating its antimicrobial activities. Sustain. Environ. Res. 2021, 31, 1–12. [Google Scholar] [CrossRef]
- Gunti, L.; Dass, R.S.; Kalagatur, N.K. Phytofabrication of selenium nanoparticles from Emblicaofficinalis fruit extract and exploring its biopotential applications: Antioxidant, antimicrobial, and biocompatibility. Front. Microbiol. 2019, 10, 931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murugesan, G.; Nagaraj, K.; Sunmathi, D.; Subramani, K. Methods involved in the synthesis of selenium nanoparticles and their different applications—A review. Eur. J. Biomed. 2019, 6, 189–194. [Google Scholar]
- Rogachev, A.A.; Yarmolenko, M.A.; Rogachou, A.V.; Tapalski, D.V.; Liu, X.; Gorbachev, D.L. Morphology and structure of antibacterial nanocomposite organic–polymer and metal–polymer coatings deposited from active gas phase. RSC Adv. 2013, 3, 11226–11233. [Google Scholar] [CrossRef]
- Saha, B.; Bhattacharya, J.; Mukherjee, A.; Ghosh, A.; Santra, C.; Dasgupta, A.K.; Karmakar, P. In vitro structural and functional 506 evaluation of gold nanoparticles conjugated antibiotics. Nanoscale Res. Lett. 2007, 2, 614–622. [Google Scholar] [CrossRef] [Green Version]
- Strober, W. Trypan blue exclusion test of cell viability. Curr. Protoc. Immunol. 1997, 21, A–3B. [Google Scholar]
- Khunkitti, W.; Hann, A.C.; Lloyd, D.; Furr, J.R.; Russell, A.D. Biguanide-induced changes in Acanthamoeba castellanii: An electron microscopic study. J. Appl. Microbiol. 1998, 84, 53–62. [Google Scholar] [CrossRef]
- McDonnell, G.; Russell, A.D. Antiseptics and disinfectants: Activity, action, and resistance. Clin. Microbiol. Rev. 1999, 12, 147–179. [Google Scholar] [CrossRef] [Green Version]
- Anwar, A.; Khan, N.A.; Siddiqui, R. Combating Acanthamoeba spp. cysts: What are the options? Parasites Vectors 2018, 11, 26. [Google Scholar] [CrossRef] [Green Version]
- Lorenzo-Morales, J.; Martín-Navarro, C.M.; López-Arencibia, A.; Arnalich-Montiel, F.; Piñero, J.E.; Valladares, B. Acanthamoeba keratitis: An emerging disease gathering importance worldwide? Trends. Parasitol. 2013, 29, 181–187. [Google Scholar] [CrossRef]
- Sharma, G.; Kalra, S.K.; Tejan, N.; Ghoshal, U. Nanoparticles based therapeutic efficacy against Acanthamoeba: Updates and future prospect. Exp. Parasitol. 2020, 218, 108008. [Google Scholar] [CrossRef]
- Ferro, C.; Florindo, H.F.; Santos, H.A. Selenium nanoparticles for biomedical applications: From development and characterization to therapeutics. Adv. Healthc. Mater. 2021, 10, 2100598. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Zhang, J.; Xu, J.F.; Pi, J. The advancing of selenium nanoparticles against infectious diseases. Front. Pharmacol. 2021, 12, 1971. [Google Scholar] [CrossRef] [PubMed]
- Avendaño, R.; Chaves, N.; Fuentes, P.; Sánchez, E.; Jiménez, J.I.; Chavarría Vargas, M. Production of selenium nanoparticles in Pseudomonas putida KT2440. Sci. Rep. 2016, 6, 37155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shubharani, R.; Mahesh, M.; Yogananda Murthy, V. Biosynthesis and characterization, antioxidant and antimicrobial activities of selenium nanoparticles from ethanol extract of Bee Propolis. J. Nanomed. Nanotechnol. 2019, 10, 2. [Google Scholar]
- Vahdati, M.; Tohidi Moghadam, T. Synthesis and Characterization of Selenium Nanoparticles-Lysozyme Nanohybrid System with Synergistic Antibacterial Properties. Sci. Rep. 2020, 10, 510. [Google Scholar] [CrossRef]
- Nandiyanto, A.B.D.; Oktiani, R.; Ragadhita, R. How to read and interpret FTIR spectroscope of organic material. Indones. J. Sci. Technol. 2019, 4, 97–118. [Google Scholar] [CrossRef] [Green Version]
- Seal, D.V. Acanthamoeba keratitis update—Incidence, molecular epidemiology and new drugs for treatment. Eye 2003, 17, 893–905. [Google Scholar] [CrossRef] [Green Version]
- Niyyati, M.; Sasani, R.; Mohebali, M.; Ghazikhansari, M.; Kargar, F.; Hajialilo, E.; Rezaeian, M. Anti-Acanthamoeba effects of silver and gold nanoparticles and contact lenses disinfection solutions. Iran J. Parasitol. 2018, 13, 180. [Google Scholar]
- Abjani, F.; Khan, N.A.; Jung, S.Y.; Siddiqui, R. Status of the effectiveness of contact lens disinfectants in Malaysia against keratitis-causing pathogens. Exp Parasitol. 2017, 183, 187–193. [Google Scholar] [CrossRef]
- Siddiqui, R.; Lakhundi, S.; Khan, N.A. Status of the effectiveness of contact lens solutions against keratitis-causing pathogens. Cont. Lens Anterior Eye 2015, 38, 34–38. [Google Scholar] [CrossRef]
- Salunkhe, J.D.; Mohite, B.V.; Patil, S.V. Naringenin biosynthesis and fabrication of naringenin mediated nano silver conjugate for antimicrobial potential. Nat. Prod. Res. 2022, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Padzik, M.; Hendiger, E.B.; Chomicz, L.; Grodzik, M.; Szmidt, M.; Grobelny, J.; Lorenzo-Morales, J. Tannic acid-modified silver nanoparticles as a novel therapeutic agent against Acanthamoeba. Parasitol. Res. 2018, 117, 3519–3525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aqeel, Y.; Siddiqui, R.; Anwar, A.; Shah, M.R.; Khan, N.A. Nanoparticle conjugation enhances the antiacanthamoebic effects of chlorhexidine. Antimicrob. Agents Chemother. 2016, 60, 1283–1288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xuguang, S.; Lin, C.; Yan, Z.; Zhiqun, W.; Ran, L.; Shiyun, L.; Xiuying, J. Acanthamoeba keratitis as a complication of orthokeratology. Am. J. Ophthalmol. 2003, 136, 1159–1161. [Google Scholar] [CrossRef] [PubMed]
- Henrissat, B.; Callebaut, I.; Fabrega, S.; Lehn, P.; Mornon, J.P.; Davies, G. Conserved catalytic machinery and the prediction of a common fold for several families of glycosyl hydrolases. Proc. Nat. Acad. Sci. USA 1995, 92, 7090–7094. [Google Scholar] [CrossRef] [Green Version]
- Hadas, E.; Mazur, T. Proteolytic enzymes of pathogenic and non-pathogenic strains of Acanthamoeba spp. Trop. Med. Parasitol. Off. Organ Dtsch. Trop. Ges. Dtsch. Ges. Tech. (GTZ) 1993, 44, 197–200. [Google Scholar]
- Mitro, K.; Bhagavathiammai, A.; Zhou, O.M.; Bobbett, G.; McKerrow, J.H.; Chokshi, R.; Chokshi, B.; James, E.R. Partial characterization of the proteolytic secretions of Acanthamoeba polyphaga. Exp. Parasitol. 1994, 78, 377–385. [Google Scholar] [CrossRef]
- Khan, N.A.; Jarroll, E.L.; Panjwani, N.; Cao, Z.; Paget, T.A. Proteases as markers for differentiation of pathogenic and nonpathogenic species of Acanthamoeba. J. Clin. Microbiol. 2000, 38, 2858–2861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrante, A.; Bates, E.J. Elastase in the pathogenic free-living amoebae Naegleria and Acanthamoeba spp. Infect. Immun. 1998, 56, 3320–3321. [Google Scholar] [CrossRef] [Green Version]
- Leher, H.; Silvany, R.; Alizadeh, H.; Huang, J.; Niederkorn, J.Y. Mannose induces the release of cytopathic factors from Acanthamoeba castellanii. Infect. Immun. 1998, 66, 5–10. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Test Species | Test Products | LC50 ± SD (mg lit–1) | 95% Fiducial Limits | LC90 ± SD (mg lit–1) | 95% Fiducial Limits | Regression Equation |
|---|---|---|---|---|---|---|
| Acanthamoeba sp. | SeNPs | 72.084 ± 10.87 | 56.593–106.58 | 115.058 ± 19.264 | 87.7664–176.491 | Y = 0.423 + 0.149X |
| Cipro | 56.2608 ± 2.2078 | 51.7279–60.43 | 125.0330 ± 3.13 | 119.2960–131.69 | Y = 9.27 + 0.274 X | |
| Cipro–Se | 53.9078 ± 5.4802 | 45.4017–68.6011 | 94.1228 ± 11.056 | 77.1133–124.0217 | Y = 1.55 + 0.302 X | |
| Se–Cipro | 37.452 ± 3.31 | 32.196–46.153 | 94.122 ± 11.05 | 77.113–124.021 | Y= 4.78 + 0.980 X | |
| Sodium selenite | 120.143± 2.56 | 115.316- 125.444 | 205.1626± 6.389 | 193.82–219.166 | Y= 8.53 + 0.960 X | |
| Chlorohexidine | 24.580 ± 1.25 | 22.361- 27.36 | 56.070 ± 3.61 | 49.95–64.52 | Y= 6.52 + 0.754 X |
| Sr. No. | Concentrations ppm | Total No. of Cysts | No. Cyst After 24 hrs. | % Encystment | % Excystment |
|---|---|---|---|---|---|
| 1 | 56.260 Cipro | 50 | 5 | 10.66 | Nil |
| 2 | 72.084 Se | 50 | 10 | 22.066 | Nil |
| 3 | 53.907 Cipro–Se | 50 | 3 | 7.33 | Nil |
| 4 | 37.452 Se–Cipro | 50 | 2 | 4.66 | Nil |
| 5 | Control | 50 | 45 | 39.33 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nikam, P.B.; Salunkhe, J.D.; Marathe, K.R.; Alghuthaymi, M.A.; Abd-Elsalam, K.A.; Patil, S.V. Rhizobium pusense-Mediated Selenium Nanoparticles–Antibiotics Combinations against Acanthamoeba sp. Microorganisms 2022, 10, 2502. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10122502
Nikam PB, Salunkhe JD, Marathe KR, Alghuthaymi MA, Abd-Elsalam KA, Patil SV. Rhizobium pusense-Mediated Selenium Nanoparticles–Antibiotics Combinations against Acanthamoeba sp. Microorganisms. 2022; 10(12):2502. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10122502
Chicago/Turabian StyleNikam, Pradnya B., Jitendra D. Salunkhe, Kiran R. Marathe, Mousa A. Alghuthaymi, Kamel A. Abd-Elsalam, and Satish V. Patil. 2022. "Rhizobium pusense-Mediated Selenium Nanoparticles–Antibiotics Combinations against Acanthamoeba sp." Microorganisms 10, no. 12: 2502. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10122502