
Screening and Application of Ligninolytic Microbial Consortia to Enhance Aerobic Degradation of Solid Digestate

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Initial Environments
2.2. LAMACs—Enrichment and Selection
- (i)
- In R1 reactors, aerobic enrichment was carried out under a liquid state. Initial environments were diluted with the enrichment solution (ratio 1/10 w/w) to favor the development of lignin-degrading microorganisms that would be able to thrive in liquid [12]. An HRT of 10 days was set to avoid any preselection in ligninolytic microorganisms based on their growth rate. Every day, 20 mL of fresh enrichment solution was pumped into R1, and 20 mL of “enriched solution” was pumped out. Of this volume, 10 mL was pumped out the system and discarded, whereas from the remaining volume, 5 mL was pumped into each of the reactors R2 and R3.
- (ii)
- In R2 and R3 reactors, aerobic lignin-degrading bacteria were selected under a solid state. Wheat straw and chestnut wood were only partially emerged as the liquid content was set to 50 mL. Enriched liquids, from R1 reactors, before being injected in R2 and R3, were ten times diluted with M9 media (45 mL supplied by another peristaltic pump). The HRT in this vessel was 1 day, as 50 mL of liquid was pumped out and discarded every day. The aim of all these conditions was to select microorganisms, coming from liquid media, that are able to fix themselves quickly and irreversibly (to avoid washout due to the short HRT) to wood and straw, and use them as the main carbon source (dilution aims to reduce Kraft lignin concentration). Finally, these reactors were shaken manually once a week to ensure that all wood and straw were in contact with the pumped-in liquid.
2.3. Consortia Propagation
2.3.1. Preparation of the Carbon Source and Solutions
2.3.2. Transfer Procedure
2.3.3. Propagation in Liquid Phase
- (i)
- Two Erlenmeyer flasks where only sterile complex SD was used and no source of microorganisms was added. These were the negative controls and aimed to check the sterility of the complex SD used as the carbon source in this propagation experiment. These are referred to as “Sterility-check” samples.
- (ii)
- Erlenmeyer flasks where the source of microorganisms was fresh SD. Practically, fresh SD was washed with sterile PBS (2 g of TS in 50 mL of PBS). Then this liquid was poured in the same proportion (25 mL) as previously in three flasks containing sterile complex SD and minimal mineral media. These were positive controls that allowed measurement of the endogenous activity of microorganisms of SD during this propagation experiment. These are referred to as “Dig” samples.
- (iii)
- Erlenmeyer flasks where the source of microorganisms was fresh dry compost fertilizer (CF). Practically, fresh CF was washed with sterile PBS (2 g of TS in 50 mL of PBS). Then, this liquid was poured in the same proportion (25 mL) as previously in three flasks containing sterile complex SD and minimal mineral media. These were controls that allowed measurement of the activity of microorganisms from CF without any enrichment and selection steps using LAMACs. These are referred to as “Direct-CF” samples.
- (iv)
- Erlenmeyer flasks where the source of microorganisms was a quick selection of endogenous SD microorganisms able to degrade Kraft lignin. Practically, fresh SD was washed with sterile PBS (2 g of TS in 50 mL of PBS). Then, this liquid was poured under sterile conditions into a dozen Petri dishes containing M9 medium complemented with Kraft lignin at 1 g∙L−1 and Agar at 15 g∙L−1. After one week at 30 °C, the biggest colonies were harvested and mixed directly into the minimal mineral medium. It was then supplemented in three flasks with sterile complex SD. These were positive controls that allowed the measurement of the activity of endogenous microorganisms of SD, quickly selected on Petri dishes (instead of the LAMACs system), after their culture in Erlenmeyer flasks. These are referred to as “Petri” samples.
2.4. Addition of a Consortia Solution to Enhance Short-Term Aerobic Post-Treatment
2.5. Microbial Community Analysis
- (i)
- Liquid samples were centrifuged. Supernatants were eliminated and remaining pellets were further stored at −20 °C before use.
- (ii)
- For solid sampling, they were stored in 2 mL Eppendorf tubes (generally between 0.1 and 0.5 g of solid samples) at −20 °C before genomic DNA extraction.
- (i)
- The six initial environments were sampled before their use in R1 reactors of LAMACs. For F1, F2, F3, CF and RS, this corresponded to a solid sampling, whereas for RU it was a liquid sampling.
- (ii)
- At the end of the LAMACs step, liquid sampling was performed on the twelve solutions obtained after PBS solid washing of wheat straw and chestnut wood, for each of the six initial environments. Liquid solutions were also sampled following the PBS washing step of solids used for the additional conditions tested (Dig, Direct-CF, Petri). In addition, the Sterility-check sample consisted of a solid sample of sterilized SD.
- (iii)
- Finally, at the end of the Erlenmeyer step, liquid sampling was carried out on all Erlenmeyer flasks that were used as inoculate for the short-term aerobic post-treatment.
3. Results
3.1. Alpha Diversity Analysis
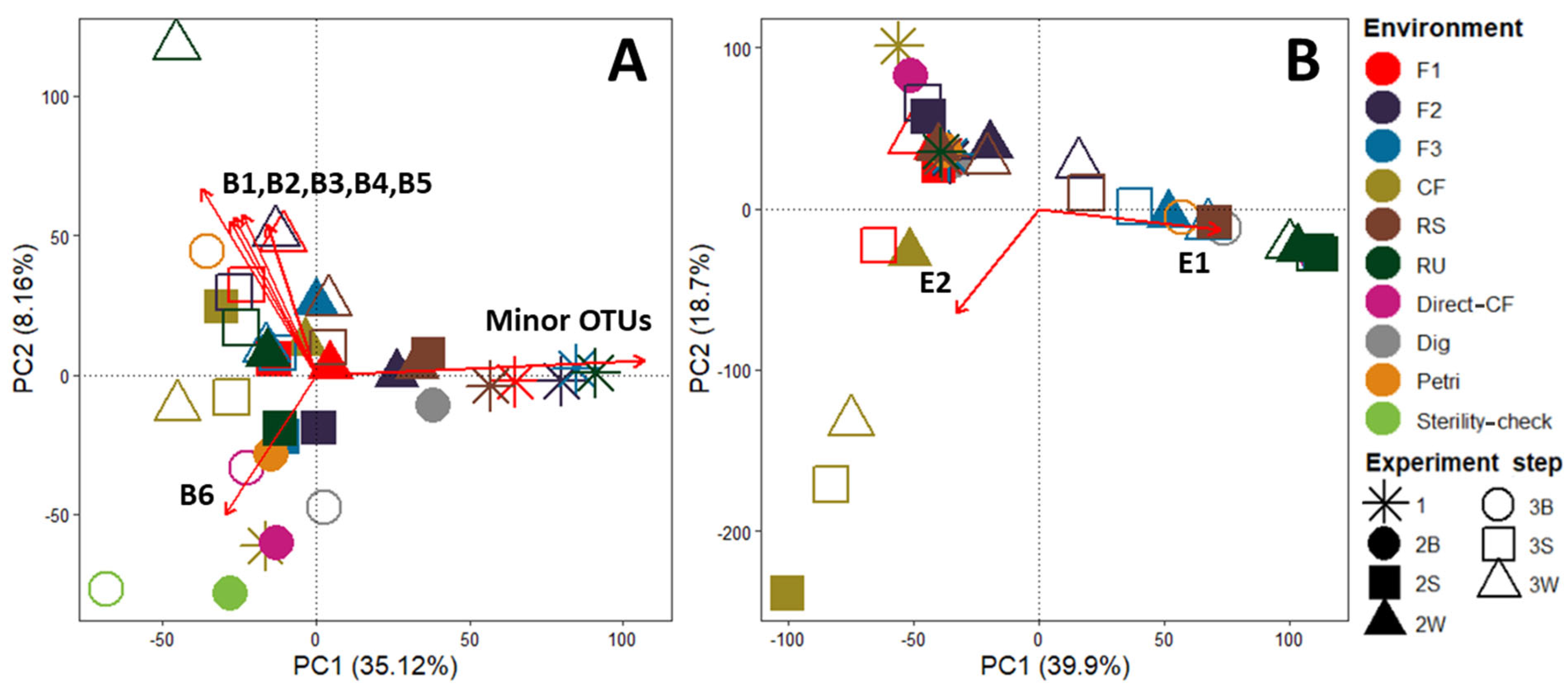
3.2. Beta Diversity Analysis
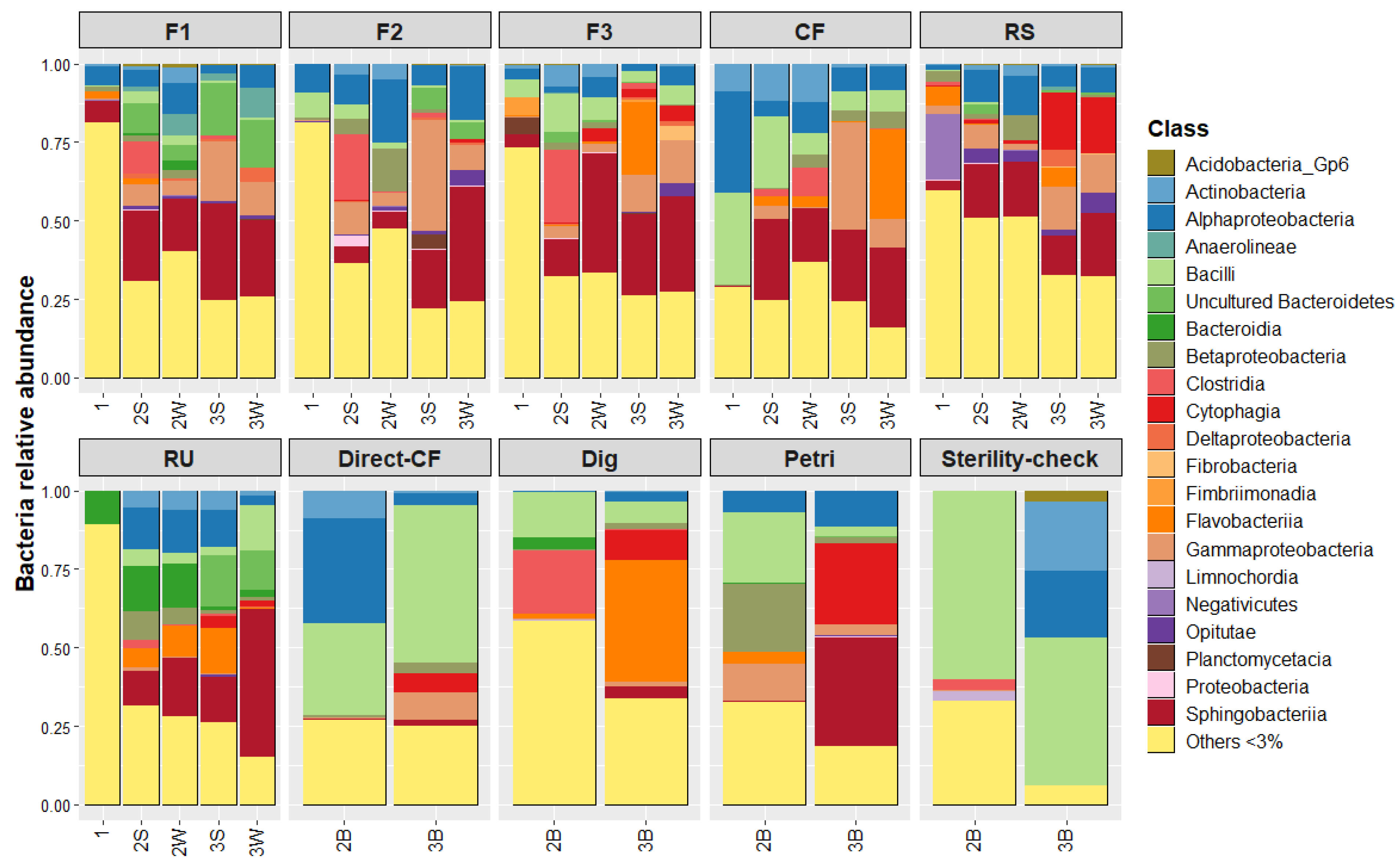
3.3. Microbial Communities: Composition and Quantity over Experiment Steps
3.4. Evaluation of the Impact of Consortia Addition on Efficiency of Short-Term Aerobic Post-Treatment—Ecosystem Function Relationship
4. Discussion
- (i)
- Microbial lignin degradation requires a multiplicity of oxidative enzymes and heterogeneous small molecule co-factors that are produced by ligninolytic fungi and bacteria [23]. Currently, lignin biodegradation in nature is thought to occur in two main stages that consist of its depolymerization followed by the mineralization of resultant heterogeneous aromatics [24]. Although bacteria are reported to be dominant and the most active in the mineralization step, this is not the case for the depolymerization step [25]. Indeed, filamentous Basidiomycetes white-rot fungi were identified as major actors in this step due to their capacity to produce high quantities of various oxidative enzymes (e.g., laccase and lignin peroxidase) [26]. In comparison to fungi, the identified ligninolytic bacteria (e.g., Pseudomonas sp., Rhodococcus sp.) have significantly lower activities during the depolymerization step [24]. For this study, depolymerization of the SD lignin-like fraction is sought because aromatic units can be converted to methane during anaerobic digestion [27]. Assuming that most depolymerization activity is due to dominant microbial species, it is interesting to look at those that were enriched through the experiment steps. For Sordariomycetes fungi and, in general, for Ascomycete, it is reported that the lignin depolymerization rate is slower than for white-rot fungi due to the difference in enzymatic systems (e.g., lack of ligninolytic Class II enzymes) [28]. More generally, fungi obtained after the propagation steps were mostly Ascomycetes. Basidiomycetes that were notably present in the initial forest environments disappeared after the LAMACs step. One hypothesis is that the liquid state during enrichment with Kraft lignin (R1) favored the growth of Ascomycetes over Basidiomycetes. Indeed, Basidiomycetes, due to their filamentous nature, are reported to grow better during solid state fermentation [29]. For bacteria, Paenibacillus sp. used in a consortium treating pulp and paper wastewater were reported to be able to degrade and metabolize the higher molecular weight lignin molecules [30]. However, with the exceptions of RU, Direct-CF and Sterility-check, their abundances were relatively low. Finally, Chitinophagia, the most significantly enriched bacteria, are not ligninolytic strains. Instead, their enzymatic activities are directed toward carbohydrate degradation (e.g., deconstruction of dead fungal material via chitin hydrolysis) [20,31]. Thus, dominant fungi and bacteria that were enriched through this experiment are not reported as efficient actors in lignin depolymerization, notably in comparison to white-rot fungi.
- (ii)
- The designed approach to obtain consortia was unconventional in comparison to other existing studies focusing on screening of microbial ligninolytic consortia. Successive transfer with Erlenmeyer flasks or sequential batch reactors containing the targeted final lignin rich substrate are often preferred [32,33,34]. With these kinds of cultivation strategies, it is possible, notably during transfer steps, to perform precise monitoring of the ligninolytic activity (microbial sampling, enzymatic activity, percentage of degraded lignin, etc.). However, with LAMACs, a system that operates in a continuous mode, reactors were closed and such activity monitoring of solids was not possible. It was thus not possible to determine during the experiment if long-term LAMACs operations were sufficient to obtain reduced, stable and ligninolytic-active microbial communities. Afterwards, it appeared that it was not the case and earlier monitoring would have been beneficial to further select consortia.
- (iii)
- From our knowledge, studies on microbial ligninolytic consortia use either classical lignocellulosic biomasses, such as straw or wood, or industrial lignin derivatives [29,32]. In both cases, cellulose, hemicellulose and lignin are the main molecules present. In comparison to these lignin-rich substrates, SD composition is more diverse and contains higher quantities of proteins, sugars, and lipids, which may provide an opportunity for microbial activities other than ligninolytic ones [35]. Thus, the realization of specific ligninolytic activities is even more challenging because consortia may reorient their activity towards easier-to-degrade fractions. It can be hypothesized that, during the short-term aerobic post-treatment, OTUs present in applied consortia and able to efficiently metabolize proteins, sugars or lipids, may have outcompeted ligninolytic species with slower metabolisms (such as Sordariomycetes or Paenibacillus). It would have been interesting to test the obtained consortia on more classical substrates (straw, wood) to only evaluate their ligninolytic activities.
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IEA. Outlook for Biogas and Biomethane. Prospects for Organic Growth; World Energy Outlook Special Report; IEA: Paris, France, 2020. [Google Scholar]
- Brémond, U.; Bertrandias, A.; Steyer, J.P.; Bernet, N.; Carrere, H. A vision of European biogas sector development towards 2030: Trends and challenges. J. Clean. Prod. 2021, 287, 125065. [Google Scholar] [CrossRef]
- Monlau, F.; Sambusiti, C.; Ficara, E.; Aboulkas, A.; Barakat, A.; Carrère, H. New opportunities for agricultural digestate valorization: Current situation and perspectives. Energy Environ. Sci. 2015, 8, 2600–2621. [Google Scholar] [CrossRef]
- Brémond, U.; Bertrandias, A.; de Buyer, R.; Latrille, E.; Jimenez, J.; Escudié, R.; Steyer, J.-P.; Bernet, N.; Carrere, H. Recirculation of solid digestate to enhance energy efficiency of biogas plants: Strategies, conditions and impacts. Energy Convers. Manag. 2021, 231, 113759. [Google Scholar] [CrossRef]
- Brémond, U.; de Buyer, R.; Steyer, J.-P.; Bernet, N.; Carrere, H. Biological pretreatments of biomass for improving biogas production: An overview from lab scale to full-scale. Renew. Sustain. Energy Rev. 2018, 90, 583–604. [Google Scholar] [CrossRef]
- Ali, S.S.; Abomohra, A.E.F.; Sun, J. Effective bio-pretreatment of sawdust waste with a novel microbial consortium for enhanced biomethanation. Bioresour. Technol. 2017, 238, 425–432. [Google Scholar] [CrossRef]
- Zhang, Q.; He, J.; Tian, M.; Mao, Z.; Tang, L.; Zhang, J.; Zhang, H. Enhancement of methane production from cassava residues by biological pretreatment using a constructed microbial consortium. Bioresour. Technol. 2011, 102, 8899–8906. [Google Scholar] [CrossRef]
- Hua, B.; Dai, J.; Liu, B.; Zhang, H.; Yuan, X.; Wang, X.; Cui, Z. Pretreatment of non-sterile, rotted silage maize straw by the microbial community MC1 increases biogas production. Bioresour. Technol. 2016, 216, 699–705. [Google Scholar] [CrossRef]
- Zhong, C.; Wang, C.; Wang, F.; Jia, H.; Wei, P.; Zhao, Y. Enhanced biogas production from wheat straw with the application of synergistic microbial consortium pretreatment. RSC Adv. 2016, 6, 60187–60195. [Google Scholar] [CrossRef]
- Bertide, A. Recherche de relations entre le microbiote du rumen et la production laitière des brebis Lacaune. Ph.D. Thesis, Université Paul Sabatier (Toulouse 3), Ecole Nationale Vétérinaire de Toulouse, Toulouse, France, 2018. [Google Scholar]
- Plouchart, D.; Milferstedt, K.; Guizard, G.; Latrille, E.; Hamelin, J. Multiplexed chemostat system for quantification of biodiversity and ecosystem functioning in anaerobic digestion. PLoS ONE 2018, 13, e0193748. [Google Scholar] [CrossRef] [Green Version]
- Bandounas, L.; Wierckx, N.J.P.; de Winde, J.H.; Ruijssenaars, H.J. Isolation and characterization of novel bacterial strains exhibiting ligninolytic potential. BMC Biotechnol. 2011, 11, 94. [Google Scholar] [CrossRef] [Green Version]
- Jimenez, J.; Aemig, Q.; Doussiet, N.; Steyer, J.P.; Houot, S.; Patureau, D. A new organic matter fractionation methodology for organic wastes: Bioaccessibility and complexity characterization for treatment optimization. Bioresour. Technol. 2015, 194, 344–353. [Google Scholar] [CrossRef] [PubMed]
- Atlas, R.M. Handbook of Microbiological Media; CRC Press: Boca Raton, FL, USA, 2010; ISBN 978-1-4398-0406-3. [Google Scholar]
- Alberdi, A.; Gilbert, M.T.P. Hilldiv: An R package for the integral analysis of diversity based on Hill numbers. bioRxiv 2019, 545665. [Google Scholar] [CrossRef] [Green Version]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [Green Version]
- Whitman, W.B. Bergey’s Manual of Systematic Bacteriology—Volume Three the Firmicutes; Springer: New York, NY, USA, 2009; ISBN 978-0-387-95041-9. [Google Scholar]
- Thomas, F.; Hehemann, J.H.; Rebuffet, E.; Czjzek, M.; Michel, G. Environmental and gut Bacteroidetes: The food connection. Front. Microbiol. 2011, 2, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenberg, E. The Family Chitinophagaceae. In The Prokaryotes; Springer: New York, NY, USA, 2014; ISBN 978-3-642-38953-5. [Google Scholar]
- Mckee, L.S.; Martínez-Abad, A.; Ruthes, A.C.; Vilaplana, F.; Brumer, H. Focused Metabolism of Beta-Glucans by the Soil Bacteroidetes Species Chitinophaga pinensis. Appl. Environ. Microbiol. 2019, 85, e02231-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, N.; Castlebury, L.A.; Miller, A.N.; Huhndorf, S.M.; Schoch, C.L.; Seifert, K.A.; Rossman, A.Y.; Rogers, J.D.; Kohlmeyer, J.; Volkmann-Kohlmeyer, B.; et al. An overview of the systematics of the Sordariomycetes based on a four-gene phylogeny. Mycologia 2006, 98, 1076–1087. [Google Scholar] [CrossRef] [PubMed]
- Moletta, R.; Berge, S.; Bernet, N.; Bouchez, T.; Braumann, A.; Buffière, P.; Bultel, Y.; Camacho, P.; Carrère, H.; Chatain, V.; et al. La Méthanisation; Lavoisier: Cachan, France, 2008; ISBN 978-2-7430-1036-2. [Google Scholar]
- Brown, M.E.; Chang, M.C.Y. Exploring bacterial lignin degradation. Curr. Opin. Chem. Biol. 2014, 19, 1–7. [Google Scholar] [CrossRef]
- Kamimura, N.; Sakamoto, S.; Mitsuda, N.; Masai, E.; Kajita, S. Advances in microbial lignin degradation and its applications. Curr. Opin. Biotechnol. 2019, 56, 179–186. [Google Scholar] [CrossRef]
- Kamimura, N.; Takahashi, K.; Mori, K.; Araki, T.; Fujita, M.; Higuchi, Y.; Masai, E. Bacterial catabolism of lignin-derived aromatics: New findings in a recent decade: Update on bacterial lignin catabolism. Environ. Microbiol. Rep. 2017, 9, 679–705. [Google Scholar] [CrossRef] [Green Version]
- Leonowicz, A.; Cho, N.; Luterek, J.; Wilkolazka, A.; Wojtas-Wasilewska, M.; Matuszewska, A.; Hofrichter, M.; Wesenberg, D.; Rogalski, J. Fungal laccase: Properties and activity on lignin. J. Basic Microbiol. 2001, 41, 185–227. [Google Scholar] [CrossRef]
- Healy, J.B.; Young, L.Y. Anaerobic biodegradation of eleven aromatic compounds to methane. Appl. Environ. Microbiol. 1979, 38, 84–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eichlerová, I.; Homolka, L.; Žifčáková, L.; Lisá, L.; Dobiášová, P.; Baldrian, P. Enzymatic systems involved in decomposition reflects the ecology and taxonomy of saprotrophic fungi. Fungal Ecol. 2015, 13, 10–22. [Google Scholar] [CrossRef]
- Asina, F.N.U.; Brzonova, I.; Kozliak, E.; Kubátová, A.; Ji, Y. Microbial treatment of industrial lignin: Successes, problems and challenges. Renew. Sustain. Energy Rev. 2017, 77, 1179–1205. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, Q.; Yan, L.; Gao, Y.; Wang, Y.; Wang, W. A novel lignin degradation bacterial consortium for efficient pulping. Bioresour. Technol. 2013, 139, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Zhang, K.; Liu, P.; Han, H.; Zhao, S.; Kakade, A.; Khan, A.; Du, D.; Li, X. Lignin depolymerization and utilization by bacteria. Bioresour. Technol. 2018, 269, 557–566. [Google Scholar] [CrossRef]
- Fang, X.; Li, Q.; Lin, Y.; Lin, X.; Dai, Y.; Guo, Z.; Pan, D. Screening of a microbial consortium for selective degradation of lignin from tree trimmings. Bioresour. Technol. 2018, 254, 247–255. [Google Scholar] [CrossRef]
- Lazuka, A.; Auer, L.; Bozonnet, S.; Morgavi, D.P.; O’Donohue, M.; Hernandez-Raquet, G. Efficient anaerobic transformation of raw wheat straw by a robust cow rumen-derived microbial consortium. Bioresour. Technol. 2015, 196, 241–249. [Google Scholar] [CrossRef]
- Liang, J.; Fang, X.; Lin, Y.; Wang, D. A new screened microbial consortium OEM2 for lignocellulosic biomass deconstruction and chlorophenols detoxification. J. Hazard. Mater. 2018, 347, 341–348. [Google Scholar] [CrossRef]
- Maynaud, G.; Druilhe, C.; Daumoin, M.; Jimenez, J.; Patureau, D.; Torrijos, M.; Pourcher, A.M.; Wéry, N. Characterisation of the biodegradability of post-treated digestates via the chemical accessibility and complexity of organic matter. Bioresour. Technol. 2017, 231, 65–74. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Microorganisms | Acronyms | LAMACs Enrichment & Selection | Propagation Erlenmeyer Flasks | Aerobic Post-Treatment | BMP Tests |
|---|---|---|---|---|---|
| Natural environments | |||||
| Soil with wood decomposition | F1 | Straw | ×1 | ×3 | ×3 |
| Wood | ×1 | ×3 | ×3 | ||
| Deep forest litter | F2 | Straw | ×1 | ×3 | ×3 |
| Wood | ×1 | ×3 | ×3 | ||
| Rotten wood | F3 | Straw | ×1 | ×3 | ×3 |
| Wood | ×1 | ×3 | ×3 | ||
| Fresh sheep rumen | RU | Straw | ×1 | ×3 | ×3 |
| Wood | ×1 | ×3 | ×3 | ||
| Rotten wheat straw | RS | Straw | ×1 | ×3 | ×3 |
| Wood | ×1 | ×3 | ×3 | ||
| commercial granulated organic fertilizer | CF | Straw | ×1 | ×3 | ×3 |
| Wood | ×1 | ×3 | ×3 | ||
| Directly from SD via a washing step | Dig | ×2 | ×3 | ×3 | |
| Isolated from SD via Kraft lignin petri dishes | Petri | ×3 | ×3 | ×3 | |
| Directly from CF fertilizer via a washing step | Direct-CF | ×3 | ×3 | ×3 | |
| Sterilized complex SD | Sterility-check | ×3 | ×3 | ×3 | |
| Blanks | |||||
| Water added to SD instead of liquid consortia | Water | ×3 | ×3 | ||
| SD without post-treatment | Control | ×3 |
| Envfit Pr (>r) | Super Kingdom/Kingdom | Phylum | Class | Genus | Species | % ID | |
|---|---|---|---|---|---|---|---|
| B1 | 0.001 | Bacteria | Bacteroidetes | Chitinophagia (ex-Sphingobacteriia) | Pseudoflavitalea | Pseudoflavitalea sp. | 97.85 |
| B2 | 0.001 | Bacteria | Bacteroidetes | Chitinophagia (ex-Sphingobacteriia) | / | Uncultured Chitinophagaceae bacterium | 99.46 |
| B3 | 0.007 | Bacteria | Bacteroidetes | Chitinophagia (ex-Sphingobacteriia) | Terrimonas | Uncultured Terrimonas sp. | 98.39 |
| B4 | 0.003 | Bacteria | Bacteroidetes | / | / | Uncultured Bacteroidetes bacterium | 96.12 |
| B5 | 0.008 | Bacteria | Firmicutes | Bacilli | Paenibacillus | Paenibacillus sp. | 97.33 |
| B6 | 0.009 | Bacteria | Firmicutes | Bacilli | Paenibacillus | Enrichment culture clone LDC-5 | 99.47 |
| E1 | 0.001 | Eukaryota/Fungi | Ascomycota | Sordariomycetes | Scopulariopsis | Scopulariopsis sp. | 98.76 |
| E2 | 0.003 | Eukaryota/Fungi | Ascomycota | Sordariomycetes | Acremonium | Acremonium sp. | 98.55 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brémond, U.; Bertrandias, A.; Hamelin, J.; Milferstedt, K.; Bru-Adan, V.; Steyer, J.-P.; Bernet, N.; Carrere, H. Screening and Application of Ligninolytic Microbial Consortia to Enhance Aerobic Degradation of Solid Digestate. Microorganisms 2022, 10, 277. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10020277
Brémond U, Bertrandias A, Hamelin J, Milferstedt K, Bru-Adan V, Steyer J-P, Bernet N, Carrere H. Screening and Application of Ligninolytic Microbial Consortia to Enhance Aerobic Degradation of Solid Digestate. Microorganisms. 2022; 10(2):277. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10020277
Chicago/Turabian StyleBrémond, Ulysse, Aude Bertrandias, Jérôme Hamelin, Kim Milferstedt, Valérie Bru-Adan, Jean-Philippe Steyer, Nicolas Bernet, and Hélène Carrere. 2022. "Screening and Application of Ligninolytic Microbial Consortia to Enhance Aerobic Degradation of Solid Digestate" Microorganisms 10, no. 2: 277. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10020277