
Analysis of Ensifer aridi Mutants Affecting Regulation of Methionine, Trehalose, and Inositol Metabolisms Suggests a Role in Stress Adaptation and Symbiosis Development


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Plasmids
2.2. Mutants’ Constructions
2.3. Phenotypic Characterization of the Mutants
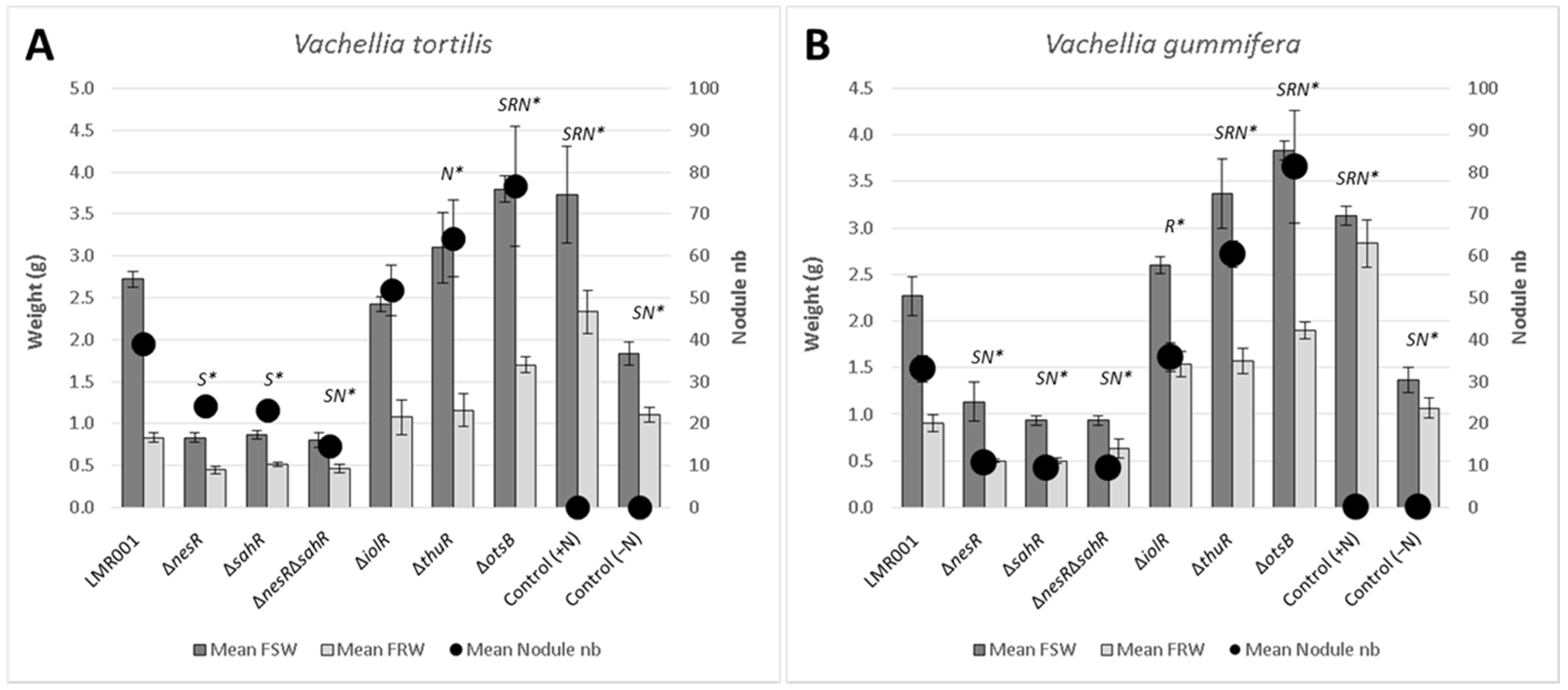
2.3.1. Plant Nodulation Assay
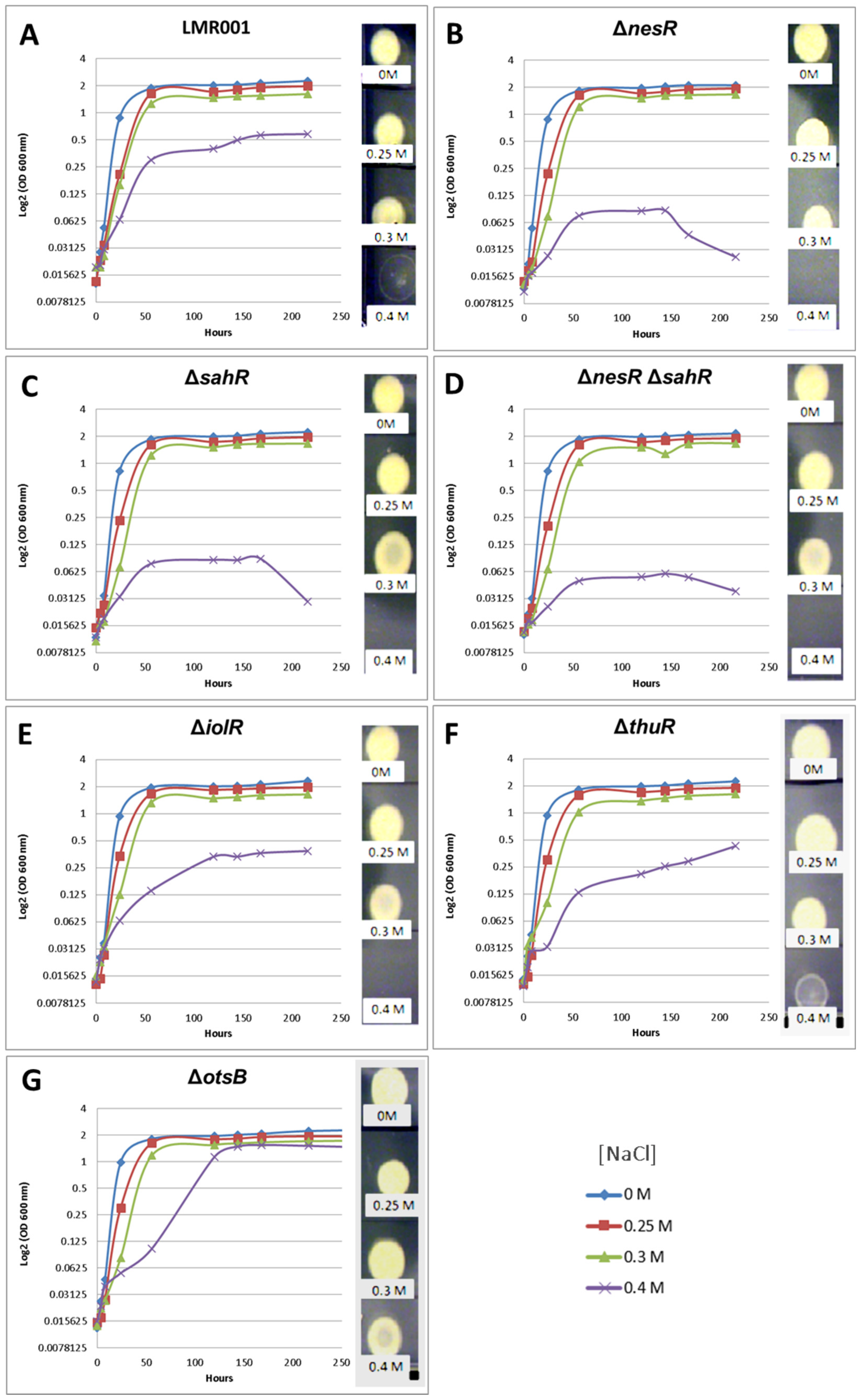
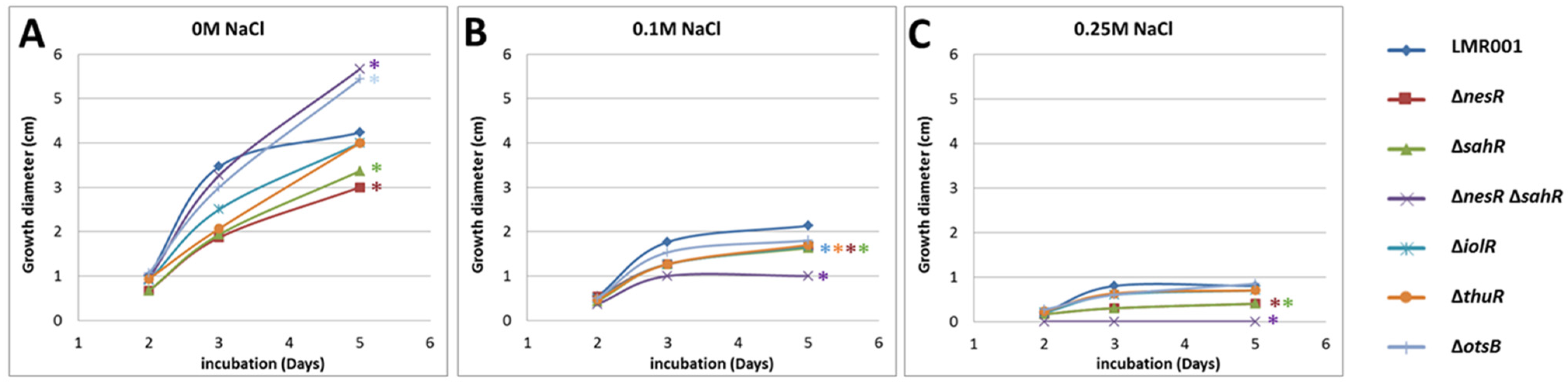
2.3.2. Motility, Tolerance to Salt, Detergent, Acid and Oxidative Stresses
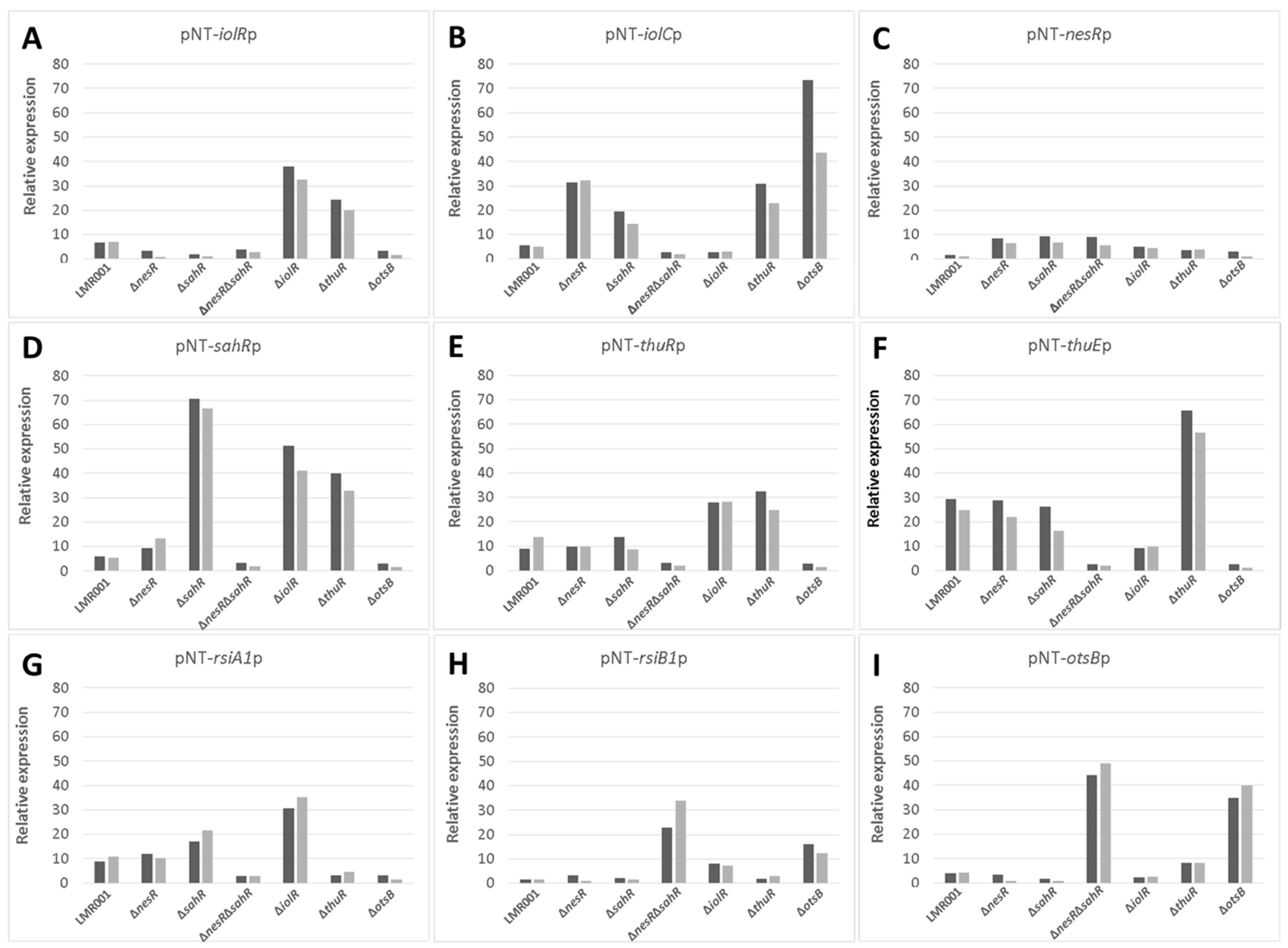
2.4. Transcirptional Fusion Assays
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hakim, S.; Imran, A.; Mirza, M.S. Phylogenetic Diversity Analysis Reveals Bradyrhizobium Yuanmingense and Ensifer aridi as Major Symbionts of Mung Bean (Vigna radiata L.) in Pakistan. Braz. J. Microbiol. 2021, 52, 311–324. [Google Scholar] [CrossRef] [PubMed]
- Lamin, H.; Alami, S.; Bouhnik, O.; ElFaik, S.; Abdelmoumen, H.; Bedmar, E.J.; Missbah-El Idrissi, M. Nodulation of Retama monosperma by Ensifer aridi in an Abandonned Lead Mine Soils in Eastern Morocco. Front. Microbiol. 2019, 10, 1456. [Google Scholar] [CrossRef] [PubMed]
- Rocha, G.; Le Queré, A.; Medina, A.; Cuéllar, A.; Contreras, J.-L.; Carreño, R.; Bustillos, R.; Muñoz-Rojas, J.; del Carmen Villegas, M.; Chaintreuil, C.; et al. Diversity and Phenotypic Analyses of Salt- and Heat-Tolerant Wild Bean Phaseolus filiformis Rhizobia Native of a Sand Beach in Baja California and Description of Ensifer aridi sp. Nov. Arch. Microbiol. 2020, 202, 309–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakrouhi, I.; Belfquih, M.; Sbabou, L.; Moulin, P.; Bena, G.; Filali-Maltouf, A.; Le Quéré, A. Recovery of Symbiotic Nitrogen Fixing Acacia Rhizobia from Merzouga Desert Sand Dunes in South East Morocco—Identification of a Probable New Species of Ensifer Adapted to Stressed Environments. Syst. Appl. Microbiol. 2016, 39, 122–131. [Google Scholar] [CrossRef]
- Tak, N.; Awasthi, E.; Bissa, G.; Meghwal, R.R.; James, E.K.; Sprent, J.S.; Gehlot, H.S. Multi Locus Sequence Analysis and Symbiotic Characterization of Novel Ensifer Strains Nodulating Tephrosia Spp. in the Indian Thar Desert. Syst. Appl. Microbiol. 2016, 39, 534–545. [Google Scholar] [CrossRef]
- Le Quéré, A.; Tak, N.; Gehlot, H.S.; Lavire, C.; Meyer, T.; Chapulliot, D.; Rathi, S.; Sakrouhi, I.; Rocha, G.; Rohmer, M.; et al. Genomic Characterization of Ensifer aridi, a Proposed New Species of Nitrogen-Fixing Rhizobium Recovered from Asian, African and American Deserts. BMC Genom. 2017, 18, 85. [Google Scholar] [CrossRef]
- Cytryn, E.J.; Sangurdekar, D.P.; Streeter, J.G.; Franck, W.L.; Chang, W.; Stacey, G.; Emerich, D.W.; Joshi, T.; Xu, D.; Sadowsky, M.J. Transcriptional and Physiological Responses of Bradyrhizobium Japonicum to Desiccation-Induced Stress. J. Bacteriol. 2007, 189, 6751–6762. [Google Scholar] [CrossRef] [Green Version]
- López-Leal, G.; Tabche, M.; Castillo-Ramírez, S.; Mendoza-Vargas, A.; Ramírez-Romero, M.A.; Dávila, G. RNA-Seq Analysis of the Multipartite Genome of Rhizobium etli CE3 Shows Different Replicon Contributions under Heat and Saline Shock. BMC Genom. 2014, 15, 770. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Montaño, F.; del Cerro, P.; Jiménez-Guerrero, I.; López-Baena, F.J.; Cubo, M.T.; Hungria, M.; Megías, M.; Ollero, F.J. RNA-Seq Analysis of the Rhizobium tropici CIAT 899 Transcriptome Shows Similarities in the Activation Patterns of Symbiotic Genes in the Presence of Apigenin and Salt. BMC Genom. 2016, 17, 198. [Google Scholar] [CrossRef]
- Alexandre, A.; Laranjo, M.; Oliveira, S. Global Transcriptional Response to Heat Shock of the Legume Symbiont Mesorhizobium loti MAFF303099 Comprises Extensive Gene Downregulation. DNA Res. 2014, 21, 195–206. [Google Scholar] [CrossRef] [Green Version]
- Laranjo, M.; Alexandre, A.; Oliveira, S. Genes Commonly Involved in Acid Tolerance Are Not Overexpressed in the Plant Microsymbiont Mesorhizobium loti MAFF303099 upon Acidic Shock. Appl. Microbiol. Biotechnol. 2014, 98, 7137–7147. [Google Scholar] [CrossRef] [PubMed]
- Laranjo, M.; Alexandre, A.; Oliveira, S. Global Transcriptional Response to Salt Shock of the Plant Microsymbiont Mesorhizobium loti MAFF303099. Res. Microbiol. 2017, 168, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Luo, Y.; Mohamed, O.A.; Liu, D.; Wei, G. Global Transcriptome Analysis of Mesorhizobium alhagi CCNWXJ12-2 under Salt Stress. BMC Microbiol. 2014, 14, 319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maynaud, G.; Brunel, B.; Mornico, D.; Durot, M.; Severac, D.; Dubois, E.; Navarro, E.; Cleyet-Marel, J.-C.; Le Quéré, A. Genome-Wide Transcriptional Responses of Two Metal-Tolerant Symbiotic Mesorhizobium Isolates to Zinc and Cadmium Exposure. BMC Genom. 2013, 14, 292. [Google Scholar] [CrossRef] [Green Version]
- Belfquih, M.; Sakrouhi, I.; Ait-Benhassou, H.; Dubois, E.; Severac, D.; Filali-Maltouf, A.; Le Quere, A. Ensifer aridi LMR001T Symbiosis and Tolerance to Stress Do Not Require the Alternative Sigma Factor RpoE2. Agronomy 2021, 11, 1787. [Google Scholar] [CrossRef]
- de Lucena, D.K.; Pühler, A.; Weidner, S. The Role of Sigma Factor RpoH1 in the PH Stress Response of Sinorhizobium meliloti. BMC Microbiol. 2010, 10, 265. [Google Scholar] [CrossRef] [Green Version]
- Domínguez-Ferreras, A.; Pérez-Arnedo, R.; Becker, A.; Olivares, J.; Soto, M.J.; Sanjuán, J. Transcriptome Profiling Reveals the Importance of Plasmid PSymB for Osmoadaptation of Sinorhizobium meliloti. J. Bacteriol. 2006, 188, 7617–7625. [Google Scholar] [CrossRef] [Green Version]
- Hellweg, C.; Pühler, A.; Weidner, S. The Time Course of the Transcriptomic Response of Sinorhizobium meliloti 1021 Following a Shift to Acidic PH. BMC Microbiol. 2009, 9, 37. [Google Scholar] [CrossRef] [Green Version]
- Lu, M.; Jiao, S.; Gao, E.; Song, X.; Li, Z.; Hao, X.; Rensing, C.; Wei, G. Transcriptome Response to Heavy Metals in Sinorhizobium meliloti CCNWSX0020 Reveals New Metal Resistance Determinants That Also Promote Bioremediation by Medicago lupulina in Metal-Contaminated Soil. Appl. Environ. Microbiol. 2017, 83, e01244-17. [Google Scholar] [CrossRef] [Green Version]
- Sauviac, L.; Philippe, H.; Phok, K.; Bruand, C. An Extracytoplasmic Function Sigma Factor Acts as a General Stress Response Regulator in Sinorhizobium meliloti. J. Bacteriol. 2007, 189, 4204–4216. [Google Scholar] [CrossRef] [Green Version]
- Vriezen, J.A.C.; de Bruijn, F.J.; Nüsslein, K. Responses of Rhizobia to Desiccation in Relation to Osmotic Stress, Oxygen, and Temperature. Appl. Environ. Microbiol. 2007, 73, 3451–3459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beringer, J.E. R Factor Transfer in Rhizobium leguminosarum. Microbiology 1974, 84, 188–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sambrook, J.; Fritsch, E.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor: Long Island, NY, USA, 1989; Volume 2. [Google Scholar]
- Hanahan, D. Studies on Transformation of Escherichia coli with Plasmids. J. Mol. Biol. 1983, 166, 557–580. [Google Scholar] [CrossRef]
- Simon, R.; Priefer, U.; Pühler, A. A Broad Host Range Mobilization System for In Vivo Genetic Engineering: Transposon Mutagenesis in Gram Negative Bacteria. Bio/Technology 1983, 1, 784–791. [Google Scholar] [CrossRef]
- Quandt, J.; Hynes, M.F. Versatile Suicide Vectors Which Allow Direct Selection for Gene Replacement in Gram-Negative Bacteria. Gene 1993, 127, 15–21. [Google Scholar] [CrossRef]
- Figurski, D.H.; Helinski, D.R. Replication of an Origin-Containing Derivative of Plasmid RK2 Dependent on a Plasmid Function Provided in Trans. Proc. Natl. Acad. Sci. USA 1979, 76, 1648–1652. [Google Scholar] [CrossRef] [Green Version]
- Miller, W.G.; Leveau, J.H.J.; Lindow, S.E. Improved Gfp and InaZ Broad-Host-Range Promoter-Probe Vectors. Mol. Plant-Microbe Interact. 2000, 13, 1243–1250. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Kuo, T. A Simple and Rapid Method for the Preparation of Gram-Negative Bacterial Genomic DNA. Nucleic Acids Res. 1993, 21, 2260. [Google Scholar] [CrossRef]
- Broughton, W.J.; Dilworth, M.J. Control of Leghaemoglobin Synthesis in Snake Beans. Biochem. J. 1971, 125, 1075–1080. [Google Scholar] [CrossRef] [Green Version]
- Patankar, A.V.; González, J.E. An Orphan LuxR Homolog of Sinorhizobium meliloti Affects Stress Adaptation and Competition for Nodulation. Appl. Environ. Microbiol. 2009, 75, 946–955. [Google Scholar] [CrossRef] [Green Version]
- Novichkov, P.S.; Li, X.; Kuehl, J.V.; Deutschbauer, A.M.; Arkin, A.P.; Price, M.N.; Rodionov, D.A. Control of Methionine Metabolism by the SahR Transcriptional Regulator in Proteobacteria: Control of Methionine Metabolism in Proteobacteria. Environ. Microbiol. 2014, 16, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robledo, M.; García-Tomsig, N.I.; Matia-González, A.M.; García-Rodríguez, F.M.; Jiménez-Zurdo, J.I. Synthetase of the Methyl Donor S-Adenosylmethionine from Nitrogen-Fixing α-Rhizobia Can Bind Functionally Diverse RNA Species. RNA Biol. 2021, 18, 1111–1123. [Google Scholar] [CrossRef] [PubMed]
- Ravcheev, D.A.; Khoroshkin, M.S.; Laikova, O.N.; Tsoy, O.V.; Sernova, N.V.; Petrova, S.A.; Rakhmaninova, A.B.; Novichkov, P.S.; Gelfand, M.S.; Rodionov, D.A. Comparative Genomics and Evolution of Regulons of the LacI-Family Transcription Factors. Front. Microbiol. 2014, 5, 294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suárez, R.; Wong, A.; Ramírez, M.; Barraza, A.; Orozco, M.D.C.; Cevallos, M.A.; Lara, M.; Hernández, G.; Iturriaga, G. Improvement of Drought Tolerance and Grain Yield in Common Bean by Overexpressing Trehalose-6-Phosphate Synthase in Rhizobia. Mol. Plant-Microbe Interact. 2008, 21, 958–966. [Google Scholar] [CrossRef] [Green Version]
- Moussaid, S.; Domínguez-Ferreras, A.; Muñoz, S.; Aurag, J.; Berraho, E.B.; Sanjuán, J. Increased Trehalose Biosynthesis Improves Mesorhizobium ciceri Growth and Symbiosis Establishment in Saline Conditions. Symbiosis 2015, 67, 103–111. [Google Scholar] [CrossRef]
- Paul, M.J.; Watson, A.; Griffiths, C.A. Trehalose 6-Phosphate Signalling and Impact on Crop Yield. Biochem. Soc. Trans. 2020, 48, 2127–2137. [Google Scholar] [CrossRef]
- Baena-González, E.; Lunn, J.E. SnRK1 and Trehalose 6-Phosphate—Two Ancient Pathways Converge to Regulate Plant Metabolism and Growth. Curr. Opin. Plant Biol. 2020, 55, 52–59. [Google Scholar] [CrossRef]
- da-Silva, J.R.; Alexandre, A.; Brígido, C.; Oliveira, S. Can stress response genes be used to improve the symbiotic performance of rhizobia? AIMS Microbiol. 2017, 3, 365–382. [Google Scholar] [CrossRef]
- Kohler, P.R.A.; Zheng, J.Y.; Schoffers, E.; Rossbach, S. Inositol Catabolism, a Key Pathway in Sinorhizobium meliloti for Competitive Host Nodulation. Appl. Environ. Microbiol. 2010, 76, 7972–7980. [Google Scholar] [CrossRef] [Green Version]
- Kohler, P.R.A.; Choong, E.-L.; Rossbach, S. The RpiR-Like Repressor IolR Regulates Inositol Catabolism in Sinorhizobium meliloti. J. Bacteriol. 2011, 193, 5155–5163. [Google Scholar] [CrossRef] [Green Version]
- Dong, Y.; Li, S.; Zhao, D.; Liu, J.; Ma, S.; Geng, J.; Lu, C.; Liu, Y. IolR, a Negative Regulator of the Myo-Inositol Metabolic Pathway, Inhibits Cell Autoaggregation and Biofilm Formation by Downregulating RpmA in Aeromonas hydrophila. NPJ Biofilms Microbiomes 2020, 6, 22. [Google Scholar] [CrossRef] [PubMed]
- Lang, C.; Barnett, M.J.; Fisher, R.F.; Smith, L.S.; Diodati, M.E.; Long, S.R. Most Sinorhizobium meliloti Extracytoplasmic Function Sigma Factors Control Accessory Functions. mSphere 2018, 3, e00454-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, J.B.; Peters, N.K.; Bhuvaneswari, T.V. Redundancy in Periplasmic Binding Protein-Dependent Transport Systems for Trehalose, Sucrose, and Maltose in Sinorhizobium meliloti. J. Bacteriol. 2002, 184, 2978–2986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- diCenzo, G.C.; Muhammed, Z.; Østerås, M.; O’Brien, S.A.P.; Finan, T.M. A Key Regulator of the Glycolytic and Gluconeogenic Central Metabolic Pathways in Sinorhizobium meliloti. Genetics 2017, 207, 961–974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onwe, R.O.; Onwosi, C.O.; Ezugworie, F.N.; Ekwealor, C.C.; Okonkwo, C.C. Microbial Trehalose Boosts the Ecological Fitness of Biocontrol Agents, the Viability of Probiotics during Long-Term Storage and Plants Tolerance to Environmental-Driven Abiotic Stress. Sci. Total Environ. 2022, 806, 150432. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, S.; Banerjee, S.; Lahiri, S.; Dutta, T.; Dhar, T.K.; Ghosh, A.K. Purification, Characterization, Sequencing and Molecular Cloning of a Novel Cysteine Methyltransferase That Regulates Trehalose-6-Phosphate Synthase from Saccharomyces cerevisiae. Biochim. Biophys. Acta 2014, 1840, 1861–1871. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain/Plasmid Name | Description | Source |
|---|---|---|
| Bacterial strains | ||
| LMR001T | Ensifer aridi LMR001T isolated from Vachellia gummifera grown in Merzouga desert sand dune | [4] |
| LMR001∆nesR | nesR deletion mutant of LMR001 | This work |
| LMR001∆sahR | sahR deletion mutant of LMR001 | This work |
| LMR001∆nesR∆sahR | nesR deletion mutant of LMR001∆sahR | This work |
| LMR001∆iolR | iolR deletion mutant of LMR001 | This work |
| LMR001∆thuR | thuR deletion mutant of LMR001 | This work |
| LMR001∆otsB | otsB deletion mutant of LMR001 | This work |
| E. coli DH5α | hsdR17 endA1 thi-1 gyrA96 relA1 recA1 supE44 DlacU169 (f80lacZDM15) | [24] |
| E. coli S17-1 | recA [SmR], thi, pro, RP4-2-Tc:Mu: aph::Tn7λpir. | [25] |
| E. coli XL2 Blue Ultra-competent | endA1 supE44 thi-1 hsdR17 recA1 gyrA96 relA1 lac [F’ proAB lacIqZΔM15 Tn10 (TetR) Amy CamR] | Stratagene |
| Plasmids | ||
| pJQ200SK | pACYC184-derived (p15A) suicide vector (GmR) | [26] |
| pRK2013 | Helper plasmid containing the ColE1 replicon with RK2 tra genes (KmR) | [27] |
| pGEM-T Easy | pUC origin, Multi Cloning Sites (MCS), lacZ gene fusion, β-lactamase coding region, lac operon sequences (ApR) | Promega |
| pBluescript KS(+) | ColEI, F1 Origin, MCS, lacZ gene fusion, β-lactamase coding region, lac operon sequences (ApR) | Stratagene |
| TOPO vecteur | pUC origin, région Plac promoteur, lacZ α-ccdB gene fuision. Topoisomerase enzyme (KmR, ApR) | Invitrogen |
| pPROBE-NT | pVS1-derived (p15a) vector, gfp (GmR) | [28] |
| pNT-sahRp | pPROBE NT containing sahR (PEG2239) promoter fused to gfp (KmR) | [15] |
| pNT-nesRp | pPROBE NT containing nesR (PEG5735) promoter fused to gfp (KmR) | [15] |
| pNT-rsiA1p | pPROBE NT containing rsiA1 (PEG2540) promoter fused to gfp (KmR) | [15] |
| pNT-rsiB1p | pPROBE NT containing rsiB1 (PEG2541) promoter fused to gfp (KmR) | [15] |
| pNT-thuEp | pPROBE NT containing thuE (PEG6268) promoter fused to gfp (KmR) | [15] |
| pNT-otsBp | pPROBE NT containing otsB (PEG4868) promoter fused to gfp (KmR) | [15] |
| pNT-thuRp | pPROBE NT containing thuR (PEG6269) promoter fused to gfp (KmR) | [15] |
| pNT-iolCp | pPROBE NT containing iolC (PEG373) promoter fused to gfp (KmR) | [15] |
| pNT-iolRp | pPROBE NT containing iolR (PEG374) promoter fused to gfp (KmR) | [15] |
| Target Gene (bp) | Primer * | Sequence (5′-3′) * | 5′ Region Size (bp)/Distance to Start (nt) | 3′ Region Size (bp)/Distance to Stop (nt) | Deletion (bp) |
|---|---|---|---|---|---|
| iolR (861) | IOLR-P374-A-XI | CCTCTAGACGAGATTTACGGCTCCAAGG | 457/−10 | 689 | |
| IOLR-P374-B-HIII | TCAAAGACACCACAAGCTTCTCCAGCTTGCTCGTGTTC | ||||
| IOLR-P374-C-HIII | AAGCTTGTGGTGTCTTTGACCGACTC | 658/−183 | |||
| IOLR-P374-D-XI | CCTCTAGAGCAATTGTCGCGATAGAAGA | ||||
| otsB (750) | OSTB-P4868-A-XI | GGTCTAGATGCGCGATCTTCATGAACAA | 454/+75 | 503 | |
| OSTB-P4868-B-HIII | TTCATCCGTCAGAAAGCTTGTCGGAGCAATGTCGAGAAG | ||||
| OSTB-P4868-C-HIII | AAGCTTTCTGACGGATGAAGGAATGTT | 500/−172 | |||
| OSTB-P4868-D-XI | GGTCTAGAATAGCCGGACATCTCATGCC | ||||
| thuR (1020) | THUR-P6269-A-XI | GGTCTAGAGGTCCTCAGAAGCATTGTCA | 473/+87 | 865 | |
| THUR-P6269-B-HIII | GTTCGTGAATCTCAAGCTTCTCGCTGACCTCGGGATAG | ||||
| THUR-P6269-C-HIII | AAGCTTGAGATTCACGAACTCTGGCC | 453/−69 | |||
| THUR-P6269-D-XI | GGTCTAGATAGGTGCGATGAACATGACG | ||||
| nesR (783) | NESR-P5735-A-XI | CGCTCTAGAGGCTCGATATCACGCCAC | 186/+39 | 749 | |
| NESR-P5735-B | TTAAGCCGCGGCGGTGAACTGGTTCCTGATC | ||||
| NESR-P5735-C | GCCGCGGCTTAAAACACC | 160/+6 | |||
| NESR-P5735-D-XI | GCGTCTAGAACGACAAGCGGCAGCTTGC | ||||
| sahR (1023) | SAHR-P2239-A-AI | GGGCCCGACAACATCTCGAAGG | 1062/+36 | 973 | |
| SAHR-P2239-B-PI | CTGCAGAGCATCCAAACCAAGCG | ||||
| SAHR-P2239-C-PI | CTGCAGGGGGAGTTTGAGAATGA | 1132/−14 | |||
| SAHR-P2239-D-XI | TCTAGACCGAATTAGGGACTATAATTCCG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Belfquih, M.; Filali-Maltouf, A.; Le Quéré, A. Analysis of Ensifer aridi Mutants Affecting Regulation of Methionine, Trehalose, and Inositol Metabolisms Suggests a Role in Stress Adaptation and Symbiosis Development. Microorganisms 2022, 10, 298. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10020298
Belfquih M, Filali-Maltouf A, Le Quéré A. Analysis of Ensifer aridi Mutants Affecting Regulation of Methionine, Trehalose, and Inositol Metabolisms Suggests a Role in Stress Adaptation and Symbiosis Development. Microorganisms. 2022; 10(2):298. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10020298
Chicago/Turabian StyleBelfquih, Meryem, Abdelkarim Filali-Maltouf, and Antoine Le Quéré. 2022. "Analysis of Ensifer aridi Mutants Affecting Regulation of Methionine, Trehalose, and Inositol Metabolisms Suggests a Role in Stress Adaptation and Symbiosis Development" Microorganisms 10, no. 2: 298. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10020298