
Beyond the ABCs—Discovery of Three New Plasmid Types in Rhodobacterales (RepQ, RepY, RepW)


1
Leibniz-Institut DSMZ—Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH, Inhoffenstraße 7 B, 38124 Braunschweig, Germany
2
Institut für Mikrobiologie, Technische Universität Braunschweig, Spielmannstraße 7, 38106 Braunschweig, Germany
*
Authors to whom correspondence should be addressed.
Microorganisms 2022, 10(4), 738; https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10040738
Submission received: 1 March 2022
/
Revised: 18 March 2022
/
Accepted: 24 March 2022
/
Published: 29 March 2022
(This article belongs to the Special Issue Mobile Genetic Elements in Adaptation of Bacteria to the Changing Environment)
Abstract
:Copiotrophic marine bacteria of the Roseobacter group (Rhodobacterales, Alphaproteobacteria) are characterized by a multipartite genome organization. We sequenced the genomes of Sulfitobacter indolifex DSM 14862T and four related plasmid-rich isolates in order to investigate the composition, distribution, and evolution of their extrachromosomal replicons (ECRs). A combination of long-read PacBio and short-read Illumina sequencing was required to establish complete closed genomes that comprised up to twelve ECRs. The ECRs were differentiated in stably evolving chromids and genuine plasmids. Among the chromids, a diagnostic RepABC-8 replicon was detected in four Sulfitobacter species that likely reflects an evolutionary innovation that originated in their common ancestor. Classification of the ECRs showed that the most abundant plasmid system is RepABC, followed by RepA, DnaA-like, and RepB. However, the strains also contained three novel plasmid types that were designated RepQ, RepY, and RepW. We confirmed the functionality of their replicases, investigated the genetic inventory of the mostly cryptic plasmids, and retraced their evolutionary origin. Remarkably, the RepY plasmid of S. pontiacus DSM 110277 is the first high copy-number plasmid discovered in Rhodobacterales.
1. Introduction
Bacteria are capable of rapidly adapting to changing environments via the acquisition of mobile genetic elements [1,2,3]. Proteobacterial plasmids carry a wide range of accessory genes that are beneficial for their bacterial hosts and might even pave the way for the colonization of novel environmental niches. In addition to the spread of antibiotic resistances [4,5,6], they provide protection against heavy metal and xenobiotic pollutants [3,7,8], facilitate interactions with algae and plants [9,10], mediate the formation of biofilms [11], and allow the utilization of specific carbon sources [12,13]. Even large genetic units with sizes of more than 40 kb, such as the gene clusters for aerobic anoxygenic photosynthesis or the formation of functional flagella, are occasionally encoded on extrachromosomal replicons (ECRs) [14,15].
Regardless of the highly variable accessory genes, plasmids are characterized by their essential replicase, and a set of conserved backbone genes required for their maintenance and transfer [3,16]. Low copy number plasmids comprise a partitioning system for stable maintenance homologous to those of the bacterial chromosome. It consists of the centromere-like binding site, a DNA-binding protein (ParB), and a motor protein (ParA) that mediates a concerted transfer of the replicated plasmids to the cell poles, thus ensuring their reliable distribution to the daughter cells [17]. In contrast, high copy number plasmids lack a partitioning system and are randomly distributed during bacterial cell division. Other characteristic backbone genes are toxin-antitoxin systems representing addiction modules that prevent the spontaneous loss of a plasmid [18]. The horizontal transfer of plasmids is essentially mediated by conserved type IV secretion systems (T4SSs) that encode the crucial relaxase for the release of single-stranded DNA, a coupling protein, and a sophisticated nanotube for their conjugative transmission [19]. Many cryptic plasmids, which are small selfish replicons without beneficial genes for the host, contain only mobilization (MOB) genes encoding the pivotal relaxase and therefore require conjugative plasmids with T4SSs for their interbacterial exchange [20]. The majority of plasmids lack the known MOB genes and are therefore considered non-mobilizable, although comparative analyses suggested that they were once horizontally transferred [20,21].
The only indispensable unit of a plasmid is its replication system, which includes the origin of replication (oriV) and a diagnostic replicase. Replication proteins are reliable markers for the plasmid classification, and phylogenetic analyses of homologous replicases even allow the discrimination of different compatibility groups [22,23]. A trustworthy and reproducible classification of ECRs is crucial for the comparison of plasmids from different bacterial lineages and the identification of novel plasmid types. Based on the phylogenetic approach, six plasmid types were so far identified within the alphaproteobacterial order Rhodobacterales. These are RepA, RepB, RepABC, DnaA-like, RepL, and RepC_soli plasmids that encompass up to nine different compatibility groups [4,7,24]. However, an analysis of small plasmids from four Paracoccus strains indicated the presence of further replicon types [25].
Many of the mentioned plasmid types were first detected in roseobacters (Roseobacteraceae), a highly metabolically and ecologically versatile group of marine Rhodobacterales [26,27]. Copiotrophic roseobacters typically exhibit a multipartite genome organization and are thus particularly suitable models to investigate the diversity and biology of plasmids [7,15]. However, a prerequisite for a systematic assessment of bacterial plasmids and the detection of novel replicon types is the availability of complete closed genomes in which all extrachromosomal elements were sequenced, e.g., [28]. Although more than 3000 Rhodobacterales genomes are available at the NCBI, over 90% of them are draft versions, which prevent the detection and unambiguous assignment of uncharacterized replicases to ECRs and thus the identification of novel plasmid types. In the current study, we established closed genomes of five different roseobacter strains from the genera Sulfitobacter and Pseudosulfitobacter, which was recently separated from Sulfitobacter as a novel genus [29]. These genera contain metabolically versatile and biogeographically widespread bacterial generalists [30,31,32]. Comparative analyses revealed the presence of three novel plasmid types designated RepQ, RepY, and RepW.
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
Five bacterial Sulfitobacter strains (DSM 14862, 2RS2_G6, 3RS2_G4b, SO248Ex84, PIC-76) were investigated in this study. Strain DSM 14862T was obtained from the Leibniz Institute DSMZ. Strains 2RS2_G6 and 3RS2_G4b were isolated from sediment samples collected in the Channel Sea near Roscoff (48.7205, −3.9651) on the 26 September 2013. Strain SO248Ex84 was isolated by Sara Billerbeck (Institute for Chemistry and Biology of the Marine Environment [ICBM], Oldenburg, Germany) from Pacific Ocean seawater collected during the Sonne cruise SO248 (45.0001, 178.7499) on the 24 May 2016. Strain PIC-76 was provided by Irene Wagner-Döbler [33]. All strains are now deposited at the Leibniz Institute DSMZ under the following numbers: DSM 14862T, DSM 110093, DSM 109990, DSM 110277, and DSM 107133.
Functionality tests of putative replication genes were conducted with Phaeobacter inhibens DSM 17395 Δ65/3 lacking the biofilm chromid for surface attachment [4]. P. inhibens and the (Pseudo)sulfitobacter strains were cultivated in marine broth medium (MB, Carl Roth, or Difco) at 28 °C and 20 °C, respectively. Competent NEB®Turbo Escherichia coli cells (New England Biolab), used for cloning, were cultivated in Luria-Bertani medium (LB, Carl Roth) at 37 °C. The selection of transformants was performed on 0.5× MB or LB plates with 120 µg mL−1 kanamycin (KAN).
2.2. Genome Sequencing, Assembly and Annotation
Strains were harvested in the early stationary phase. Genomic DNA was extracted with the QIAGEN Genomic-tips 100 Kit (Qiagen, Hilden, Germany). SMRTbell™ template libraries were prepared according to the recommended instructions, 5 µg of genomic DNA were end-repaired and ligated to hairpin adapters using P6 chemistry (Pacific Biosciences, Menlo Park, CA, USA). BluePippin™ Size-Selection to greater than 3 kb was performed according to the manufacturer’s instructions (Sage Science, Beverly, MA, USA). SMRT sequencing was carried out on the PacBio RSII platform (Pacific Biosciences). To evaluate the recovery of small ECRs, deep SMRT sequencing of libraries that were constructed without size selection was performed on the Sequel II system (Pacific Biosciences), taking a 15 or 30 h movie. Illumina libraries were prepared with the Nextera XT DNA Sample Preparation Kit (lllumina Inc., San Diego, CA, USA) with a modified protocol [34], and paired-end Illumina sequencing was either performed on the NextSeq 500 (PE75 or PE150) or for strain PIC-76 on the MiSeq platform (PE300).
PacBio reads were assembled de novo with HGAP3 in SMRT Portal 2.3.0 or for the exemplary Sequel reads using the “Microbial Assembly” protocol in SMRT Link 10.0.0. The assembled contigs were error-corrected by mapping of Illumina short reads using the Burrows-Wheeler Aligner (BWA 0.6.2) [35] and subsequent variant and consensus calling using VarScan 2.3.6 [36]. An additional Illumina short read assembly was performed using velvet 1.2.10 [37], which was also corrected as mentioned before. Redundancies in parallel assemblies were identified with the Genome Finishing Tool of the CLC Genomics Workbench 7.0.1 and removed. The resulting assembly was trimmed, circularized, and adjusted to the replication system as a start point (https://github.com/boykebunk/genomefinish, accessed on 1 February 2022), checked via mapping of Illumina (BWA) and PacBio reads (RSII: Bridgemapper; Sequel: Resequencing) and finally corrected if necessary. Methylation motifs were detected with the RS Modification and Motif Analysis in SMRT Portal. The genomes were annotated with Prokka 1.13 [38] with a subsequent manual curation of the replication systems. The complete genomes are deposited in NCBI GenBank under the accession numbers: CP084951-58, CP084959-65, CP085144-53, CP085154-66, and CP085167-72.
2.3. Characterization and Comparison of Genomes, Phylogenetic Analyses and Data Handling
Illumina reads were mapped on the corresponding final assembly with BWA 0.6.2 [35] to estimate the copy number of the ECRs. The median coverage per element was calculated from the coverage per position extracted with SAMtools (v0.1.19) [39]. The copy number of ECRs was calculated as the ratio of their median coverage to the median chromosomal coverage. In order to differentiate genuine plasmids from chromids [40], four criteria were applied: (i) All chromids contained a parAB partitioning operon, (ii) represented low copy number replicons with coverage below two, (iii) had a GC content that deviated less than 2.5% from the chromosome, and (iv) showed a genomic imprint that was comparable to those of the chromosome. The last criterion was independently investigated with clustering analyses of the relative synonymous codon usage (CU) and the tetranucleotide frequency (tetra), both performed in R version 4.1. The well-characterized genome of Dinoroseobacter shibae DFL12 (GCA_000018145.1) served as an internal reference [41]. However, ECRs were only classified as chromids if the results of both methods (CU, tetra) agreed.
All replicons were screened for the presence of a type IV secretion system (T4SS) or a characteristic relaxase (mob with MOBscan https://castillo.dicom.unican.es/mobscan/, accessed on 29 September 2021), which is required for plasmid mobilization [20,42].
Taxonomic classification of the five investigated strains were based on genome sequences of all type strains from Sulfitobacter, Roseobacter, and Pseudosulfitobacter (including non-validated ones) mentioned at LPSN [43]. Further genomic comparisons were made with all closed Sulfitobacter genomes available at NCBI in October 2021, as well as closed genomes of 24 reference strains representing different clades within the Rhodobacterales (see Table S1). A genome-based phylogenetic tree was reconstructed from a concatenated amino acid alignment of 92 housekeeping genes generated with UBCG [44]. An approximately Maximum Likelihood phylogenetic tree was inferred using FastTree version 2.1.11 (double-precision) with default parameters [45]. The digital DNA-DNA hybridizations of selected strains were calculated with TYGS [46]. Replication systems of ECRs were identified and classified [4,7,24,47].
Novel replicases were compared with the Pfam database [48]. RepQ, RepY, and RepW type plasmid replication proteins have further been identified with BLASTP searches in the NCBI database. Most replicases were located on contigs from draft genomes, which makes it difficult to draw clear conclusions about their extrachromosomal localization. Accordingly, the 5′ and 3′ end of all linear contigs were compared via BLASTN, and a replicon was defined as circular if at least 50 nucleotides overlap with 100% identity.
The protein sequences of new replicases (RepQ, RepY, RepW) were aligned with muscle [49], and all positions containing gaps and missing data were eliminated. The evolutionary history was inferred by the Maximum Likelihood method based on the JTT matrix-based model [50]. The analyses, with 100 bootstrap replicates, were conducted in MEGA7 [51]. Initial tree(s) for the heuristic search were obtained automatically by applying Neighbor-Join and BioNJ algorithms to a pairwise distance matrix estimated using a JTT model, and then selecting the topology with the best log likelihood value.
2.4. Functionality Test of RepQ, RepY and RepW
Plasmid replication systems of the five ECRs with new replication systems were amplified with the Phusion® High-Fidelity DNA Polymerase, PCR products were cloned into the commercial vector pCR2.1, and the absence of PCR errors was confirmed via Sanger sequencing as previously reported [4]. Primer sequences and the respective size of the PCR products are listed below: (i) RepQ module of the Sulfitobacter indolifex DSM 14862T plasmid pDSM14862_g (P2211: 5′-AGGGTATGGCGACGGTAAAC-3′, P2210: 5′-TAGGGATGGTAGGAGTGGAGAG-3′; 2579 bp), (ii) RepY module of the S. pontiacus DSM 110277 plasmid pDSM110277_f (P2193: 5′-AGCACCAAGCAAGATGATAC-3′, P2194: 5′-GAGCGAGACACCCTTTTTAC-3′; 3423 bp), (iii) RepW module of the S. dubius DSM 109990 plasmid pDSM109990_i (P2206: 5′-GAGGGAAGGGGAGAAGAAAC-3′, P2207: 5′-CATCAACAGCCACAGGATAC-3′; 2350 bp), (iv) RepW module of the Sulfitobacter sp. DSM 110093 plasmid pDSM110093_e (P2208: 5′-TTTGCTCATACCGTTTCTCC-3′, P2209: 5′-CTTTGTTCGCCGTTTTACTC-3′; 2492 bp), and (v) RepW module of the Pseudosulfitobacter sp. DSM 107133 plasmid pDSM107133_l (P2547: 5′-ACGGTGAGTCCCAGATCAGA-3′, P2546: 5′-GCTTTGTTCGGTTTGACGCT-3′; 1992 bp). The functionality of the replicases was tested in the plasmid-cured P. inhibens DSM 17395 Δ65/3 mutant lacking the 65 kb biofilm chromid as previously described [4,55]. The standardized test used the re-isolation of the plasmid construct and its transformation in E. coli to exclude chromosomal integration of the construct, including the kanamycin resistance marker. This procedure provides the final proof for active replication of the construct in P. inhibens.
3. Results and Discussion
3.1. Genome Sequencing and Classification of Five (Pseudo)Sulfitobacter Strains
To generate closed and complete genomes of the (Pseudo)sulfitobacter strains, at least two PacBio HGAP3 assemblies were computed. A reproducible outcome irrespective of the applied assembly parameters, such as the specified genome size, served as an internal control for the reliability of the assembly. Additionally, an Illumina velvet assembly was conducted, and all assemblies were compared to retrieve non-redundant contigs. Both assembly strategies were necessary because smaller plasmids (<10 kb) were often not assembled with PacBio reads due to shearing of the DNA and the size selection of the libraries. For instance, the PacBio assembly of S. dubius DSM 109990 only comprised the chromosome and five extrachromosomal replicons with sizes between 48 and 284 kb (Table 1). Four smaller plasmids ranging from 3.8 to 6.3 kb in size would have been overlooked without the additional Illumina assembly. Subsequent mapping of PacBio sequences on these plasmids revealed a very low number of long reads (Figure S1G–J in comparison to Figure S1A–F), which were insufficient for a de novo assembly, but allowed to confirm the results. On the contrary, the mapping of the Illumina reads even showed that those small plasmids had 10 to 15 times higher copy numbers than the chromosome (Table 1). As both sequencing libraries were prepared from the same DNA extract, small plasmids were obviously quantitatively depleted during the PacBio library preparation. Even high-throughput sequencing (>5000x coverage) without size selection on the novel PacBio Sequel system could not compensate for this effect. The assemblies of medium-sized plasmids also suffered from that depletion. For instance, the 17 kb replicon pDSM14862_g of S. indolifex with 5.3 copies per cell reached only 3% of the chromosomal PacBio coverage (pDSM14862_g: 13x, chromosome: 434x). Altogether, ten plasmids would have been overseen in five strains investigated in the current study if only long-read assemblies had been performed. We conclude that the establishment of high-quality complete bacterial genomes requires a combination of long- and short-read sequencing combined with manual data curation.
All five investigated strains belong to clade 2 of the Rhodobacterales (Figure 1) [56]. Strain DSM 14862T is the type strain of Sulfitobacter indolifex, while the others represent taxonomically uncharacterized isolates. An additional well-supported phylogenomic tree with all related type strains allowed us to identify the closest relative(s) of our four new strains (Figure S2). One Sulfitobacter strain could be classified as S. dubius DSM 109990 (Reference: S. dubius DSM 16472T; digital DNA-DNA hybridization, dDDH; [57]: 70.8%) and one as S. pontiacus DSM 110277 (Reference: S. pontiacus DSM 10014T; dDDH: 77.1%). In contrast, the diagnostic values between strain DSM 110093 and the closest related type strain S. dubius DSM 16472T (dDDH: 50.3%) clearly document that this strain represents a novel species. An analogous conclusion can be drawn for Pseudosulfitobacter sp. DSM 107133 (PIC-76), which forms a distinct branch with Pseudosulfitobacter pseudonitzschiae DSM 26824T (Figure S2), but exhibits a low dDDH value of only 32.3%.
3.2. Characterization of Extrachromosomal Replicons
All five investigated strains comprise multipartite genomes harboring between five and twelve ECRs (Table 1), which is clearly above the average number of about three ECRs that was previously reported for Rhodobacterales ([26] Suppl. 2). Pseudosulfitobacter sp. DSM 107133 carries 12 ECRs and therefore represents together with “Candidatus Marinibacterium anthonyi” La 6, the current record holder in terms of multi-partitioning among roseobacters followed by Marinovum algicola DG898 with eleven ECRs [7,15]. In contrast to La 6, which harbors a large chromosome (5.6 Mbp), all investigated (Pseudo)sulfitobacter strains have chromosomes with a moderate size ranging from 3.0 to 3.6 Mbp. However, the numerous ECRs of Pseudosulfitobacter sp. DSM 107133, especially the 571 kb replicon pDSM107133_a, increased its genome size to 5.2 Mbp, while the genomes of the Sulfitobacter strains have only an average size of about 4 Mbp, which is characteristic for many roseobacters [58]. ECRs above 500 kb are commonly found in Rhizobiales [9,59] but were only sporadically detected in Rhodobacterales, i.e., in the genera Paracoccus, Ruegeria, and now Pseudosulfitobacter [12,60]. Although these ECR are often referred to as megaplasmids, the comparative analysis of the Pseudosulfitobacter sp. DSM 107133 replicons clearly showed that pDSM107133_a has a chromosome-like genetic imprint (Figure S3). Analogous to the 750 kb replicon pAMV3 of P. aminovorans JCM 7685, it can be classified as a chromid that is supposed to carry essential core genes [12,40,41].
The five investigated Sulfitobacter and Pseudosulfitobacter strains contain between one and three chromids (Table 1). Homologs of the sole chromid pDSM14862_c from S. indolifex (CP084954) are also present in the closely related strains S. profundi D7 (CP020695), Sulfitobacter sp. DSM 110093 (CP085170) and S. dubius DSM 109990 (CP085146; Table 1, Figure 1). The four chromids with sizes between 201 and 268 kb share the same RepABC-8 type replication system and exhibit a long-range synteny (Figure S4), which unequivocally documents their common origin. The absence of syntenic homologs in all other strains of Figure 1 supports the following scenario for their origin: (i) A RepABC-8 plasmid was acquired from the common ancestor of S. dubius, Sulfitobacter sp. DSM 110093, S. profundi, and S. indolifex, (ii) it stably co-evolved with the chromosome due to an essential function and (iii) was finally ameliorated into a chromid. Accordingly, this replicon exemplifies the ancient recruitment of a chromid amid the speciation process in the genus Sulfitobacter. Its acquisition antedates, and possibly even triggered the origin of a distinct phylogenetic lineage comprising at least four species (Figure 1).
All chromids of the investigated strains lack genes for their conjugative transfer, which is in agreement with their stable co-evolution with the chromosome. In contrast, mobilization genes are abundant on plasmids and were identified on eight of nine plasmids from Pseudosulfitobacter sp. DSM 107133. Four larger plasmids (>100 kb) of DSM 107133 encode T4SSs and should hence be self-transmissible via conjugation (Table 1), while pDSM107133_d and three cryptic plasmids are probably mobilizable due to the presence of relaxase-encoding MOB genes [20]. The four Sulfitobacter strains harbor five additional plasmids with T4SSs and six with MOB genes. The multipartite genome organization and great abundance of mobilization genes in the five investigated strains support the idea of a huge pan-mobilome of roseobacters serving as a genetic backup in a changing marine environment [61]. It likely also reflects the active role of the genera Sulfitobacter and Pseudosulfitobacter in the network of plasmid exchange in the ocean.
3.3. Identification of Novel Plasmid-Types (RepQ, RepY, RepW)
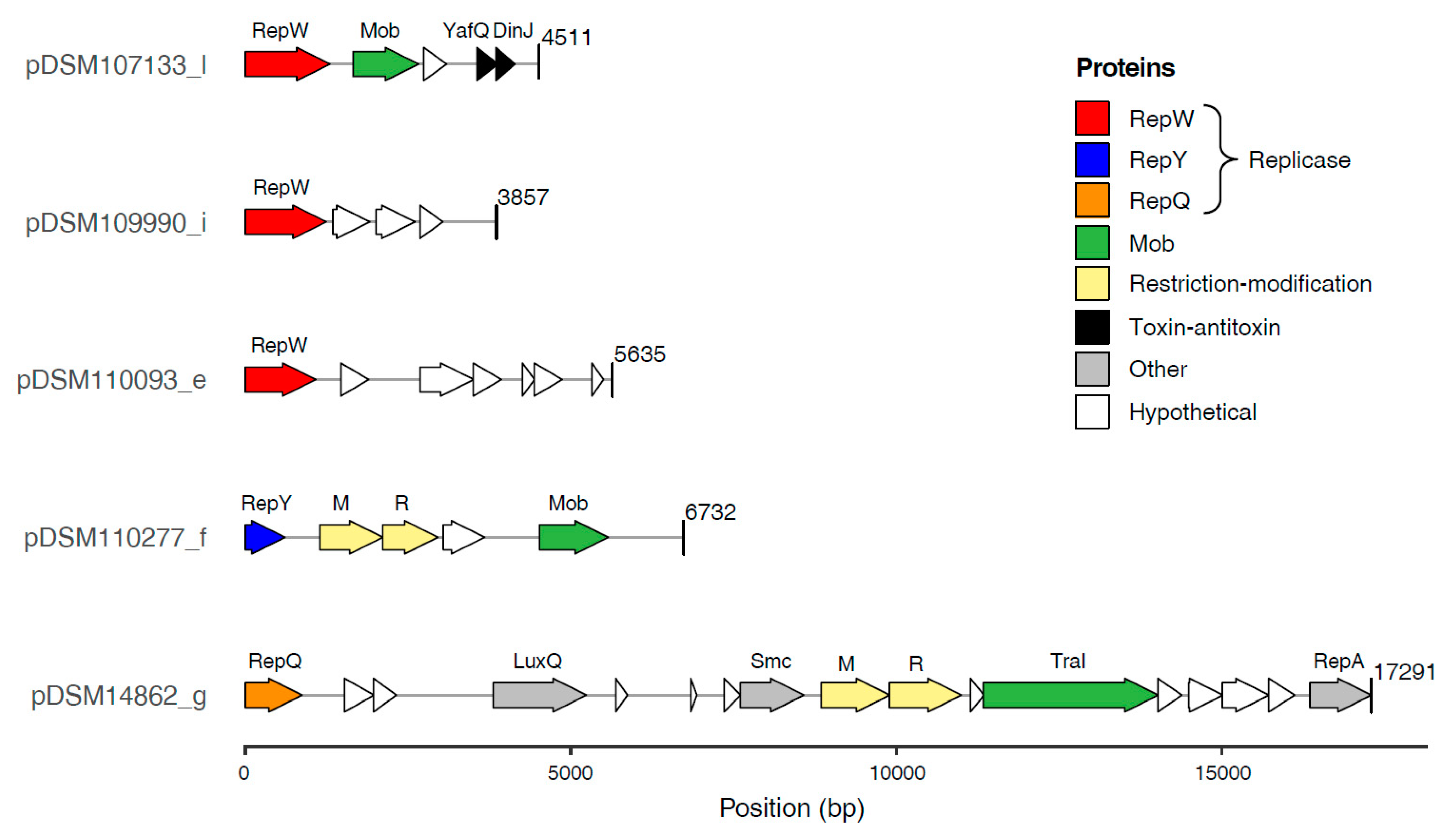
The ECRs of the newly sequenced (Pseudo)sulfitobacter strains encompass the replicases of all known plasmid types (RepA, RepB, RepABC, DnaA-like, RepL) described for roseobacters so far except RepC_soli (Figure 1) [4,7,24]. However, the smallest plasmid of each of the five strains lacks a replication gene that is homologous to these replicases. Comparative genome analyses suggested that the respective plasmids encode three uncharacterized replicases representing novel plasmid types. (i) RepQ: The putative replicase of the 17 kb plasmid pDSM14862_g from S. indolifex DSM 14862T was designated RepQ in reminiscence of homologous replicases on IncQ-type plasmids from Gammaproteobacteria [5,62,63]. A conserved repAQ tandem array is characteristic for these plasmids (Figure 2), and the respective genes of, e.g., the 6388 bp plasmid pHP18 from Aeromonas caviae HP18 were annotated as ‘helicase’ and ‘replication protein,’ respectively (NZ_KU644672.1). (ii) RepY: The putative replicase of pDSM110277_f showed a 68% sequence identity to the functional replication protein of the Paracoccus marcusii OS22 plasmid pMOS6 [25]. A naming according to the conserved Pfam protein domain ‘RepL’ from Firmicutes (PF05732) would be misleading due to the lack of any sequence homology with the recently described RepL-type plasmid replicase from Rhodobacterales [7]. Therefore, we named the novel replicase ‘RepY’ to indicate its unique evolutionary origin and distinguish the corresponding plasmids from all other plasmid types of Rhodobacterales. (iii) RepW: The putative replicase of the plasmids pDSM107133_l, pDSM109990_i, and pDSM110093_e was designated RepW. It showed homology to the Pfam protein family Rep_1 (PF01446) that initiates a rolling circle replication [64].
In accordance with other plasmids lacking a partitioning system, the new plasmid types are characterized by an increased copy number (Table 1). The highest number of 124 copies was observed for the RepY plasmid from S. pontiacus (pDSM110277_f), which is roughly equivalent to the copy number of cloning vectors in E. coli [4]. Moreover, this high copy number plasmid also has the lowest GC content of all investigated replicons (52.1%) and the largest difference to the respective chromosome (Δ8.4%). A comparable observation was made for the novel RepQ and RepW-type plasmids, whose GC content is at least 5% lower than those of the chromosome (Table 1). It is quite common that ‘intracellular genetic parasites’ such as cryptic plasmids, phages, and insertion sequences tend to be AT rich [65]. As G and C are less available and energetically more expensive, a reduced GC content makes these elements less expensive to carry for the host. Therefore, it might be selectively favored and may also promote higher copy numbers [40,65,66]. However, the observed difference may alternatively be explained by the horizontal acquisition of the RepQ, RepY, and RepW plasmids from phylogenetically distinct hosts with a lower GC content.
3.4. Characterization of RepQ, RepY and RepW-Type Plasmids
3.4.1. Functionality of the Three Novel Plasmid Replicases
Plasmid replication of the putative replicases RepQ, RepY, and RepW were investigated in the model organism Phaeobacter inhibens DSM 17395 with our established functionality test [4,7]. We amplified the five newly discovered replication genes, including a 5′ upstream region of at least 450 bp and a 3′ downstream region of at least 100 bp, which should contain all essential cis-acting elements for plasmid replication, and cloned the PCR products into the E. coli vector pCR2.1. Successful transformation and replication in P. inhibens DSM 17395 verified the functionality of all five constructs (Figure S5) since the empty pCR2.1 vector generally does not replicate in Alphaproteobacteria (Petersen et al., 2019). The isolation of the constructs from P. inhibens and their subsequent successful re-transformation into E. coli provided the final proof of their autonomous replication in Rhodobacterales. Accordingly, we could document the functionality of the replicase RepQ from S. indolifex DSM 14862T, RepY from S. pontiacus DSM 110277, and three different RepW-type replicases from S. dubius DSM 109990, Sulfitobacter sp. DSM 110093, and Pseudosulfitobacter sp. DSM 107133.
3.4.2. Gene Content of the Novel Plasmids
Despite its small size, the 17 kb RepQ plasmid pDSM14862_g of S. indolifex is obviously not cryptic. It encodes the quorum-sensing autoinducer 2 sensor kinase/phosphatase LuxQ (Figure 2, Table S2), which regulates biofilm formation in Vibrio [67,68]. Beyond the conserved repAQ replication module, it contains several genes for plasmid-related functions such as the relaxase TraI, which is part of the MOBP family, that likely mediates the mobilization of the replicon [69]. Another protein, which harbours a DNA-binding domain (Pfam: PF11740) and was annotated as chromosome partition protein Smc (HAMAP signature: MF_01894), might be involved in plasmid partitioning [70]. However, the calculated number of 21 plasmid copies per cell (Table 1) indicates that it has at most an auxiliary function for the RepQ plasmid [17]. The gene located downstream of smc, DSM14862_04460 (Figure 2, Table S2), is homologous to the EcoRI methylase from E. coli (P00472; 52% aa identity) [71], which methylates (6-methyladenine) the second adenine of the palindromic sequence GAATTC. However, our PacBio sequencing data clearly documented that this motif is not methylated in S. indolifex DSM 14862T. Besides the 6-methyladenine modification motifs GANTC and RGATCY, which were detected in all Sulfitobacter genomes sequenced in the current study, the motif GGCATC was exclusively identified in strain DSM 14862T. This pattern has no match in ‘REBASE’ [72], so it remains to be investigated if its methylation is catalyzed by the gene product of DSM14862_04460. A corresponding restriction enzyme could not be unequivocally identified, but the adjacent gene (DSM14862_044601) contained an HNH endonuclease Pfam domain (PF01844.23), suggesting that the gene pair may represent a novel restriction-modification system.
The 7 kb RepY plasmid pDSM110277_f of S. pontiacus is a cryptic high copy number plasmid encoding only four genes beyond its eponymous replicase, i.e., a mobilization protein, a hypothetical protein, and a type II restriction-modification module (Figure 2). The respective methylation protein is annotated as ‘DNA (cytosine-5-)-methyltransferase’ (EC 2.1.1.37), but the corresponding modification motif has not yet been identified. The PacBio coverage (97x) was sufficient to detect two 6-methyladenine modification motifs mentioned above, but the detection of 5-methylcytosine requires a much higher sequencing depth (~250x). The adjacent gene encodes an NgoMIV family type II restriction enzyme [72], but the phylogenetic distance to well-characterized homologs allows no conclusion about the palindromic recognition site (EEZ47687: 56% aa identity). However, the presence of a comparable type II restriction-modification module on the cryptic plasmid pAES2 of Paracoccus aestuarii DSM 19484T [25] is indicative of a functional role as a bacterial addiction module using post-segregational killing as a protection mechanism against plasmid loss [73,74].
The three RepW type replicons also represent small cryptic plasmids with a size of less than 6 kb, which carry only three to six annotated genes besides their replicase (Figure 2, Table S2). Two of the RepW plasmids only contain hypothetical proteins without a known function, but the RepW plasmid of Pseudosulfitobacter sp. (pDSM107133_l) harbors a MOB gene indicating its transferability. pDSM107133_l also encodes the mRNA interferase toxin YafQ that inhibits protein translation and the corresponding antitoxin DinJ, whose mode of toxicity has been investigated in E. coli [75,76]. Plasmid encoded toxin-antitoxin modules represent another type of selfish genetic element that acts as an addiction module and ensures the maintenance of the mobile replicons [18]. The recent discovery of a plasmid-encoded toxin-antitoxin system that directly controls the plasmid copy number provides a new perspective on the functional role of these widespread modules for plasmid biology [77].
3.5. Distribution, Evolution and Function of the New Plasmid Types
3.5.1. Presence of RepQ, RepY and RepW in Closed Rhodobacterales Genomes
The distribution of the major replicon types in our newly sequenced strains is representative of copiotrophic Rhodobacterales (Figure 1). In the set of 41 complete genomes, the RepABC type (86) was the most abundant, followed by RepA (34), DnaA-like (28), and RepB-type modules (24). These types were widely distributed among the different Rhodobacterales clades and even present in the distinct clade 8 comprising inter alia the genera Paracoccus and Rhodobacter. In contrast, RepC_soli, RepL, and RepW-type plasmids occurred only occasionally in the collection of 41 genome sequenced Rhodobacterales and RepQ, and RepY-type plasmids were only detected in a single Sulfitobacter strain. The sporadic and scattered distribution of the newly discovered RepQ, RepY, and RepW-type plasmids likely reflects their generally sparse occurrence within Rhodobacterales. However, as closed genomes only account for a small portion of genomes available at NCBI, we searched for all homologous replicases in the NCBI protein database. An extrachromosomal localization of several replicases could be confirmed, partially via manual circularization. Sequence sampling for phylogenetic maximum likelihood analyses, which was conducted with BLASTP searches, was aimed to identify (i) the closest homologs of the replicases (Figure 3, Figure 4 and Figure 5) and (ii) circular plasmids from more distantly related taxa (e.g., Gammaproteobacteria, Figures S6 and S7).
3.5.2. RepQ-Type Plasmids
Comprehensive BLASTP searches revealed a very rare occurrence of RepQ type plasmids in Rhodobacterales (Figure 3). Apart from S. indolifex DSM 14862T, the replicase has only been identified in two Roseovarius sp. strains that were both isolated from the brown alga Ectocarpus subulatus. On the other hand, RepQ type plasmids are frequently found in different orders of Gammaproteobacteria, yet are missing in other alphaproteobacterial lineages. The closely related gammaproteobacterial sister lineage of the three Rhodobacterales replicases comprise small (5 to 10 kb) RepQ type plasmids, which were identified in Oceanospirillales (Halomonas massiliensis Marseille-P2426T), Alteromonadales (Marinobacter antarcticus CGMCC 1.10835), Enterobacterales (e. g., Escherichia coli LMLEEc034) and Pseudomonadales (Pseudomonas cremoris WS 5096; Figure 3, Table S3A). Like pDSM14862_g, all plasmids comprise mobilization genes as well as the characteristic repAQ cassette. The RepQ homologs sampled from Betaproteobacteria are always located on the chromosome (Table S3A), which might reflect an alternate function beyond plasmid replication in this bacterial class. However, we confirmed an extrachromosomal localization for the majority of RepQ proteins (20/34), and the size of the respective plasmids ranges from 5 to 53 kb. The small 5 kb replicons of P. cremoris and Salmonella enterica might represent cryptic plasmids, but virtually all other ECRs comprise additional genes.
The abundance of mobilization genes on RepQ type plasmids and the presence of identical replicases in different Gammaproteobacteria provide strong evidence that these replicons are potent mediators of horizontal gene transfer (HGT; Figure S6). A prime example for trans-order HGT is the nearly identical 7212 bp plasmids of Aeromonas caviae AB5 (Aeromonadales) and Enterobacter roggenkampii IPM1H6 (Enterobacterales) that differ in only one SNP. The presence of a qnrS2 quinolone resistance gene, which was also identified on ten other ECRs analyzed in the current study (Table S3A), emphasizes the relevance of these plasmids for the horizontal spread of antibiotic resistance. The respective quinolone-resistant strains have been isolated from municipal wastewater treatment facilities in Germany and Israel [63,78] as well as rivers near hospitals and aquaculture in China [5] and other environments. The relevance of genetic recombination is exemplified for the 8789 bp plasmid pKA2Q of Kluyvera sp. KA2 that shares a nearly identical backbone of 6 kb with the plasmid pHP5 from Aeromonas allosaccharophila (5915/5927; 99.8% identity) but with the qnrS2 gene replaced by a blaFox gene conferring resistance against cephalosporin antibiotics. Some qnrS2-containing RepQ plasmids (Escherichia coli pUR19829-KPC21 and Aeromonas taiwanensis p1713-KPC) acquired an additional resistance gene against carbapenems (blaKPC-21), which are used as antibiotics of last resort [62].
The discovery of the functional RepQ type plasmid pDSM14862_g in S. indolifex DSM 14862T expanded the host range of these promiscuous ECRs from Gammaproteobacteria to Alphaproteobacteria. The structural conservation of the plasmid backbone indicates that this replicon can be transferred between Rhodobacterales. Accordingly, it might mediate rapid environmental adaptations as previously shown for the small mobilizable RepL type chromate resistance plasmid of “Candidatus Marinibacterium anthonyi” DSM 107130 [7].
3.5.3. RepY-Type Plasmids
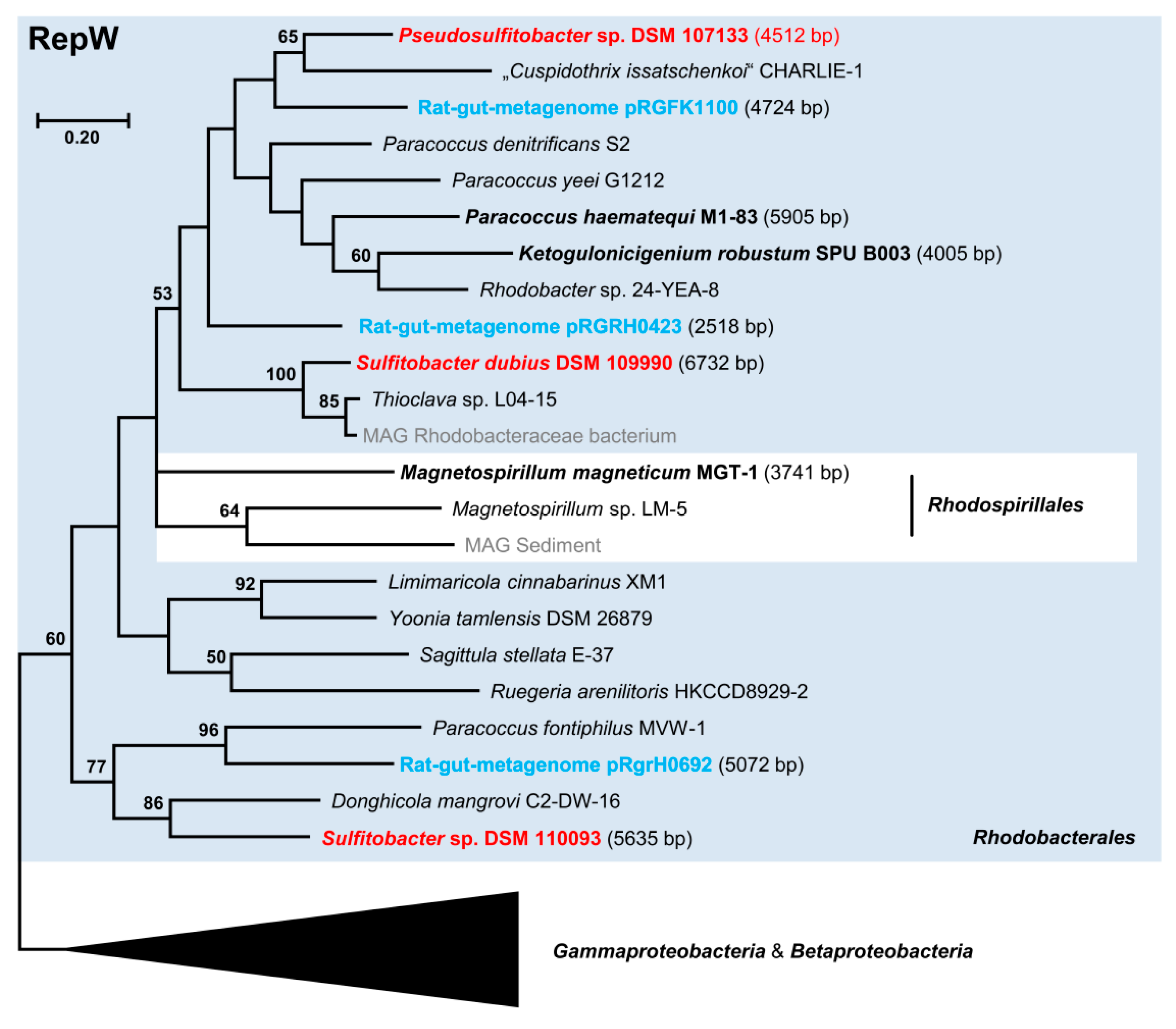
The functionality of the novel replicase has been shown for S. pontiacus DSM 110277 (current study) and for P. marcusii OS22 [25], which represent a typical marine and non-marine Rhodobacterales strain, respectively. RepY proteins, which were present in eleven of about 3000 sequenced Rhodobacterales genomes, form a distinct alphaproteobacterial subtree with a basal positioning of S. pontiacus DSM 110277 (Figure 4). All other RepY proteins were found in non-marine taxa of clade 8 represented by the genera Paracoccus, Rhodobacter, and (Falsi-)Gemmobacter (Table S3B), which indicates that RepY-type plasmids are of minor relevance for the mobilome of the Roseobacter group. However, the discovery of the cryptic plasmid pDSM110277_f in the genus Sulfitobacter is notable due to its exceptional abundance (Table 1). With a calculated number of 124 copies per chromosome equivalent, it represents the first natural high copy number plasmid that has been discovered in roseobacters. Horizontal transfer of the alphaproteobacterial RepY-type plasmids is reflected by the nested positioning of Bartonella sp. HY038 (Hyphomicrobiales, Bartonellaceae), which probably recruited its 15 kb plasmid from the genus Paracoccus. This conclusion is supported by the presence of mobilization genes, e.g., on the plasmids of Sulfitobacter (Figure 2) and P. marcusii [25].
The alphaproteobacterial RepY replicons are generally small (6.5 to 15.0 kb). Those below 10 kb are supposed to be cryptic, but the phylogenetic sister group with beta- and gammaproteobacterial RepY proteins represents ECRs with sizes of up to 45 kb (Figure 4). Three plasmids of Pseudomonas savastanoi pv. savastanoi NCPPB 3335 were investigated for their role for tumor induction in olive plants [79]; the 42 kb RepY-type plasmid pPsv48C is not required for pathogenesis, but it encodes a putative isopentenyl-diphosphate delta-isomerase catalyzing a key step in isoprenoid biosynthesis. The essential gene phzF for the biosynthesis of phenazines, which represent heterocyclic nitrogen-containing metabolites with antibiotic and antitumor activity [80], was found on the RepY plasmid SGTM_pl1 of the betaproteobacterium Sulfuriferula nivalis SGTM. These extrachromosomal genes illustrate the relevance of RepY-type plasmids for Proteobacteria.
3.5.4. RepW-Type Plasmids
The presence of three cryptic RepW-type plasmids in different (Pseudo)sulfitobacter strains, which were discovered in the current study, reflects a wider distribution among Rhodobacterales than RepQ- and RepY-type replicons (Figure 1). This novel plasmid type is mainly found in Alpha-, Beta-, and Gammaproteobacteria, and most alphaproteobacterial sequences form a distinct subtree (Figure 5 and Figure S7). In contrast to RepY, RepW replicases were detected in several clades of the marine Roseobacter group represented, e.g., by the genera Ruegeria (clade 1), Sulfitobacter (clade 2), Sagittula (clade 3), Yoonia (clade 4), and Donghicola (clade 8; Figure 5; Table S3C). The RepW proteins of the three (Pseudo)sulfitobacter plasmids are only distantly related, reflecting the diversity and horizontal exchange of RepW plasmids in Rhodobacterales.
We identified 25 circular RepW-type plasmids with a median size of only 2.6 kb (Table S3C). Although these plasmids are very small (2.2–6.8 kb), it is rather unlikely that they have been systematically overlooked in sequenced bacteria because most of the available genomes are based on Illumina shotgun assemblies. Nevertheless, the identification of twelve circular plasmids from the rat-gut metamobilome documents a great sampling gap of cryptic RepW-type plasmids (Figure 5, Figure S7). The sequences were determined from the cecal content of a dozen rats from Danish hospital sewers and the Falkland Islands [81], following up a seminal pilot study that was aimed to identify complete small plasmids in metagenome datasets [82]. Metamobilomics of a single animal intestine microbiome resulted in nearly the doubling (12/25) of the investigated circular(ized) RepW-type replicons, thus providing a first glimpse into the tremendous under-sampling of plasmids from natural habitats.
The characterization of the small cryptic RepW-type plasmids pTJ86-1 and pTJ86-2 from Cuprividus taiwanensis TJ86 (Betaproteobacteria, Burkholderiales; Figure S7, Table S3C) showed that these replicons are using a rolling-circle replication [83]. The derived shuttle vector pS4-tetR was suitable for the transformation of the genera Burkholderia and Cupriavidus. Another example is the cryptic RepW plasmid pMGT of Magnetospirillum magneticum MGT-1 (Alphaproteobacteria, Rhodospirillales) that served as a backbone for the construction of the shuttle vector pUMG [84], which showed stability, a high copy number of about 40 plasmids per cell and allowed a higher gene expression of the tested reporter-luciferase construct. The novel 4511 bp RepW-type plasmid pDSM107133_l from Pseudosulfitobacter sp. DSM 107133 might be suitable for the development of a new cloning vector for Rhodobacterales because it already harbors a mob gene and a toxin-antitoxin system (Figure 2).
4. Conclusions
Current high throughput DNA sequencing technologies make it possible to decipher a complete bacterial genome for the price of a good bottle of wine, and they pave the way for the systematic characterization of ECRs. Based on the results of the present study, a combination of long-read and short-read sequencing is required to establish genomes of the highest quality encompassing all replicons and ideally lacking any sequencing errors. Conclusions about ECRs in the environment that were formerly drawn from a mobilization-dependent capturing of resistance plasmids from soil samples by conjugation into new host bacteria [3] or from metagenomic analyses of the gut mobilome [82] can now be complemented by holistic insights from an organismic perspective. A good starting point for understanding the organization and evolution of multipartite genomes is the differentiation between plasmids and chromids [40]. The classification of Rhodobacterales ECRs based on their crucial replicases allows to understand the individual genome architecture of a single strain, but it moreover provides first clues about commonalities and differences shared between related taxa. A prime example is the identification of a syntenic RepABC-8 chromid with a size of about 200 kb that is commonly shared by four closely related Sulfitobacter species (S. dubius, Sulfitobacter sp. DSM 110093, S. profundi, S. indolifex) but missing in the genomes of all more distantly related species of this genus. The acquisition of a new chromid is a unique and diagnostic event for the reconstruction of bacterial evolution. Its detection provides the promising perspective to identify key genes that once triggered bacterial speciation via the conquest of a novel ecological niche.
The central finding of the current study was the discovery of the three novel plasmid systems RepQ, RepY, and RepW based on a systematic assessment of all ECRs from five completely sequenced strains. The analyzed replicons are small cryptic, medium to high copy number plasmids without an obvious advantage for the bacterial host. However, they encode replication, stability, and mobility genes as plasmid backbone and could acquire beneficial genes that can easily be exchanged across species and genus borders [7,85]. These replicons might also promote evolutionary innovation as it was observed that multi-copy plasmids allow bacteria to escape from fitness trade-offs [86]. The wealth of different plasmid systems characterized in the current study also provides a promising perspective to develop new molecular tools for biotechnological applications in Rhodobacterales. Genetic complementation of roseobacters is usually performed with the medium copy number broad host range pBBR1MCS vector series [87,88], but neither low nor high copy number vectors are yet available. RepABC-derived vectors would be ideal for applications where a single gene copy per cell is required [89]. In contrast, the RepY-type plasmid from S. pontiacus DSM 110277 seems to be predestined to develop a high copy number vector for protein expression in Rhodobacterales.
Supplementary Materials
The following supporting information can be downloaded at: https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/microorganisms10040738/s1, Figure S1: Mapping of PacBio and Illumina reads on replicons of Sulfitobacter dubius DSM 109990. For larger replicons only the first 25 kb are shown; Figure S2: Phylogenomic UBCG tree based on (draft-) genomes the five newly sequenced strains (red) and all available type strains of the genera Sulfitobacter, Roseobacter and Pseudosulfitobacter; Figure S3: Dendrograms of the relative synonymous codon usage (CU) and tetranucleotide frequency composition (Tetra) analyses from the five newly sequenced Sulfitobacter and Pseudosulfitobacter strains for the differentiation between chromids and plasmids. The well-characterized genome of Dinoroseobacter shibae DSM 16493T was used as a reference for internal calibration; Figure S4: Visualization of synthenic RepABC-8 chromids with Mauve; Figure S5: Functionality tests of RepQ, RepY and RepW plasmids. Restriction and PCR assays for the detection of the pCR2.1-construct isolated from (i) E. coli [lane 1], (ii) P. inhibens DSM 17395 65/3 [lane 2] and (iii) E. coli re-transformed with the P. inhibens plasmid [lane 3]. Upper gel: Plasmid restriction assay with EcoRI resulting in the 3909 bp vector band and one to three insert bands. M, marker (1 kb Plus DNA ladder, Invitrogen). Lower gel: PCR assay with specific primers of the respective replication system: (A) RepQ–S. indolifex DSM 14862T (P2210/P2211), 2579 bp; (B) RepY–S. pontiacus DSM 110277 (P2193/P2194), 3423 bp; (C) RepW–S. dubius DSM 109990 (P2206/P2207), 2350 bp; RepW–Sulfitobacter sp. DSM 110093 (P2208/P2209), 2492 bp; RepW–Pseudosulfitobacter sp. DSM 107133 (P2546/P2547), 1992 bp; Figure S6: Complete tree of the phylogenetic Maximum Likelihood analysis of 34 RepQ type plasmid replication proteins shown in Figure 3. The reference replicase of Sulfitobacter indolifex DSM 14862T is shown in orange, replicases from metagenome-assembled genomes (MAGs) are shown in gray. Circular plasmids or circularized contigs are highlighted in bold. Sequences from Rhodobacterales are highlighted by a blue box. Plasmid names and antibiotic resistance genes are highlighted in red, respectively; Figure S7: Complete tree of the phylogenetic Maximum Likelihood analysis of 61 RepW type plasmid replication proteins shown in Figure 5. Reference replicases of the current study are shown in red, replicases from metagenome-assembled genomes (MAGs) are shown in gray and rat-gut metamobilome-derived sequences are shown in light blue. Circular plasmids or circularized contigs are highlighted in bold. Sequences from Rhodobacterales are highlighted by a blue box. Plasmid names are highlighted in dark blue; Table S1: Overview of 41 Rhodobacterales strains shown in Figure 1. An asterisk in the column ECR_sum (*) indicates that one of the ECRs contained only a putative replicase, whose function has not been tested in Rhodobacterales. # The correct taxonomic name is Cereibacter sphaeroides 2.4.1.; Table S2: Annotation of genes located on the five novel RepW, RepY and RepW plasmids identified in the current study. RepW, RepY and RepQ replication proteins are highlighted in red, blue and orange, respectively; Table S3A: RepQ-type plasmid replicases. Homologous replicases of RepQ with 289 amino acids from the 17,291 bp plasmid from Sulfitobacter indolifex DSM 14862 (orange) were mainly identified in Gammaproteobacteria. MAG, metagenome-assembled genome; yes*, circularized plasmid; QnrS2, quinolone resistance protein; BlaKPC-21, carbapenem resistance protein; BlaFOX, FOX family cephalosporin-hydrolyzing class C beta-lactamase. The presence of antibiotic resistance genes is highlighted in red. B: RepY-type plasmid replicases. Homologous replicases of RepY with 202 amino acids from the 6732 bp plasmid pDSM110277_f from Sulfitobacter pontiacus DSM 110277 (blue). yes*, circularized plasmid. C: RepW-type plasmid replicases. Homologous replicases of RepW with 432 amino acids from the 4511 bp plasmid pDSM107133 from Pseudosulfitobacter sp. DSM 107133. MAG, metagenome-assembled genome; yes*, circularized plasmid. RepW plasmids established in the current study, derived from the rat-gut-metagenome and other MAGs are highlighted in red, blue and gray, respectively.
Author Contributions
J.P. and H.M.F. developed the specific concept of the study within a larger project designed by H.M.F. and J.O. Laboratory work on plasmid biology was carried out by V.R. Genome assembly and data deposition was performed by H.M.F., further analyses were carried out by H.M.F. and J.P. The manuscript was written by H.M.F. and J.P. supported by contributions of J.O. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the Deutsche Forschungsgemeinschaft (DFG, German Research Foundation)—Project-ID 34509606–TRR 51.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Genome sequences are deposited in NCBI GenBank under the accession numbers: CP084951-58, CP084959-65, CP085144-53, CP085154-66 and CP085167-72.
Acknowledgments
We would like to thank Sara Billerbeck and Irene Wagner Döbler, who provided the strains SO248Ex84 and PIC-76, Cendrella Lepleux for supplying sediment samples, and Anika Methner, Orsola Päuker, Franziska Burkart and Nicole Heyer for excellent technical assistance in isolating, cloning, Illumina and PacBio sequencing. We also thank Lukas Birmes, Cathrin Spröer, Boyke Bunk and Henner Brinkmann for further support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Smalla, K.; Jechalke, S.; Top, E.M. Plasmid detection, characterization and ecology. Microbiol. Spectr. 2015, 3, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Elsas, J.D.; Bailey, M.J. The ecology of transfer of mobile genetic elements. FEMS Microbiol. Ecol. 2002, 42, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Heuer, H.; Smalla, K. Plasmids foster diversification and adaptation of bacterial populations in soil. FEMS Microbiol. Rev. 2012, 36, 1083–1104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birmes, L.; Freese, H.M.; Petersen, J. RepC_soli: A Novel Promiscuous Plasmid Type of Rhodobacteraceae Mediates Horizontal Transfer of Antibiotic Resistances in the Ocean. Environ. Microbiol. 2021, 23, 5395–5411. [Google Scholar] [CrossRef]
- Wen, Y.; Pu, X.; Zheng, W.; Hu, G. High prevalence of plasmid-mediated quinolone resistance and IncQ plasmids carrying qnrS2 gene in bacteria from rivers near hospitals and aquaculture in China. PLoS ONE 2016, 11, e0159418. [Google Scholar] [CrossRef]
- Carattoli, A. Resistance plasmid families in Enterobacteriaceae. Antimicrob. Agents Chemother. 2009, 53, 2227–2238. [Google Scholar] [CrossRef] [Green Version]
- Petersen, J.; Vollmers, J.; Ringel, V.; Brinkmann, H.; Ellebrandt-Sperling, C.; Spröer, C.; Howat, A.M.; Murrell, J.C.; Kaster, A.K. A marine plasmid hitchhiking vast phylogenetic and geographic distances. Proc. Natl. Acad. Sci. USA 2019, 116, 20568–20573. [Google Scholar] [CrossRef] [Green Version]
- Dziewit, L.; Pyzik, A.; Szuplewska, M.; Matlakowska, R.; Mielnicki, S.; Wibberg, D.; Schluter, A.; Puhler, A.; Bartosik, D. Diversity and role of plasmids in adaptation of bacteria inhabiting the Lubin copper mine in Poland, an environment rich in heavy metals. Front. Microbiol. 2015, 6, 152. [Google Scholar] [CrossRef] [Green Version]
- Pérez Carrascal, O.M.; VanInsberghe, D.; Juárez, S.; Polz, M.F.; Vinuesa, P.; González, V. Population genomics of the symbiotic plasmids of sympatric nitrogen-fixing Rhizobium species associated with Phaseolus vulgaris. Environ. Microbiol. 2016, 18, 2660–2676. [Google Scholar] [CrossRef]
- Wang, H.; Tomasch, J.; Michael, V.; Bhuju, S.; Jarek, M.; Petersen, J.; Wagner-Döbler, I. Identification of genetic modules mediating the Jekyll and Hyde interaction of Dinoroseobacter shibae with the dinoflagellate Prorocentrum minimum. Front. Microbiol. 2015, 6, 1262. [Google Scholar] [CrossRef]
- Frank, O.; Michael, V.; Päuker, O.; Boedeker, C.; Jogler, C.; Rohde, M.; Petersen, J. Plasmid curing and the loss of grip—The 65 kb replicon of Phaeobacter inhibens DSM 17395 is required for biofilm formation, motility and the colonization of marine algae. Syst. Appl. Microbiol. 2015, 38, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Czarnecki, J.; Dziewit, L.; Puzyna, M.; Prochwicz, E.; Tudek, A.; Wibberg, D.; Schlüter, A.; Pühler, A.; Bartosik, D. Lifestyle-determining extrachromosomal replicon pAMV1 and its contribution to the carbon metabolism of the methylotrophic bacterium Paracoccus aminovorans JCM 7685. Environ. Microbiol. 2017, 19, 4536–4550. [Google Scholar] [CrossRef] [PubMed]
- Koch, H.; Freese, H.M.; Hahnke, R.L.; Simon, M.; Wietz, M. Adaptations of Alteromonas sp. 76-1 to polysaccharide degradation: A CAZyme plasmid for ulvan degradation and two alginolytic systems. Front. Microbiol. 2019, 10, 504. [Google Scholar] [CrossRef] [PubMed]
- Petersen, J.; Brinkmann, H.; Bunk, B.; Michael, V.; Päuker, O.; Pradella, S. Think pink: Photosynthesis, Plasmids and the Roseobacter Clade. Environ. Microbiol. 2012, 14, 2661–2672. [Google Scholar] [CrossRef]
- Frank, O.; Göker, M.; Pradella, S.; Petersen, J. Ocean’s Twelve: Flagellar and Biofilm Chromids in the Multipartite Genome of Marinovum Algicola DG898 Exemplify Functional Compartmentalization. Environ. Microbiol. 2015, 17, 4019–4034. [Google Scholar] [CrossRef]
- Rodriguez-Beltran, J.; DelaFuente, J.; Leon-Sampedro, R.; MacLean, R.C.; San Millan, A. Beyond horizontal gene transfer: The Role of Plasmids in Bacterial Evolution. Nat. Rev. Microbiol. 2021, 19, 347–359. [Google Scholar] [CrossRef]
- Baxter, J.C.; Funnell, B.E. Plasmid partition mechanisms. Microbiol. Spectr. 2014, 2, PLAS-0023-2014. [Google Scholar] [CrossRef] [Green Version]
- Harms, A.; Brodersen, D.E.; Mitarai, N.; Gerdes, K. Toxins, targets, and triggers: An Overview of Toxin-Antitoxin Biology. Mol. Cell 2018, 70, 768–784. [Google Scholar] [CrossRef] [Green Version]
- Christie, P.J. The Mosaic Type IV Secretion Systems. EcoSal Plus 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Smillie, C.; Garcillán-Barcia, M.P.; Francia, M.V.; Rocha, E.P.C.; de la Cruz, F. Mobility of plasmids. Microbiol. Mol. Biol. Rev. 2010, 74, 434–452. [Google Scholar] [CrossRef] [Green Version]
- Xue, H.; Cordero, O.X.; Camas, F.M.; Trimble, W.; Meyer, F.; Guglielmini, J.; Rocha, E.P.C.; Polz, M.F. Eco-evolutionary dynamics of episomes among ecologically cohesive bacterial populations. mBio 2015, 6, e00552-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sevastsyanovich, Y.R.; Krasowiak, R.; Bingle, L.E.H.; Haines, A.S.; Sokolov, S.L.; Kosheleva, I.A.; Leuchuk, A.A.; Titok, M.A.; Smalla, K.; Thomas, C.M. Diversity of IncP-9 plasmids of Pseudomonas. Microbiology 2008, 154, 2929–2941. [Google Scholar] [CrossRef] [Green Version]
- Petersen, J. Phylogeny and compatibility: Plasmid Classification in the Genomics Era. Arch. Microbiol. 2011, 193, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Petersen, J.; Brinkmann, H.; Berger, M.; Brinkhoff, T.; Päuker, O.; Pradella, S. Origin and evolution of a novel DnaA-like plasmid replication type in Rhodobacterales. Mol. Biol. Evol. 2011, 28, 1229–1240. [Google Scholar] [CrossRef] [Green Version]
- Maj, A.; Dziewit, L.; Czarnecki, J.; Wlodarczyk, M.; Baj, J.; Skrzypczyk, G.; Giersz, D.; Bartosik, D. Plasmids of carotenoid-producing Paracoccus spp. (Alphaproteobacteria)—structure, diversity and evolution. PLoS ONE 2013, 8, e80258. [Google Scholar] [CrossRef] [Green Version]
- Simon, M.; Scheuner, C.; Meier-Kolthoff, J.P.; Brinkhoff, T.; Wagner-Döbler, I.; Ulbrich, M.; Klenk, H.P.; Schomburg, D.; Petersen, J.; Göker, M. Phylogenomics of Rhodobacteraceae reveals evolutionary adaptation to marine and non-marine habitats. ISME J. 2017, 11, 1483–1499. [Google Scholar] [CrossRef] [PubMed]
- Newton, R.J.; Griffin, L.E.; Bowles, K.M.; Meile, C.; Gifford, S.; Givens, C.E.; Howard, E.C.; King, E.; Oakley, C.A.; Reisch, C.R.; et al. Genome characteristics of a generalist marine bacterial lineage. ISME J. 2010, 4, 784–798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fadeev, E.; De Pascale, F.; Vezzi, A.; Hubner, S.; Aharonovich, D.; Sher, D. Why close a bacterial genome? The plasmid of Alteromonas macleodii HOT1A3 is a vector for inter-specific transfer of a flexible genomic island. Front. Microbiol. 2016, 7, 248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, K.Y.H.; Orata, F.D.; Boucher, Y.F.; Case, R.J. Roseobacters in a sea of poly- and paraphyly: Whole Genome-Based Taxonomy of the Family Rhodobacteraceae and the Proposal for the Split of the “Roseobacter Clade” into a Novel Family, Roseobacteraceae fam. nov. Front. Microbiol. 2021, 12, 683109. [Google Scholar] [CrossRef]
- Yang, Q.; Ge, Y.-M.; Iqbal, N.M.; Yang, X.; Zhang, X.-L. Sulfitobacter alexandrii sp. nov., a new microalgae growth-promoting bacterium with exopolysaccharides bioflocculanting potential isolated from marine phycosphere. Antonie Leeuwenhoek 2021, 114, 1091–1106. [Google Scholar] [CrossRef]
- Pohlner, M.; Degenhardt, J.; von Hoyningen-Huene, A.J.E.; Wemheuer, B.; Erlmann, N.; Schnetger, B.; Badewien, T.H.; Engelen, B. The biogeographical distribution of benthic Roseobacter group members along a Pacific transect is structured by nutrient availability within the sediments and primary production in different oceanic provinces. Front. Microbiol. 2017, 8, 2550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Kang, S.-H.; Yang, E.J.; Macdonald, A.M.; Joo, H.M.; Park, J.; Kim, K.; Lee, G.S.; Kim, J.-H.; Yoon, J.-E.; et al. Latitudinal distributions and controls of bacterial community composition during the summer of 2017 in western Arctic surface waters (from the Bering Strait to the Chukchi Borderland). Sci. Rep. 2019, 9, 16822. [Google Scholar] [CrossRef] [PubMed]
- Allgaier, M.; Uphoff, H.; Felske, A.; Wagner-Döbler, I. Aerobic anoxygenic photosynthesis in Roseobacter clade bacteria from diverse marine habitats. Appl. Environ. Microbiol. 2003, 69, 5051–5059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baym, M.; Kryazhimskiy, S.; Lieberman, T.D.; Chung, H.; Desai, M.M.; Kishony, R. Inexpensive multiplexed library preparation for megabases-sized genomes. PLoS ONE 2015, 10, e0128036. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [Green Version]
- Koboldt, D.C.; Zhang, Q.; Larson, D.E.; Shen, D.; McLellan, M.D.; Lin, L.; Miller, C.A.; Mardis, E.R.; Ding, L.; Wilson, R.K. VarScan 2: Somatic Mutation and Copy Number Alteration Discovery in Cancer by Exome Sequencing. Genome Res. 2012, 22, 568–576. [Google Scholar] [CrossRef] [Green Version]
- Zerbino, D.R.; Birney, E. Velvet: Algorithms for de Novo Short Read Assembly Using de Bruijn Graphs. Genome Res. 2008, 18, 821–829. [Google Scholar] [CrossRef] [Green Version]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Harrison, P.W.; Lower, R.P.J.; Kim, N.K.D.; Young, J.P.W. Introducing the bacterial ‘chromid’: Not a Chromosome, not a Plasmid. Trends Microbiol. 2010, 18, 141–148. [Google Scholar] [CrossRef]
- Petersen, J.; Frank, O.; Göker, M.; Pradella, S. Extrachromosomal, extraordinary and essential—The plasmids of the Roseobacter clade. Appl. Microbiol. Biotechnol. 2013, 97, 2805–2815. [Google Scholar] [CrossRef] [PubMed]
- Garcillán-Barcia, M.P.; Redondo-Salvo, S.; Vielva, L.; de la Cruz, F. MOBscan: Automated Annotation of MOB Relaxases. Methods Mol. Biol. 2020, 2075, 295–308. [Google Scholar] [CrossRef]
- Parte, A.C.; Sardà Carbasse, J.; Meier-Kolthoff, J.P.; Reimer, L.C.; Göker, M. List of Prokaryotic names with Standing in Nomenclature (LPSN) moves to the DSMZ. Int. J. Syst. Evol. Microbiol. 2020, 70, 5607–5612. [Google Scholar] [CrossRef]
- Na, S.I.; Kim, Y.O.; Yoon, S.H.; Ha, S.M.; Baek, I.; Chun, J. UBCG: Up-to-date bacterial core gene set and pipeline for phylogenomic tree reconstruction. J. Microbiol. 2018, 56, 280–285. [Google Scholar] [CrossRef] [PubMed]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately Maximum-Likelihood trees for large alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef] [PubMed]
- Meier-Kolthoff, J.P.; Göker, M. TYGS is an automated high-throughput platform for state-of-the-art genome-based taxonomy. Nat. Commun. 2019, 10, 2182. [Google Scholar] [CrossRef] [PubMed]
- Petersen, J.; Brinkmann, H.; Pradella, S. Diversity and evolution of repABC type plasmids in Rhodobacterales. Environ. Microbiol. 2009, 11, 2627–2638. [Google Scholar] [CrossRef]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.L.; Tosatto, S.C.E.; Paladin, L.; Raj, S.; Richardson, L.J.; et al. Pfam: The Protein Families Database in 2021. Nucleic Acids Res. 2020, 49, D412–D419. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple Sequence Alignment with High Accuracy and High Throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Jones, D.T.; Taylor, W.R.; Thornton, J.M. The rapid generation of mutation data matrices from protein sequences. Comput. Appl. Biosci. 1992, 8, 275–282. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Yu, G. Using ggtree to Visualize Data on Tree-Like Structures. Curr. Protoc. Bioinform. 2020, 69, e96. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, D. gggenes: Draw Gene Arrow Maps in ‘ggplot2’, R package version 0.4.1; 2020; Available online: https://wilkox.org/gggenes/ (accessed on 28 February 2022).
- Bartling, P.; Brinkmann, H.; Bunk, B.; Overmann, J.; Göker, M.; Petersen, J. The composite 259-kb plasmid of Martelella mediterranea DSM 17316(T)—A natural replicon with functional RepABC modules from Rhodobacteraceae and Rhizobiaceae. Front. Microbiol. 2017, 8, 1787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartling, P.; Vollmers, J.; Petersen, J. The first world swimming championships of roseobacters—Phylogenomic insights into an exceptional motility phenotype. Syst. Appl. Microbiol. 2018, 41, 544–554. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Auch, A.F.; Klenk, H.P.; Göker, M. Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinform. 2013, 14, 60. [Google Scholar] [CrossRef] [Green Version]
- Luo, H.; Swan, B.K.; Stepanauskas, R.; Hughes, A.L.; Moran, M.A. Evolutionary analysis of a streamlined lineage of surface ocean Roseobacters. ISME J. 2014, 8, 1428–1439. [Google Scholar] [CrossRef] [Green Version]
- Galardini, M.; Pini, F.; Bazzicalupo, M.; Biondi, E.G.; Mengoni, A. Replicon-dependent bacterial genome evolution: The Case of Sinorhizobium Meliloti. Genome Biol. Evol. 2013, 5, 542–558. [Google Scholar] [CrossRef]
- Sonnenschein, E.C.; Nielsen, K.F.; D’Alvise, P.; Porsby, C.H.; Melchiorsen, J.; Heilmann, J.; Kalatzis, P.G.; Lopez-Perez, M.; Bunk, B.; Sproer, C.; et al. Global occurrence and heterogeneity of the Roseobacter-clade species Ruegeria mobilis. ISME J. 2017, 11, 569–583. [Google Scholar] [CrossRef] [Green Version]
- Tomasch, J.; Ringel, V.; Wang, H.; Freese, H.M.; Bartling, P.; Brinkmann, H.; Vollmers, J.; Jarek, M.; Wagner-Döbler, I.; Petersen, J. Fatal Affairs—Conjugational transfer of a Dinoflagellate-killing plasmid between marine Rhodobacterales. Microb. Genom. 2022, 8, 000787. [Google Scholar] [CrossRef]
- Manageiro, V.; Romao, R.; Moura, I.B.; Sampaio, D.A.; Vieira, L.; Ferreira, E.; Canica, M.; Network Eu, S.P. Molecular epidemiology and risk factors of carbapenemase-producing Enterobacteriaceae isolates in Portuguese hospitals: Results from European survey on carbapenemase-producing Enterobacteriaceae (EuSCAPE). Front. Microbiol. 2018, 9, 8. [Google Scholar] [CrossRef] [Green Version]
- Kaplan, E.; Sela, N.; Doron-Faigenboim, A.; Navon-Venezia, S.; Jurkevitch, E.; Cytryn, E. Genomic and functional characterization of qnr-encoding plasmids from municipal wastewater biosolid Klebsiella pneumoniae isolates. Front. Microbiol. 2015, 6, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marsin, S.; Forterre, P. A rolling circle replication initiator protein with a nucleotidyl-transferase activity encoded by the plasmid pGT5 from the hyperthermophilic archaeon Pyrococcus abyssi. Mol. Microbiol. 1998, 27, 1183–1192. [Google Scholar] [CrossRef] [PubMed]
- Rocha, E.P.C.; Danchin, A. Base composition bias might result from competition for metabolic resources. Trends Genet. 2002, 18, 291–294. [Google Scholar] [CrossRef]
- Dietel, A.-K.; Merker, H.; Kaltenpoth, M.; Kost, C. Selective advantages favour high genomic AT-contents in intracellular elements. PLoS Genet. 2019, 15, e1007778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Zhu, X.; Zhang, C.; Zhao, Z. LuxQ-LuxU-LuxO pathway regulates biofilm formation by Vibrio parahaemolyticus. Microbiol. Res. 2021, 250, 126791. [Google Scholar] [CrossRef]
- Miller, M.B.; Skorupski, K.; Lenz, D.H.; Taylor, R.K.; Bassler, B.L. Parallel quorum sensing systems converge to regulate virulence in Vibrio cholerae. Cell 2002, 110, 303–314. [Google Scholar] [CrossRef] [Green Version]
- Byrd, D.R.; Matson, S.W. Nicking by transesterification: The Reaction Catalysed by a Relaxase. Mol. Microbiol. 1997, 25, 1011–1022. [Google Scholar] [CrossRef] [Green Version]
- Jagura-Burdzy, G.; Thomas, C.M. kfrA gene of broad host range plasmid RK2 encodes a novel DNA-binding protein. J. Mol. Biol. 1992, 225, 651–660. [Google Scholar] [CrossRef]
- Greene, P.J.; Gupta, M.; Boyer, H.W.; Brown, W.E.; Rosenberg, J.M. Sequence analysis of the DNA encoding the Eco RI endonuclease and methylase. J. Biol. Chem. 1981, 256, 2143–2153. [Google Scholar] [CrossRef]
- Roberts, R.J.; Vincze, T.; Posfai, J.; Macelis, D. REBASE—A database for DNA restriction and modification: Enzymes, Genes and Genomes. Nucleic Acids Res. 2015, 43, D298–D299. [Google Scholar] [CrossRef]
- Ichige, A.; Kobayashi, I. Stability of EcoRI restriction-modification enzymes in vivo differentiates the EcoRI restriction-modification system from other postsegregational cell killing systems. J. Bacteriol. 2005, 187, 6612–6621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dziewit, L.; Kuczkowska, K.; Adamczuk, M.; Radlinska, M.; Bartosik, D. Functional characterization of the type II PamI restriction-modification system derived from plasmid pAMI7 of Paracoccus aminophilus JCM 7686. FEMS Microbiol. Lett. 2011, 324, 56–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Motiejūnaite, R.; Armalyte, J.; Markuckas, A.; Suziedeliene, E. Escherichia coli dinJ-yafQ genes act as a toxin-antitoxin module. FEMS Microbiol. Lett. 2007, 268, 112–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prysak, M.H.; Mozdzierz, C.J.; Cook, A.M.; Zhu, L.; Zhang, Y.; Inouye, M.; Woychik, N.A. Bacterial toxin YafQ is an endoribonuclease that associates with the ribosome and blocks translation elongation through sequence-specific and frame-dependent mRNA cleavage. Mol. Microbiol. 2009, 71, 1071–1087. [Google Scholar] [CrossRef] [PubMed]
- Ni, S.W.; Li, B.Y.; Tang, K.H.; Yao, J.Y.; Wood, T.K.; Wang, P.X.; Wang, X.X. Conjugative plasmid-encoded toxin-antitoxin system PrpT/PrpA directly controls plasmid copy number. Proc. Natl. Acad. Sci. USA 2021, 118, 10. [Google Scholar] [CrossRef]
- Bönemann, G.; Stiens, M.; Pühler, A.; Schlüter, A. Mobilizable IncQ-related plasmid carrying a new quinolone resistance gene, qnrS2, isolated from the bacterial community of a wastewater treatment plant. Antimicrob. Agents Chemother. 2006, 50, 3075–3080. [Google Scholar] [CrossRef] [Green Version]
- Bardaji, L.; Pérez-Martínez, I.; Rodríguez-Moreno, L.; Rodríguez-Palenzuela, P.; Sundin, G.W.; Ramos, C.; Murillo, J. Sequence and role in virulence of the three plasmid complement of the model tumor-inducing bacterium Pseudomonas savastanoi pv. savastanoi NCPPB 3335. PLoS ONE 2011, 6, e25705. [Google Scholar] [CrossRef] [Green Version]
- Blankenfeldt, W.; Kuzin, A.P.; Skarina, T.; Korniyenko, Y.; Tong, L.; Bayer, P.; Janning, P.; Thomashow, L.S.; Mavrodi, D.V. Structure and function of the phenazine biosynthetic protein PhzF from Pseudomonas fluorescens. Proc. Natl. Acad. Sci. USA 2004, 101, 16431–16436. [Google Scholar] [CrossRef] [Green Version]
- Jørgensen, T.S.; Hansen, M.A.; Xu, Z.; Tabak, M.A.; Sørensen, S.J.; Hansen, L.H. Plasmids, viruses, and other circular elements in rat gut. bioRxiv 2017, 143420. [Google Scholar] [CrossRef] [Green Version]
- Jørgensen, T.S.; Xu, Z.; Hansen, M.A.; Sørensen, S.J.; Hansen, L.H. Hundreds of circular novel plasmids and DNA elements identified in a rat cecum metamobilome. PLoS ONE 2014, 9, e87924. [Google Scholar] [CrossRef] [Green Version]
- Sheu, S.Y.; Chen, W.M.; Lin, G.H. Characterization and application of a rolling-circle-type plasmid from Cupriavidus taiwanensis. Plasmid 2007, 57, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Okamura, Y.; Takeyama, H.; Sekine, T.; Sakaguchi, T.; Wahyudi, A.T.; Sato, R.; Kamiya, S.; Matsunaga, T. Design and application of a new cryptic-plasmid-based shuttle vector for Magnetospirillum magneticum. Appl. Environ. Microbiol. 2003, 69, 4274–4277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Attéré, S.A.; Vincent, A.T.; Paccaud, M.; Frenette, M.; Charette, S.J. The role for the small cryptic plasmids as moldable vectors for genetic innovation in Aeromonas salmonicida subsp. salmonicida. Front. Genet. 2017, 8, 211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez-Beltran, J.; Hernandez-Beltran, J.C.R.; DelaFuente, J.; Escudero, J.A.; Fuentes-Hernandez, A.; MacLean, R.C.; Pena-Miller, R.; Millan, A.S. Multicopy plasmids allow bacteria to escape from fitness trade-offs during evolutionary innovation. Nat. Ecol. Evol. 2018, 2, 873–881. [Google Scholar] [CrossRef] [PubMed]
- Kovach, M.E.; Elzer, P.H.; Steven Hill, D.; Robertson, G.T.; Farris, M.A.; Roop, R.M.; Peterson, K.M. Four new derivatives of the broad-host-range cloning vector pBBR1MCS, carrying different antibiotic-resistance cassettes. Gene 1995, 166, 175–176. [Google Scholar] [CrossRef]
- Patzelt, D.; Wang, H.; Buchholz, I.; Rohde, M.; Gröbe, L.; Pradella, S.; Neumann, A.; Schulz, S.; Heyber, S.; Münch, K.; et al. You are what you talk: Quorum sensing induces individual morphologies and cell division modes in Dinoroseobacter shibae. ISME J. 2013, 7, 2274–2286. [Google Scholar] [CrossRef] [Green Version]
- Döhlemann, J.; Wagner, M.; Happel, C.; Carrillo, M.; Sobetzko, P.; Erb, T.J.; Thanbichler, M.; Becker, A. A family of single copy repABC-type shuttle vectors stably maintained in the alpha-proteobacterium Sinorhizobium meliloti. ACS Synth. Biol. 2017, 6, 968–984. [Google Scholar] [CrossRef]
Figure 1.
Phylogenomic tree of 41 Rhodobacterales strains with closed genomes and distribution of their extrachromosomal replicons (ECRs). The phylogenetic tree was constructed from 30,310 amino acid positions of 92 housekeeping genes. The monophyly of the family Roseobacteraceae and a sister group position of the Rhodobacteraceae (Clade 8) was confirmed by a phylogenomic analysis with the draft genome of the deep branching taxon Neomegalonema perideroedes DSM 15528. The matrix on the right side depicts the number of ECRs classified by their replication system. An asterisk (*) indicates that one of the ECRs contained only a putative replicase, which function has not been tested in Rhodobacterales. Strains sequenced and investigated within this study are colored in accordance with their novel plasmid systems and shown in bold. # The correct taxonomic name is Cereibacter sphaeroides 2.4.1. Genome accession numbers are listed in Table S1.
Figure 1.
Phylogenomic tree of 41 Rhodobacterales strains with closed genomes and distribution of their extrachromosomal replicons (ECRs). The phylogenetic tree was constructed from 30,310 amino acid positions of 92 housekeeping genes. The monophyly of the family Roseobacteraceae and a sister group position of the Rhodobacteraceae (Clade 8) was confirmed by a phylogenomic analysis with the draft genome of the deep branching taxon Neomegalonema perideroedes DSM 15528. The matrix on the right side depicts the number of ECRs classified by their replication system. An asterisk (*) indicates that one of the ECRs contained only a putative replicase, which function has not been tested in Rhodobacterales. Strains sequenced and investigated within this study are colored in accordance with their novel plasmid systems and shown in bold. # The correct taxonomic name is Cereibacter sphaeroides 2.4.1. Genome accession numbers are listed in Table S1.

Figure 2.
Gene content of five (Pseudo)sulfitobacter plasmids representing the novel replicon types RepW, RepY and RepQ. The plasmids were identified in the complete genomes from Pseudosulfitobacter sp. DSM 107133, S. dubius DSM 109990, Sulfitobacter sp. DSM 110093, S. pontiacus DSM 110277 and S. indolifex DSM 14862T (Table 1). The figure was generated with the R package gggene, respective annotations are shown in Table S2.
Figure 2.
Gene content of five (Pseudo)sulfitobacter plasmids representing the novel replicon types RepW, RepY and RepQ. The plasmids were identified in the complete genomes from Pseudosulfitobacter sp. DSM 107133, S. dubius DSM 109990, Sulfitobacter sp. DSM 110093, S. pontiacus DSM 110277 and S. indolifex DSM 14862T (Table 1). The figure was generated with the R package gggene, respective annotations are shown in Table S2.

Figure 3.
Phylogenetic Maximum Likelihood tree of 34 RepQ type plasmid replication proteins based on 266 amino acid positions. The replicase of Sulfitobacter indolifex DSM 14862T is shown in orange, and replicases from metagenome-assembled genomes (MAGs) are shown in gray. Circular plasmids or circularized contigs are highlighted in bold. Sequences from Rhodobacterales are highlighted by a blue box. The complete tree is shown in Figure S6. Accession numbers, localization, and further characteristics of RepQ type plasmids are presented in Table S3A.
Figure 3.
Phylogenetic Maximum Likelihood tree of 34 RepQ type plasmid replication proteins based on 266 amino acid positions. The replicase of Sulfitobacter indolifex DSM 14862T is shown in orange, and replicases from metagenome-assembled genomes (MAGs) are shown in gray. Circular plasmids or circularized contigs are highlighted in bold. Sequences from Rhodobacterales are highlighted by a blue box. The complete tree is shown in Figure S6. Accession numbers, localization, and further characteristics of RepQ type plasmids are presented in Table S3A.

Figure 4.
Phylogenetic Maximum Likelihood tree of 23 RepY type plasmid replication proteins based on 157 amino acid positions. Bootstrap support > 40% is indicated. The replicase of Sulfitobacter pontiacus DSM 110277 is shown in blue. Circular plasmids or circularized contigs are highlighted in bold. Sequences from Rhodobacterales are highlighted by a blue box. Accession numbers, localization, and further characteristics of RepY type replicases are presented in Table S3B.
Figure 4.
Phylogenetic Maximum Likelihood tree of 23 RepY type plasmid replication proteins based on 157 amino acid positions. Bootstrap support > 40% is indicated. The replicase of Sulfitobacter pontiacus DSM 110277 is shown in blue. Circular plasmids or circularized contigs are highlighted in bold. Sequences from Rhodobacterales are highlighted by a blue box. Accession numbers, localization, and further characteristics of RepY type replicases are presented in Table S3B.

Figure 5.
Phylogenetic Maximum Likelihood tree of 61 RepW type plasmid replication proteins based on 123 amino acid positions. Bootstrap support >40% is indicated. The reference replicases of Pseudosulfitobacter sp. DSM 107133, Sulfitobacter dubius DSM 109990, and Sulfitobacter sp. DSM 110093 are shown in red. Circular plasmids or circularized contigs are highlighted in bold. Rat-gut metamobilome-derived sequences are shown in blue, and sequences from MAGs are shown in gray. Sequences from Rhodobacterales are highlighted by a blue box. The complete tree is shown in Figure S7. Accession numbers, localization, and further characteristics of RepW type replicases are presented in Table S3C.
Figure 5.
Phylogenetic Maximum Likelihood tree of 61 RepW type plasmid replication proteins based on 123 amino acid positions. Bootstrap support >40% is indicated. The reference replicases of Pseudosulfitobacter sp. DSM 107133, Sulfitobacter dubius DSM 109990, and Sulfitobacter sp. DSM 110093 are shown in red. Circular plasmids or circularized contigs are highlighted in bold. Rat-gut metamobilome-derived sequences are shown in blue, and sequences from MAGs are shown in gray. Sequences from Rhodobacterales are highlighted by a blue box. The complete tree is shown in Figure S7. Accession numbers, localization, and further characteristics of RepW type replicases are presented in Table S3C.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Replicon characteristics of investigated (Pseudo)sulfitobacter strains with closed genomes.
Table 1.
Replicon characteristics of investigated (Pseudo)sulfitobacter strains with closed genomes.
| Replicon Id | Size (bp) | Replicon Type | GC | Copy No. | Replication Module | Partitioning System | Mobility | NCBI Accession |
|---|---|---|---|---|---|---|---|---|
| Sulfitobacter indolifexDSM 14862 (HEL-45) | ||||||||
| cDSM14862 | 3,271,523 | chromosome | 60.0 | 1.0 | DnaA | yes | no | CP084951 |
| pDSM14862_a | 313,826 | plasmid | 55.4 | 0.5 | RepABC-20 | yes | no | CP084952 |
| pDSM14862_b | 307,297 | plasmid | 56.1 | 0.8 | DnaA-like-I | yes | no | CP084953 |
| pDSM14862_c | 200,719 | chromid | 60.3 | 0.8 | RepABC-8 | yes | no | CP084954 |
| pDSM14862_d | 160,516 | plasmid | 56.5 | 0.8 | DnaA-like-II | yes | no | CP084955 |
| pDSM14862_e | 123,234 | plasmid | 57.1 | 0.6 | RepABC-4 | yes | T4SS | CP084956 |
| pDSM14862_f | 104,164 | plasmid | 59.8 | 0.7 | RepABC-7 | yes | T4SS | CP084957 |
| pDSM14862_g | 17,291 | plasmid | 54.9 | 5.3 | RepQ | no | MOBP | CP084958 |
| Sulfitobacter pontiacusDSM 110277 (SO248Ex84) | ||||||||
| cDSM110277 | 3,012,962 | chromosome | 60.5 | 1.0 | DnaA | yes | no | CP084959 |
| pDSM110277_a | 239,416 | plasmid | 60.2 | 0.7 | RepABC-10 | yes | T4SS | CP084960 |
| pDSM110277_b | 230,345 | chromid | 59.7 | 0.7 | DnaA-like-II | yes | no | CP084961 |
| pDSM110277_c | 177,750 | chromid | 60.3 | 0.6 | DnaA-like-I | yes | no | CP084962 |
| pDSM110277_d | 128,813 | plasmid | 59.6 | 0.5 | RepABC-9-1 | yes | T4SS | CP084963 |
| pDSM110277_e | 53,945 | plasmid | 57.0 | 1.8 | RepABC-8 | yes | no | CP084964 |
| pDSM110277_f | 6732 | plasmid | 52.1 | 124.3 | RepY | no | MOBQ | CP084965 |
| Sulfitobacter sp. DSM 110093 (2RS2_G6) | ||||||||
| cDSM110093 | 3,434,207 | chromosome | 59.7 | 1.0 | DnaA | yes | MOBV | CP085167 |
| pDSM110093_a | 336,544 | plasmid | 55.3 | 1.1 | RepABC-21 | yes | no | CP085168 |
| pDSM110093_b | 273,772 | plasmid | 54.9 | 1.1 | RepABC-20 | yes | no | CP085169 |
| pDSM110093_c | 268,356 | chromid | 59.7 | 1.0 | RepABC-8 | yes | no | CP085170 |
| pDSM110093_d | 21,520 | plasmid | 52.6 | 4.3 | RepA | no | MOBP | CP085171 |
| pDSM110093_e | 5635 | plasmid | 53.9 | 23.4 | RepW | no | no | CP085172 |
| Sulfitobacter dubius DSM 109990 (3RS2_G4b) | ||||||||
| cDSM109990 | 3,274,709 | chromosome | 60.2 | 1.0 | DnaA | yes | no | CP085144 |
| pDSM109990_a | 284,454 | plasmid | 55.4 | 1.2 | RepB-I | yes | MOBP | CP085145 |
| pDSM109990_b | 247,035 | chromid | 60.5 | 0.9 | RepABC-8 | yes | no | CP085146 |
| pDSM109990_c | 183,486 | plasmid | 57.6 | 0.9 | RepABC-2 | yes | no | CP085147 |
| pDSM109990_d | 108,277 | plasmid | 59.9 | 0.6 | RepABC-1 | yes | T4SS | CP085148 |
| pDSM109990_e | 47,721 | plasmid | 58.7 | 2.6 | RepA_a | no | MOBQ | CP085149 |
| pDSM109990_f | 6286 | plasmid | 59.7 | 9.5 | RepA_b | no | MOBQ | CP085150 |
| pDSM109990_g | 4878 | plasmid | 52.3 | 12.5 | RepA_c | no | no | CP085151 |
| pDSM109990_h | 4609 | plasmid | 58.4 | 12.4 | RepA_d | no | MOBV | CP085152 |
| pDSM109990_i | 3857 | plasmid | 55.0 | 15.3 | RepW | no | no | CP085153 |
| Pseudosulfitobacter sp. DSM 107133 (PIC-76) | ||||||||
| cDSM107133 | 3,635,847 | chromosome | 61.1 | 1.0 | DnaA | yes | no | CP085154 |
| pDSM107133_a | 571,401 | chromid | 61.0 | 1.1 | RepABC-3 | yes | no | CP085155 |
| pDSM107133_b | 246,683 | chromid | 61.1 | 0.6 | RepB-I | yes | no | CP085156 |
| pDSM107133_c | 158,961 | plasmid | 60.3 | 0.5 | RepABC-11 | yes | T4SS | CP085157 |
| pDSM107133_d | 135,500 | plasmid | 58.0 | 0.4 | RepABC-9-1 | yes | MOBP | CP085158 |
| pDSM107133_e | 129,047 | plasmid | 58.4 | 0.4 | RepABC-5 | yes | T4SS | CP085159 |
| pDSM107133_f | 128,511 | plasmid | 60.2 | 0.4 | RepABC-1 | yes | T4SS | CP085160 |
| pDSM107133_g | 105,694 | plasmid | 60.4 | 0.4 | RepABC-9-2 | yes | T4SS | CP085161 |
| pDSM107133_h | 65,426 | chromid | 63.6 | 0.6 | RepB-III | yes | no | CP085162 |
| pDSM107133_i | 17,449 | plasmid | 53.1 | 3.8 | RepA_a | no | no | CP085163 |
| pDSM107133_j | 7190 | plasmid | 59.6 | 9.1 | RepA_b | no | MOBQ | CP085164 |
| pDSM107133_k | 5670 | plasmid | 59.0 | 8.6 | RepL | no | MOBQ | CP085165 |
| pDSM107133_l | 4511 | plasmid | 53.5 | 21.3 | RepW | no | MOBV | CP085166 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Freese, H.M.; Ringel, V.; Overmann, J.; Petersen, J. Beyond the ABCs—Discovery of Three New Plasmid Types in Rhodobacterales (RepQ, RepY, RepW). Microorganisms 2022, 10, 738. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10040738
AMA Style
Freese HM, Ringel V, Overmann J, Petersen J. Beyond the ABCs—Discovery of Three New Plasmid Types in Rhodobacterales (RepQ, RepY, RepW). Microorganisms. 2022; 10(4):738. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10040738
Chicago/Turabian StyleFreese, Heike M., Victoria Ringel, Jörg Overmann, and Jörn Petersen. 2022. "Beyond the ABCs—Discovery of Three New Plasmid Types in Rhodobacterales (RepQ, RepY, RepW)" Microorganisms 10, no. 4: 738. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10040738
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.