
Epidemiology of Non-O157 Shiga Toxin-Producing Escherichia coli in the Province of Alberta, Canada, from 2018 to 2021


Abstract
:1. Introduction
2. Materials and Methods
2.1. Submission and Testing of Clinical Samples
2.2. Data Collection
2.3. Inclusion and Exclusion Criteria
3. Results
3.1. Serogroups and Serotypes Identified
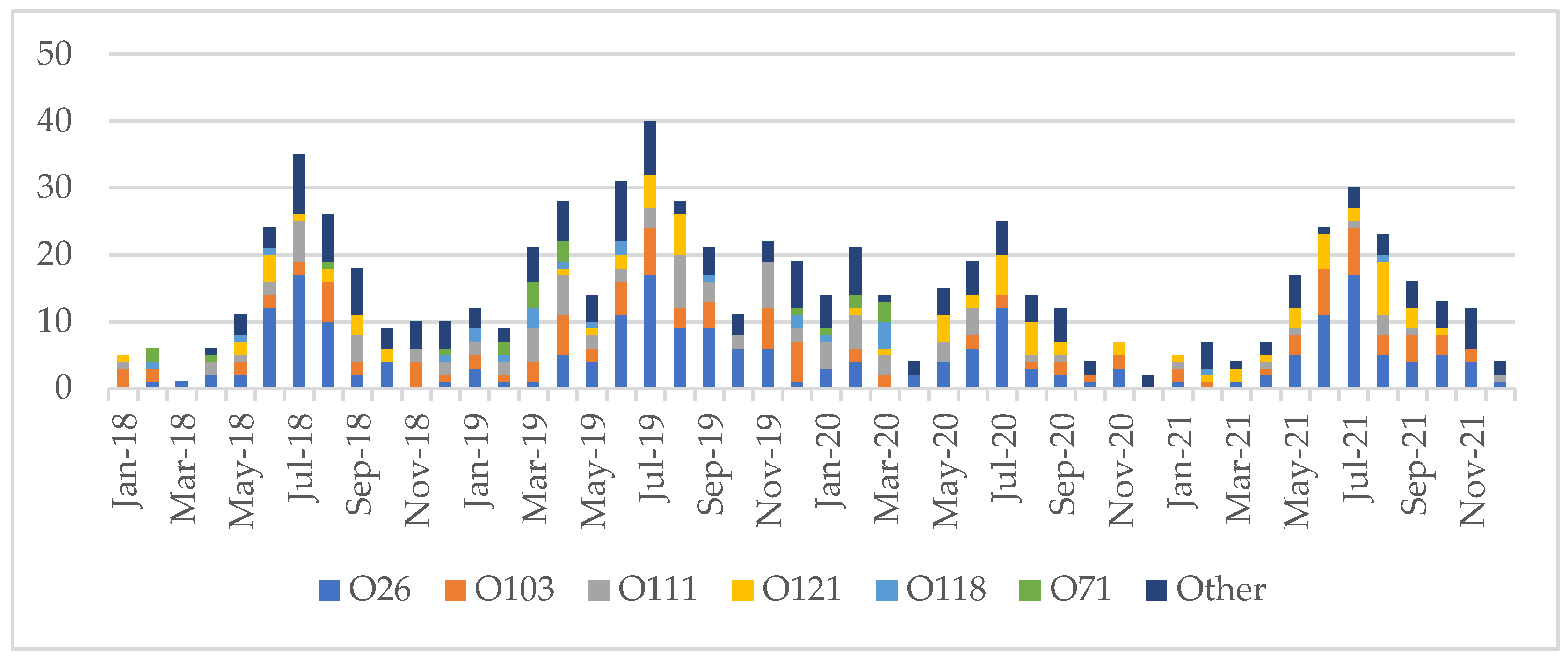
3.2. Seasonality
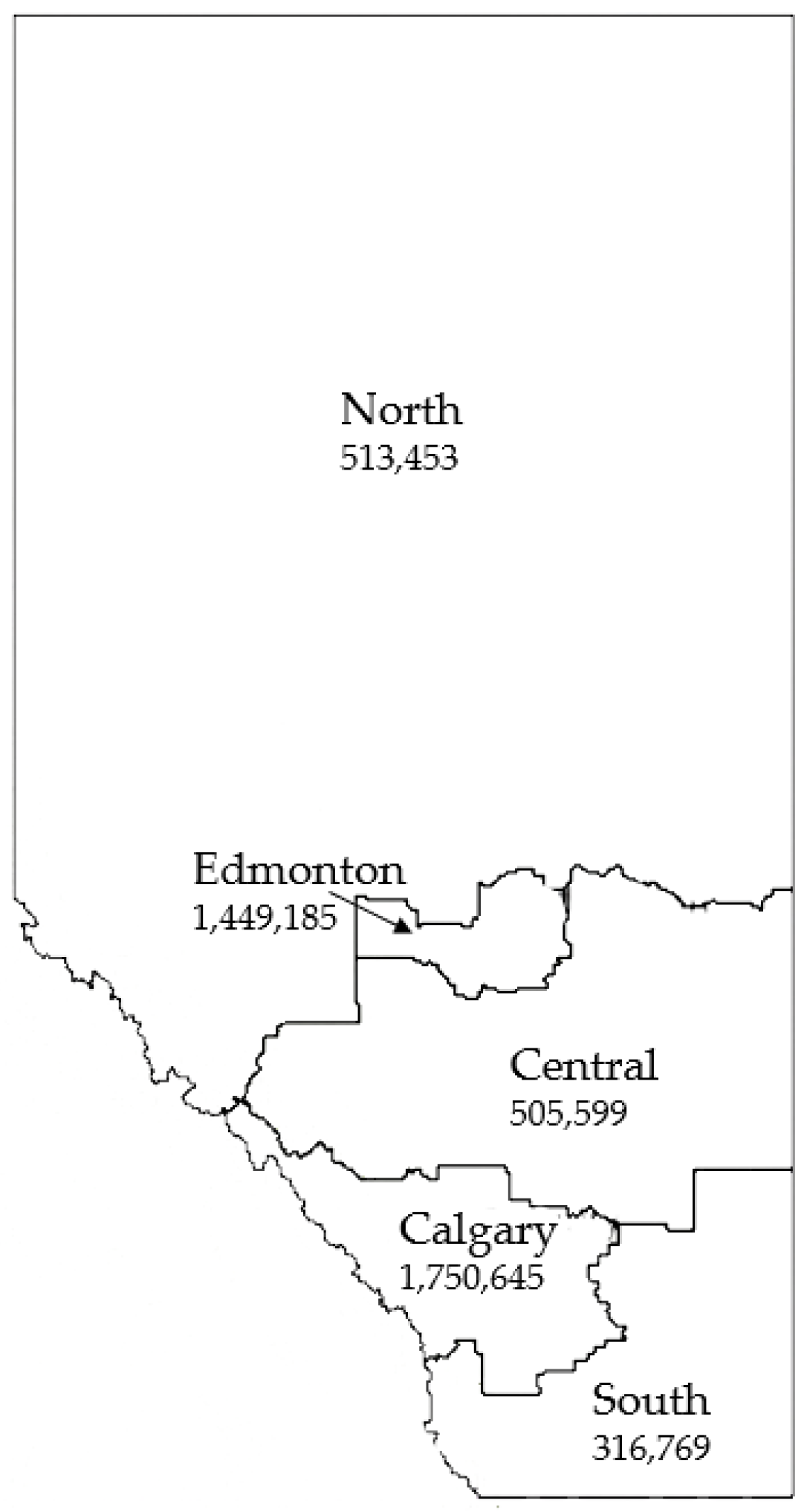
3.3. Geographic Distribution
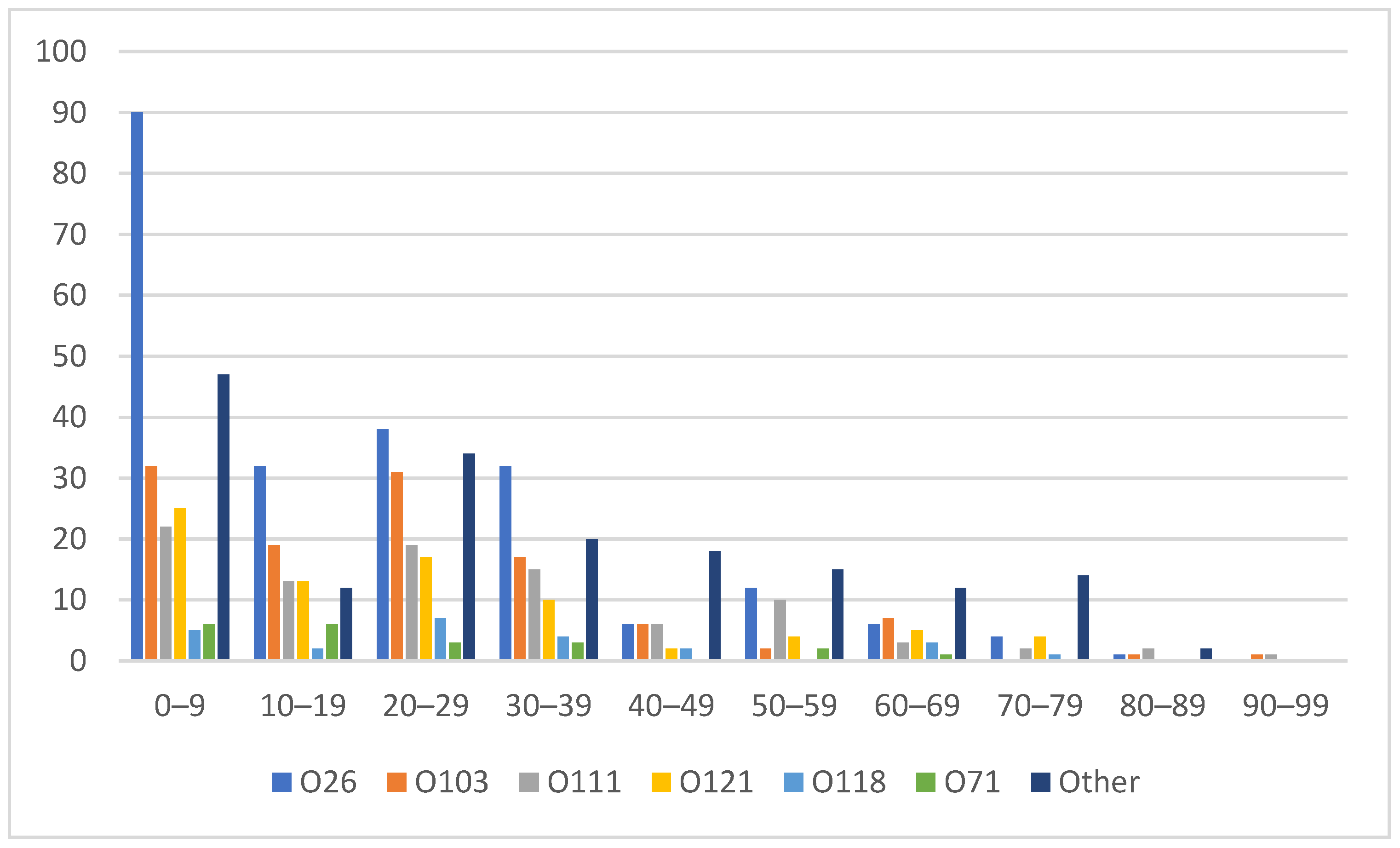
3.4. Age and Sex Distribution
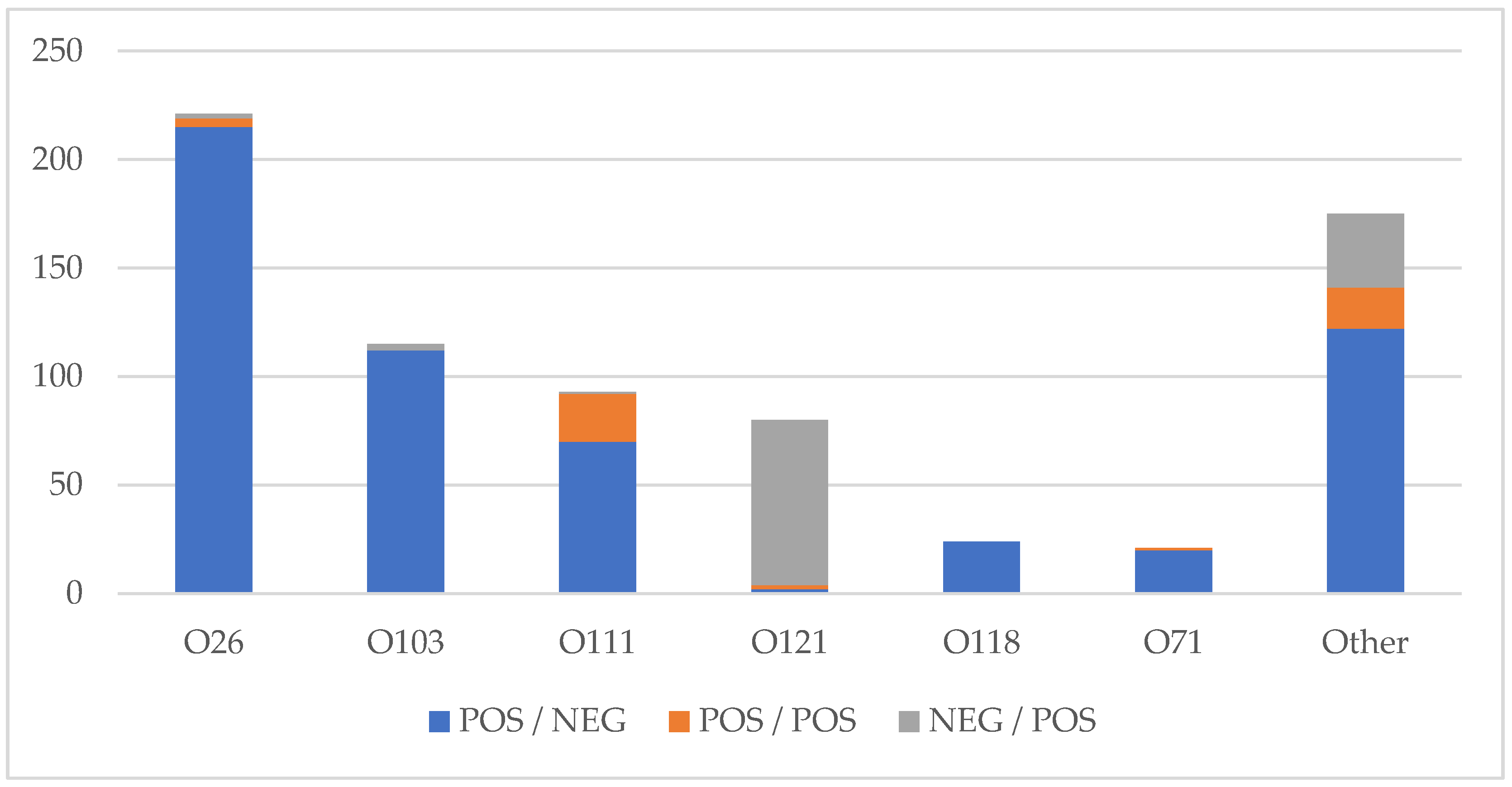
3.5. Shiga Toxin Profile
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Newell, D.G.; la Ragione, R.M. Enterohaemorrhagic and other Shiga toxin-producing Escherichia coli (STEC): Where are we now regarding diagnostics and control strategies? Transbound. Emerg. Dis. 2018, 65, 49–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vishram, B.; Jenkins, C.; Greig, D.R.; Godbole, G.; Carroll, K.; Balasegaram, S.; Byrne, L. The emerging importance of Shiga toxin-producing Escherichia coli other than serogroup O157 in England. J. Med. Microbiol. 2021, 70, 7. [Google Scholar] [CrossRef] [PubMed]
- Gould, L.H.; Mody, R.K.; Ong, K.L.; Clogher, P.; Cronquist, A.B.; Garman, K.N.; Lathrop, S.; Medus, C.; Spina, N.L.; Webb, T.H.; et al. Increased recognition of Non-O157 shiga toxin-producing Escherichia coli infections in the United States during 2000–2010: Epidemiologic features and comparison with E. coli O157 infections. Foodborne Pathog. Dis. 2013, 10, 453–460. [Google Scholar] [CrossRef]
- Bettelheim, K.A. The non-O157 Shiga-toxigenic (verocytotoxigenic) Escherichia coli; Under-Rated pathogens. Crit. Rev. Microbiol. 2007, 33, 67–87. [Google Scholar] [CrossRef] [PubMed]
- Chui, L.; Christianson, S.; Alexander, D.; Arseneau, V.; Bekal, S.; Berenger, B.; Chen, Y.; Davidson, R.; Farrell, D.J.; German, G.J.; et al. CPHLN recommendations for the laboratory detection of Shiga toxin-producing Escherichia coli (O157 and non-O157). Can. Commun. Dis. Rep. 2018, 44, 304–307. [Google Scholar] [CrossRef]
- Shane, A.L.; Mody, R.K.; Crump, J.A.; Tarr, P.I.; Steiner, T.S.; Kotloff, K.; Langley, J.M.; Wanke, C.; Warren, C.A.; Cheng, A.C.; et al. 2017 Infectious Diseases Society of America Clinical Practice Guidelines for the Diagnosis and Management of Infectious Diarrhea. Clin. Infect. Dis. 2017, 65, 1963–1973. [Google Scholar] [CrossRef]
- Parsons, B.D.; Zelyas, N.; Berenger, B.M.; Chui, L. Detection, characterization, and typing of shiga toxin-producing Escherichia coli. Front. Microbiol. 2016, 7, 478. [Google Scholar] [CrossRef] [Green Version]
- Bryan, A.; Youngster, I.; McAdam, A.J. Shiga Toxin Producing Escherichia coli. Clin. Lab. Med. 2015, 35, 247–272. [Google Scholar] [CrossRef]
- Strockbine, N.A.; Marques, L.R.M.; Newland, J.W.; Holmes, R.K.; O’Brien, A.D.; Smith, H.W. Two toxin-converting phages from Escherichia coli O157:H7 strain 933 encode antigenically distinct toxins with similar biologic activities. Infect. Immun. 1986, 53, 135–140. [Google Scholar] [CrossRef] [Green Version]
- Amézquita-López, B.A.; Soto-Beltrán, M.; Lee, B.G.; Yambao, J.C.; Quiñones, B. Isolation, genotyping and antimicrobial resistance of Shiga toxin-producing Escherichia coli. J. Microbiol. Immunol. Infect. 2018, 51, 425–434. [Google Scholar] [CrossRef]
- Fuller, C.A.; Pellino, C.A.; Weiss, A.A.; Flagler, M.J.; Strasser, J.E. Shiga toxin subtypes display dramatic differences in potency. Infect. Immun. 2011, 79, 1329–1337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valilis, E.; Ramsey, A.; Sidiq, S.; DuPont, H.L. Non-O157 Shiga toxin-producing Escherichia coli—A poorly appreciated enteric pathogen: Systematic review. Int. J. Infect. Dis. 2018, 76, 82–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Government of Alberta Notifiable Disease Report (NDR) Manual, 9th Edition. Available online: https://open.alberta.ca/publications/ndr-manual-9th-edition (accessed on 26 February 2022).
- Perelle, S.; Dilasser, F.; Grout, J.; Fach, P. Detection by 5′-nuclease PCR of Shiga-toxin producing Escherichia coli O26, O55, O91, O103, O111, O113, O145 and O157:H7, associated with the world’s most frequent clinical cases. Mol. Cell. Probes 2004, 18, 185–192. [Google Scholar] [CrossRef] [PubMed]
- GeoDiscover Alberta. Available online: https://geodiscover.alberta.ca/geoportal/#searchPanel (accessed on 26 February 2022).
- Alberta Interactive Health Data Application. Available online: http://www.ahw.gov.ab.ca/IHDA_Retrieval/ (accessed on 26 February 2022).
- European Centre for Disease Prevention and Control (ECDC). Shiga Toxin-Producing Escherichia coli (STEC) Infection. In ECDC. Annual Epidemiologic Report for 2019; ECDC: Stockholm, Sweden, 2021. [Google Scholar]
- Nüesch-Inderbinen, M.; Morach, M.; Cernela, N.; Althaus, D.; Jost, M.; Mäusezahl, M.; Bloomberga, G.; Stephan, R. Serotypes and virulence profiles of Shiga toxin-producing Escherichia coli strains isolated during 2017 from human infections in Switzerland. Int. J. Med. Microbiol. 2018, 308, 933–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, K.H.; Jung, S.M.; Shin, E.; Chung, G.T.; Seong, W.K.; Cho, S.H. Comparison of enterohemorrhagic Escherichia coli (EHEC) O157 and EHEC non-O157 isolates from patients with diarrhea in Korea. Jpn. J. Infect. Dis. 2017, 70, 320–322. [Google Scholar] [CrossRef] [Green Version]
- Kameyama, M.; Yabata, J.; Nomura, Y.; Tominaga, K. Biochemical features and virulence gene profiles of non-O157/O26 enterohemorrhagic Escherichia coli strains from humans in the Yamaguchi Prefecture, Japan. Jpn. J. Infect. Dis. 2015, 68, 216–220. [Google Scholar] [CrossRef] [Green Version]
- Noftall, K.; Taylor, M.; Hoang, L.; Galanis, E. Shiga toxin–producing Escherichia coli in British Columbia, 2011–2017: Analysis to inform exclusion guidelines. Can. Commun. Dis. Rep. 2019, 45, 238–243. [Google Scholar] [CrossRef]
- Luna-Gierke, R.E.; Griffin, P.M.; Gould, L.H.; Herman, K.; Bopp, C.A.; Strockbine, N.; Mody, R.K. Outbreaks of non-O157 Shiga toxin-producing Escherichia coli infection: USA. Epidemiol. Infect. 2014, 142, 2270–2280. [Google Scholar] [CrossRef]
- Butt, S.; Jenkins, C.; Godbole, G.; Byrne, L. The epidemiology of Shiga toxin-producing Escherichia coli serogroup O157 in England, 2009–2019. Epidemiol. Infect. 2022, 150, e52. [Google Scholar] [CrossRef]
- Marder, E.P.; Griffin, P.M.; Cieslak, P.R.; Dunn, J.; Hurd, S.; Jervis, R.; Lathrop, S.; Muse, A.; Ryan, P.; Smith, K.; et al. Preliminary incidence and trends of infections with pathogens transmitted commonly through food—Foodborne diseases active surveillance network, 10 U.S. Sites, 2006–2017. Morb. Mortal. Wkly. Report. 2017, 67, 324–328. [Google Scholar] [CrossRef]
- Lisboa, L.F.; Szelewicki, J.; Lin, A.; Latonas, S.; Li, V.; Zhi, S.; Parsons, B.D.; Berenger, B.; Fathima, S.; Chui, L. Epidemiology of shiga toxin-producing Escherichia coli O157 in the Province of Alberta, Canada, 2009–2016. Toxins 2019, 11, 613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wylie, J.L.; van Caeseele, P.; Gilmour, M.W.; Sitter, D.; Guttek, C.; Giercke, S. Evaluation of a New Chromogenic Agar Medium for Detection of Shiga Toxin-Producing Escherichia coli (STEC) and Relative Prevalences of O157 and Non-O157 STEC in Manitoba, Canada. J. Clin. Microbiol. 2013, 51, 466–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gill, A.; Gill, C.O. Non-O157 verotoxigenic Escherichia coli and beef: A Canadian perspective. Can. J. Vet. Res. 2010, 74, 161–169. [Google Scholar] [PubMed]
- Stromberg, Z.R.; Redweik, G.A.J.; Mellata, M. Detection, Prevalence, and Pathogenicity of Non-O157 Shiga Toxin-Producing Escherichia coli from Cattle Hides and Carcasses. Foodborne Pathog. Dis. 2018, 15, 119–131. [Google Scholar] [CrossRef]
- Tack, D.M.; Kisselburgh, H.M.; Richardson, L.C.; Geissler, A.; Griffin, P.M.; Payne, D.C.; Gleason, B.L. Shiga toxin-producing Escherichia coli outbreaks in the United States, 2010–2017. Microorganisms 2021, 9, 1529. [Google Scholar] [CrossRef]
- Kintz, E.; Brainard, J.; Hooper, L.; Hunter, P. Transmission pathways for sporadic Shiga-toxin producing E. coli infections: A systematic review and meta-analysis. Int. J. Hyg. Environ. Health 2017, 220, 57–67. [Google Scholar] [CrossRef] [Green Version]
- Frank, C.; Werber, D.; Cramer, J.P.; Askar, M.; Faber, M.; an der Heiden, M.; Bernard, H.; Fruth, A.; Prager, R.; Spode, A.; et al. Epidemic Profile of Shiga-Toxin–Producing Escherichia coli O104:H4 Outbreak in Germany. N. Engl. J. Med. 2011, 365, 1771–1780. [Google Scholar] [CrossRef] [Green Version]
- Outbreak of E. coli Infections Linked to Ground Beef: Final Update. Available online: https://www.cdc.gov/ecoli/2019/o103-04-19/index.html (accessed on 26 February 2022).
- Kanayama, A.; Yahata, Y.; Arima, Y.; Takahashi, T.; Saitoh, T.; Kanou, K.; Kawabata, K.; Sunagawa, T.; Matsui, T.; Oishi, K. Enterohemorrhagic Escherichia coli outbreaks related to childcare facilities in Japan, 2010–2013. BMC Infect. Dis. 2015, 15, 539. [Google Scholar] [CrossRef]
- Karmali, M.A. Factors in the emergence of serious human infections associated with highly pathogenic strains of shiga toxin-producing Escherichia coli. Int. J. Med. Microbiol. 2018, 308, 1067–1072. [Google Scholar] [CrossRef]
- Harries, M.; Dreesman, J.; Rettenbacher-Riefler, S.; Mertens, E. Faecal carriage of extended-spectrum β-lactamase-producing Enterobacteriaceae and Shiga toxin-producing Escherichia coli in asymptomatic nursery children in Lower Saxony (Germany), 2014. Epidemiol. Infect. 2016, 144, 3540–3548. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reference Gene, Primer/Probe | Sequence 5′-3′ |
|---|---|
| stx1, stx2-F | TTT GTY ACT GTS ACA GCW GAA GCY TTA CG |
| stx1, stx2-R | CCC CAG TTC ARW GTR AGR TCM ACR TC |
| stx1-P | CTG GAT GAT CTC AGT GGG CGT TCT TAT GTA A |
| stx2-P | TCG TCA GGC ACT GTC TGA AAC TGC TCC |
| Serogroup | Number of Isolates (%) | Serotype | Number of Isolates (% of Serogroup) |
|---|---|---|---|
| O26 | 221 (30.3) | H11 | 155 (70.1) |
| H Non-motile | 65 (29.4) | ||
| H Undetermined | 1 (0.5) | ||
| O103 | 116 (15.9) | H2 | 80 (69.0) |
| H25 | 20 (17.2) | ||
| H11 | 8 (6.9) | ||
| H Non-motile | 6 (5.2) | ||
| H19 | 2 (1.7) | ||
| O111 | 93 (12.8) | H Non-motile | 93 (100) |
| O121 | 80 (11.0) | H19 | 77 (96.3) |
| H Non-motile | 3 (3.7) | ||
| O118 | 24 (3.3) | H16 | 10 (41.6) |
| H2 | 9 (37.5) | ||
| H Undetermined | 3 (12.5) | ||
| H14 | 1 (4.2) | ||
| H Non-motile | 1 (4.2) | ||
| O71 | 21 (2.9) | H11 | 16 (76.2) |
| H8 | 3 (14.3) | ||
| H Non-motile | 2 (9.5) | ||
| Other | 167 (22.9) | - | - |
| Unknown * | 7 (0.9) | - | - |
| Total | 729 (100) | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Glassman, H.; Ferrato, C.; Chui, L. Epidemiology of Non-O157 Shiga Toxin-Producing Escherichia coli in the Province of Alberta, Canada, from 2018 to 2021. Microorganisms 2022, 10, 814. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10040814
Glassman H, Ferrato C, Chui L. Epidemiology of Non-O157 Shiga Toxin-Producing Escherichia coli in the Province of Alberta, Canada, from 2018 to 2021. Microorganisms. 2022; 10(4):814. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10040814
Chicago/Turabian StyleGlassman, Heather, Christina Ferrato, and Linda Chui. 2022. "Epidemiology of Non-O157 Shiga Toxin-Producing Escherichia coli in the Province of Alberta, Canada, from 2018 to 2021" Microorganisms 10, no. 4: 814. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10040814