
Changes in Lake Sturgeon Gut Microbiomes Relative to Founding Origin and in Response to Chemotherapeutant Treatments


Abstract
:1. Introduction
2. Methods
2.1. Study Site
2.2. Study Fish
2.2.1. Hatchery-Produced Gamete Collection, Fertilization and Incubation
2.2.2. Field Collection and Incubation of Wild Harvested Eggs and Larval Production
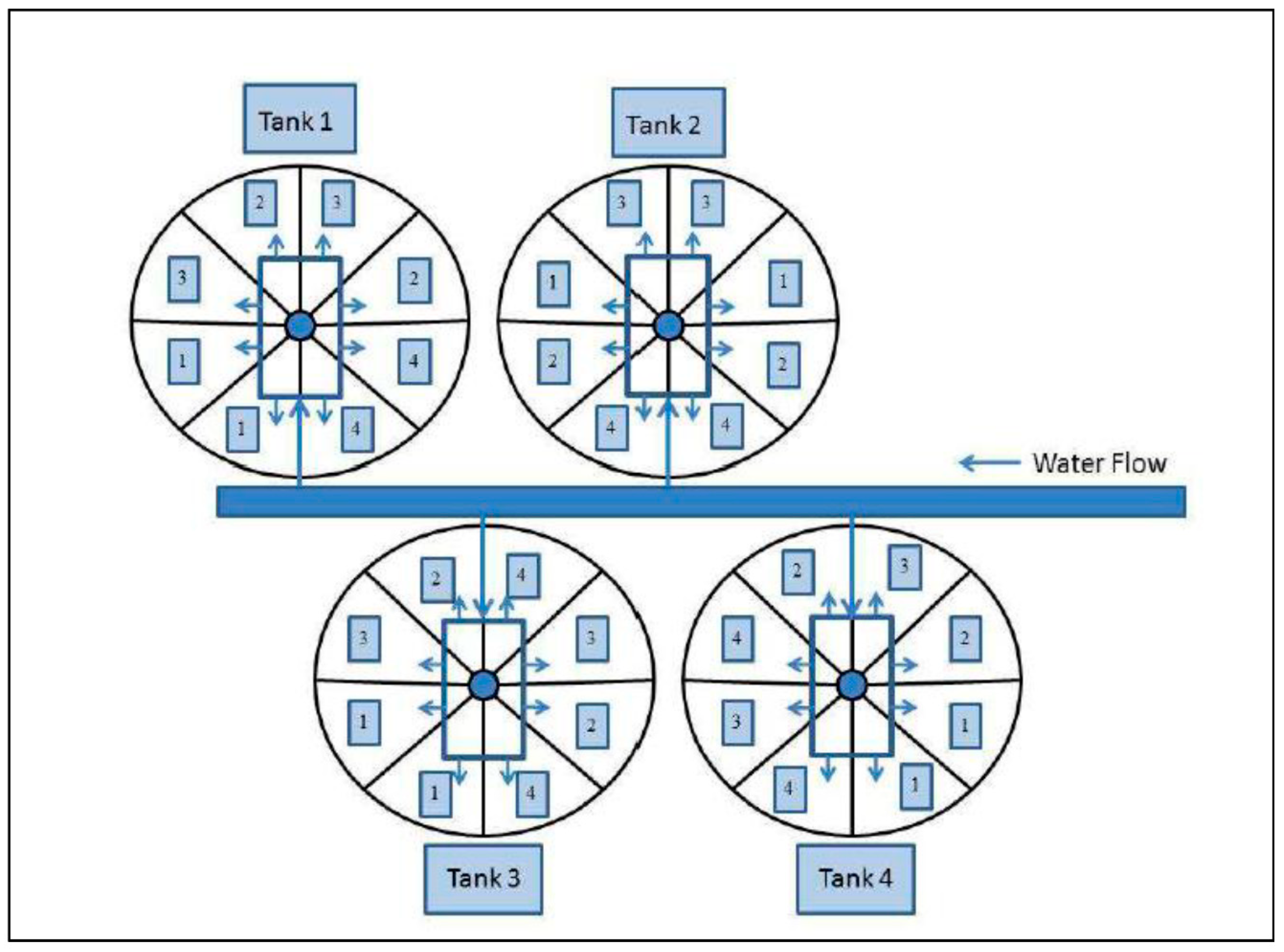
2.3. Experimental Treatments
2.4. Fish Dissection, DNA Isolation, PCR Validation
2.5. 16S rRNA Amplicon Sequencing and Sequence Pipeline Analyses
2.6. Statistical Analysis of Bacterial Community Profiles and Ecological Statistical Analyses
2.6.1. Alpha Diversity
2.6.2. Beta Diversity
2.6.3. Differential Abundance of OTUs and Biomarker Identification across Treatments
3. Results
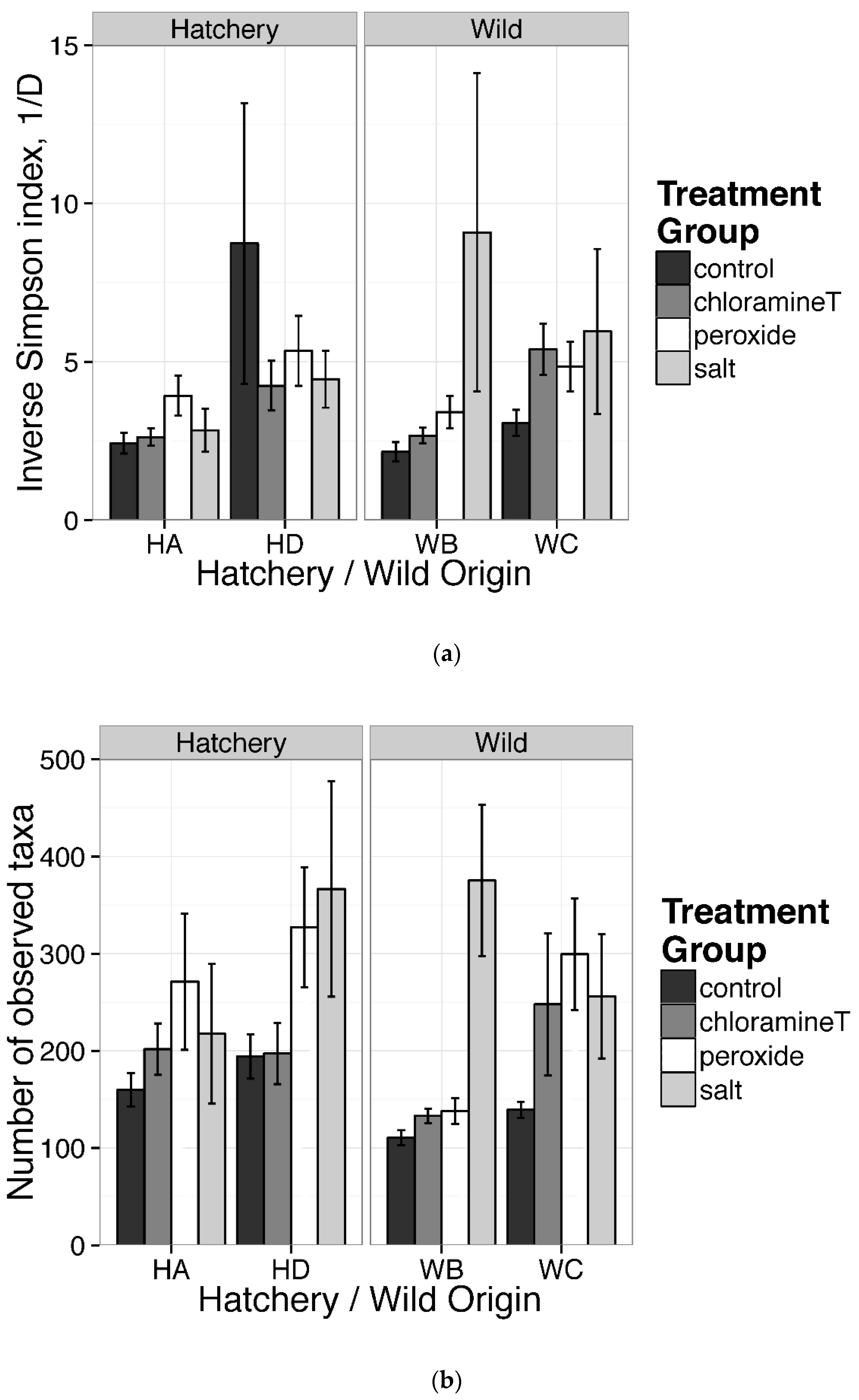
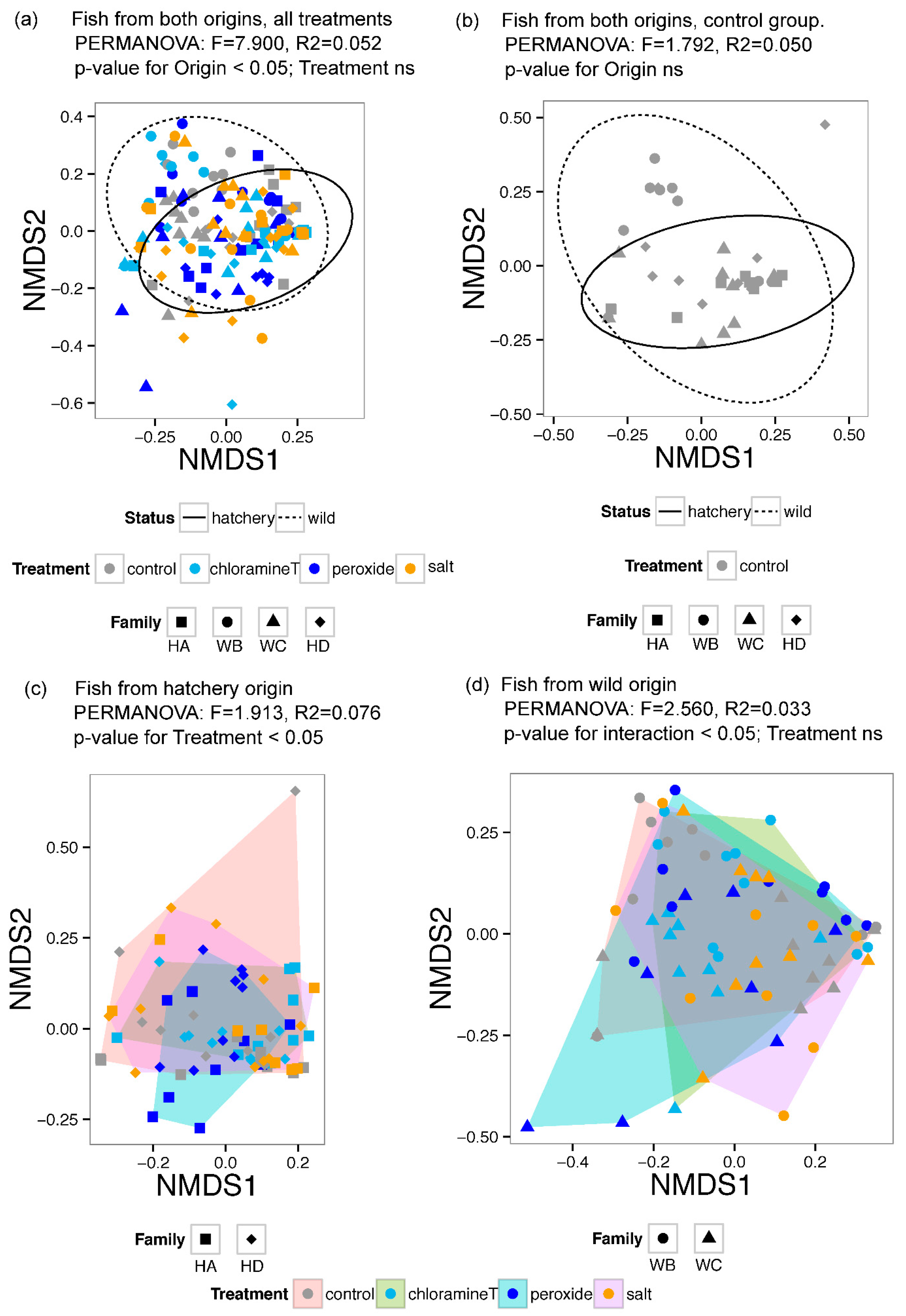
3.1. Diversity of Gut Microbial Community Composition
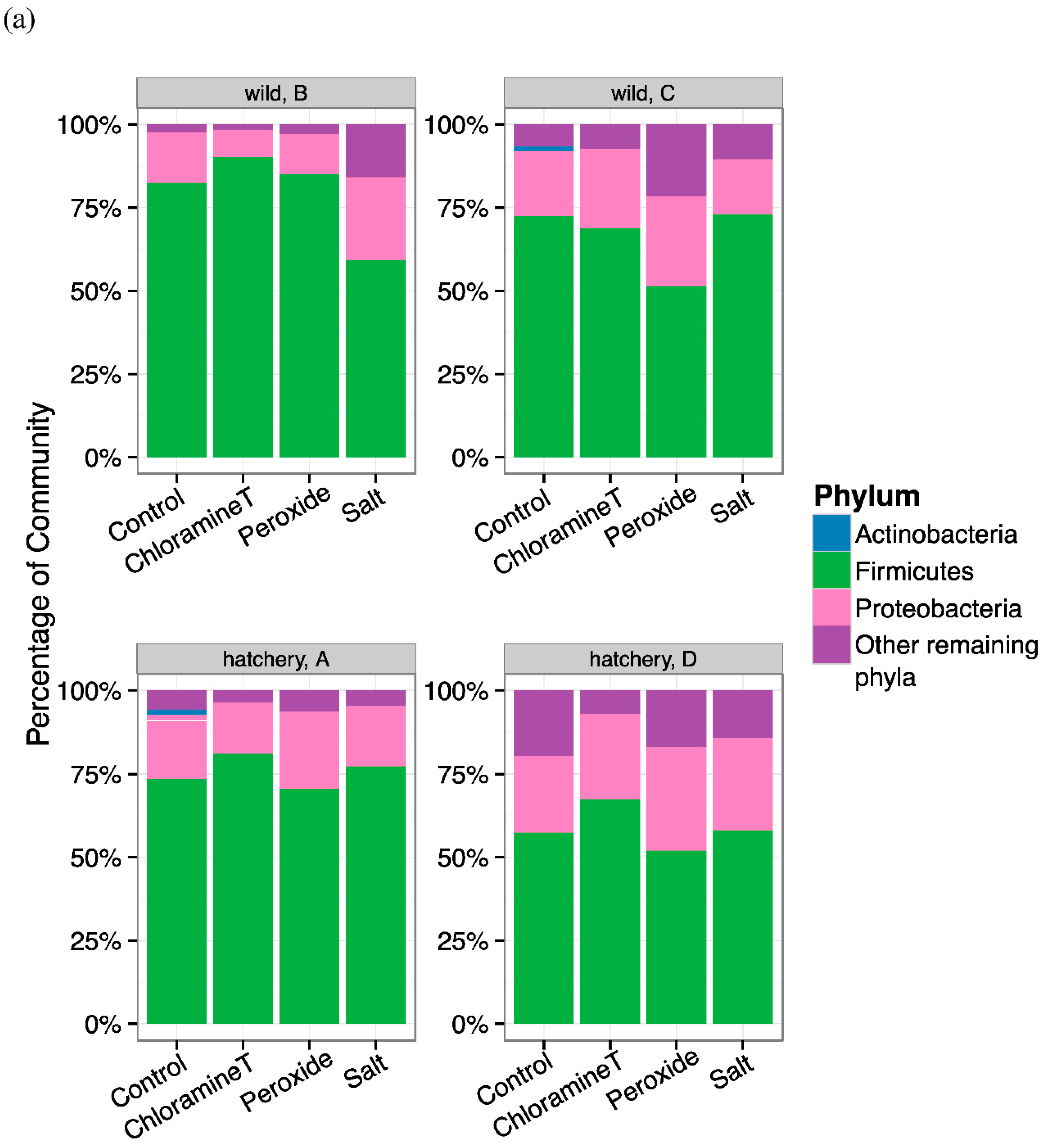
3.2. Differences in Gut Microbial Community Composition between Fish Group Origin and among Chemotherapeutant Treatments
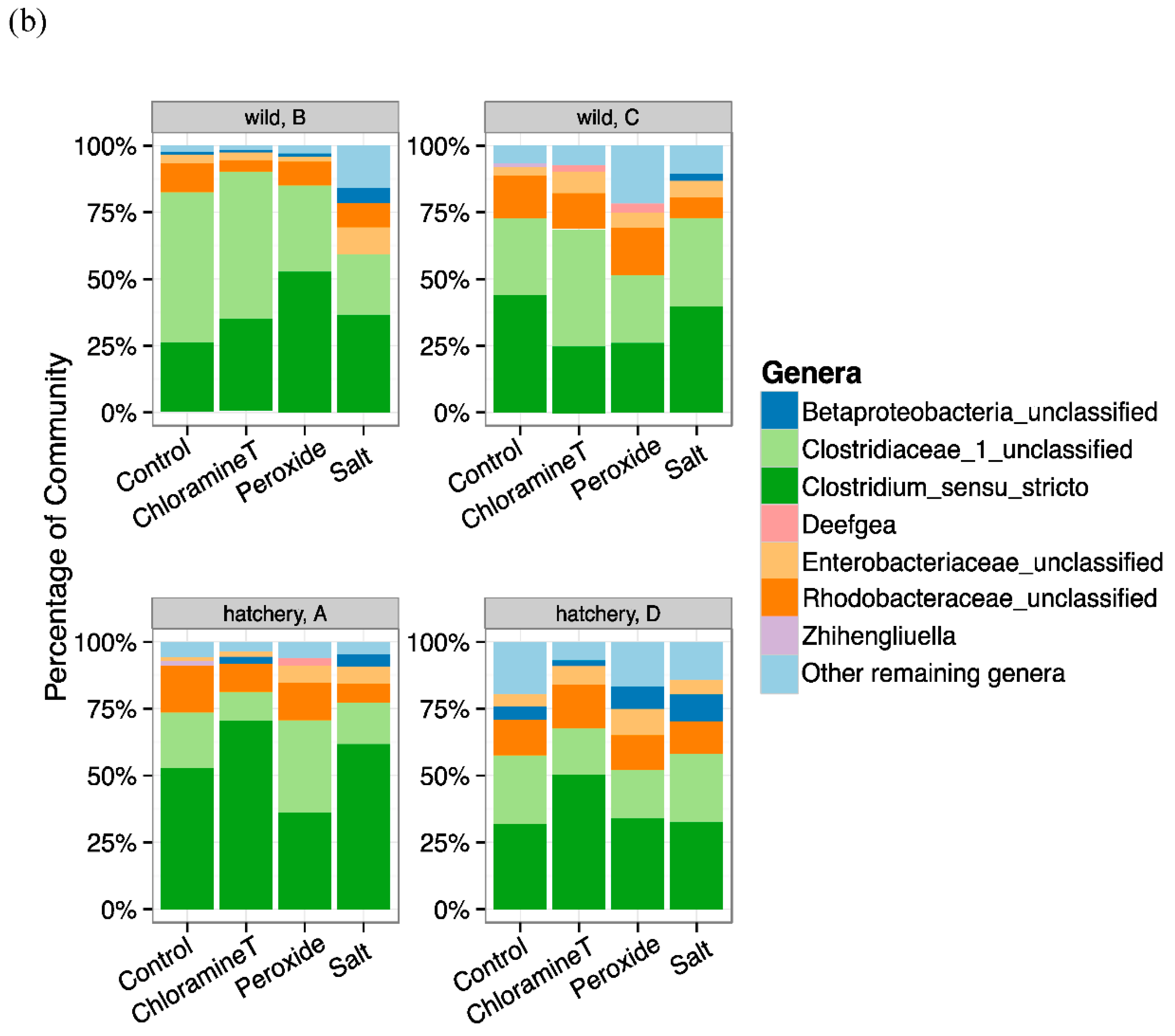
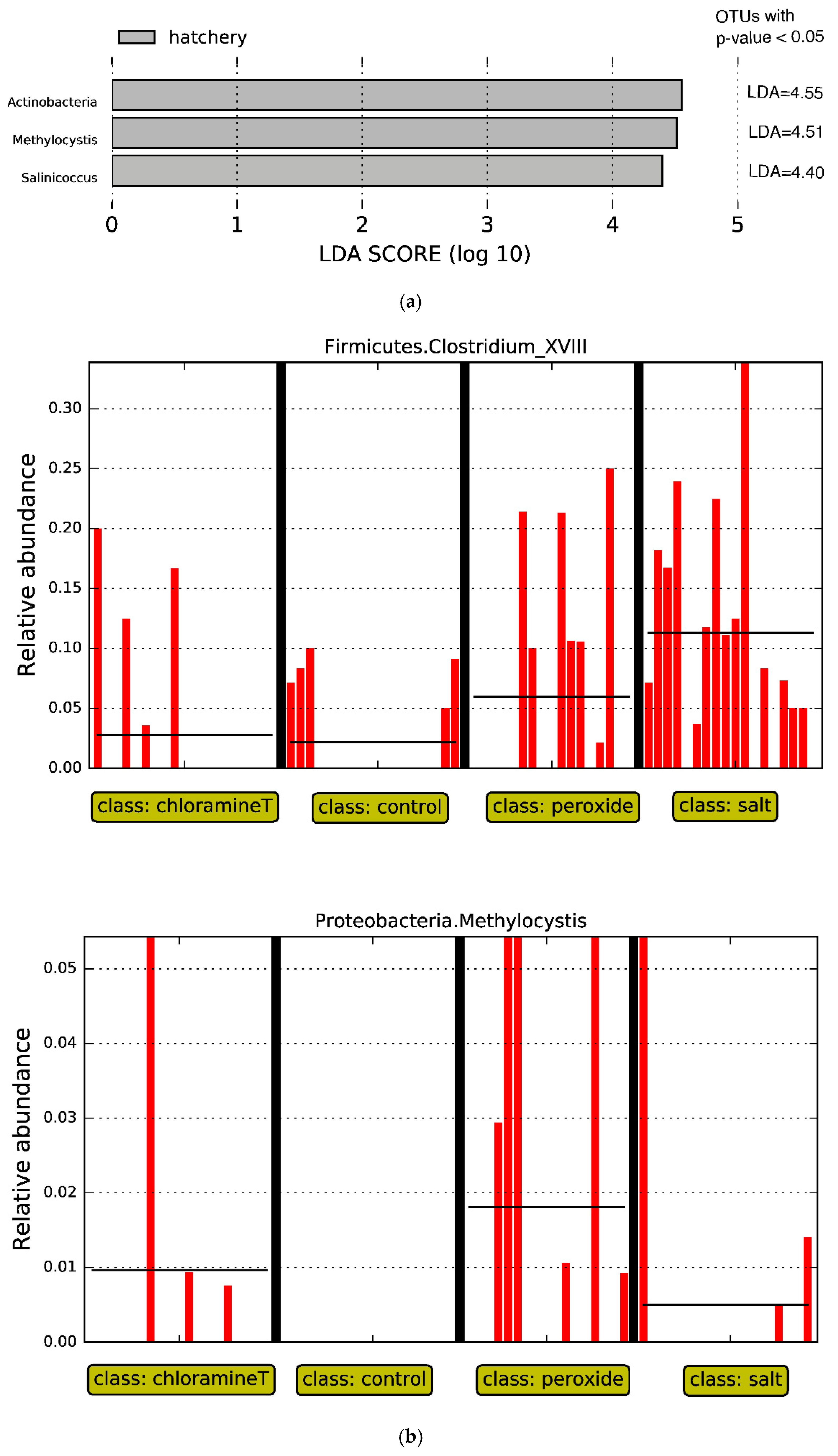
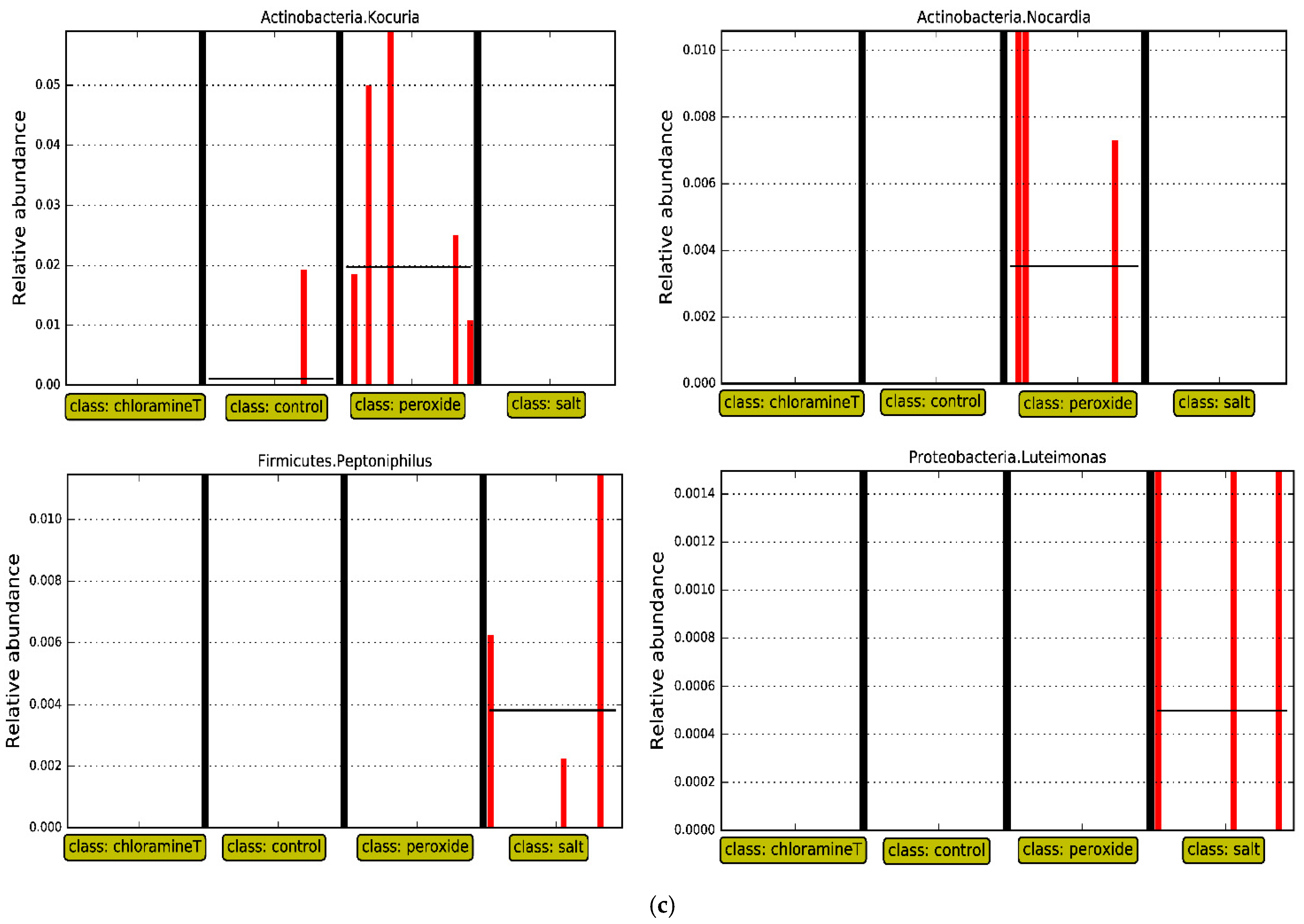
3.3. Identification of Bacterial Taxa Influenced by Chemotherapeutant Treatments
4. Discussion
4.1. General Findings and Relevance to Aquaculture
4.2. Effects of Chemotherapeutant Treatments on Larval Gut Microbial Communities
4.3. Sources of Heterogeneity Associated with Microbial Community Origin
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Post, G. Textbook of Fish Health; T.F.H. Publications: Austin, TX, USA, 1987. [Google Scholar]
- Subasinghe, R.P.; Phillips, M.J. Aquatic animal health management: Opportunities and challenges for rural, small-scale aquaculture and enhanced-fisheries development: Workshop introductory remarks. In Primary Aquatic Animal Health Care in Rural, Small-Scale, Aquaculture Development; Arthur, J.R., Phillips, M.J., Eds.; FAO: Rome, Italy, 2002; pp. 1–15. [Google Scholar]
- Conte, F.S. Stress and the welfare of cultured fish. Appl. Anim. Behav. Sci. 2004, 86, 205–223. [Google Scholar] [CrossRef]
- Davis, M.W.; Ottmar, M.L. Wounding and reflex impairment may be predictors for mortality in discarded or escaped fish. Fish. Res. 2006, 82, 1–6. [Google Scholar] [CrossRef]
- Llewellyn, M.S.; Boutin, S.; Hoseinifar, S.H.; Derome, N. Teleost microbiomes: The state of the art in their characterization, manipulation and importance in aquaculture and fisheries. Front. Microbiol. 2014, 5, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dethlefsen, L.; Huse, S.; Sogin, M.L.; Relman, D.A. The pervasive effects of an antibiotic on the human gut microbiota, as revealed by deep 16S rRNA sequencing. PLoS Biol. 2008, 6, e280. [Google Scholar] [CrossRef]
- Dethlefsen, L.; Relman, D.A. Incomplete recovery and individualized responses of the human distal gut microbiota to repeated antibiotic perturbation. Proc. Natl. Acad. Sci. USA 2011, 108, 4554–4561. [Google Scholar] [CrossRef] [Green Version]
- Bowker, J.; Trushenski, J.; Straus, D.; Gaikowski, M.; Goodwin, A. Guide to Using Drugs, Biologics, and Other Chemicals in Aquaculture; American Fisheries Society Fish Culture Section: Bethesda, MD, USA, 2016. [Google Scholar]
- Thorburn, M.A.; Moccia, R.D. Use of Chemotherapeuties on Trout Farms in Ontario. J. Aquat. Anim. Health 1993, 5, 85–91. [Google Scholar] [CrossRef]
- Gaikowski, M.P.; Larson, W.J.; Gingerich, W.H. Survival of cool and warm freshwater fish following chloramine-T exposure. Aquaculture 2008, 275, 20–25. [Google Scholar] [CrossRef]
- Tort, L.; Balasch, J.C.; Mackenzie, S. Fish immune system. A crossroads between innate and adaptive responses. Immunologia 2003, 22, 277–286. [Google Scholar]
- Speare, D.J.; Arsenault, G.J. Effects of intermittent hydrogen peroxide exposure on growth and columnaris disease prevention of juvenile rainbow trout (Oncorhynchus mykiss). Can. J. Fish. Aquat. Sci. 1997, 54, 2653–2658. [Google Scholar] [CrossRef]
- Rach, J.J.; Gaikowski, M.P.; Ramsay, R.T. Efficacy of Hydrogen Peroxide to Control Mortalities Associated with Bacterial Gill Disease Infections on Hatchery-Reared Salmonids. J. Aquat. Anim. Health 2000, 12, 119–127. [Google Scholar] [CrossRef]
- Rach, J.J.; Gaikowski, M.P.; Ramsay, R.T. Efficacy of Hydrogen Peroxide to Control Parasitic Infestations on Hatchery-Reared Fish. J. Aquat. Anim. Health 2000, 12, 267–273. [Google Scholar] [CrossRef]
- Rach, J.J.; Schleis, S.M.; Gaikowski, M.; Johnson, A. Efficacy of Hydrogen Peroxide in Controlling Mortality Associated with External Columnaris on Walleye and Channel Catfish Fingerlings. N. Am. J. Aquac. 2003, 65, 300–305. [Google Scholar] [CrossRef]
- Schelkle, B.; Doetjes, R.; Cable, J. The salt myth revealed: Treatment of gyrodactylid infections on ornamental guppies. Poecilia reticulata. Aquaculture 2011, 311, 74–79. [Google Scholar] [CrossRef]
- Noga, E.J. Fish Disease: Diagnosis and Treatment; Wiley-Blackwell: Ames, IA, USA, 2011. [Google Scholar]
- Swann, L.; Fitzgerald, S. Use and Application of Salt in Aquaculture. North Central Regional Aquaculture Center. 1992. Available online: http://agrilife.org/fisheries2/files/2013/09/The-Use-and-Application-of-Salt-in-Aquaculture.pdf (accessed on 10 January 2022).
- Durborow, R.M.; Francis-floyd, R. Medicated Feed for Food Fish. Sounthern Regional Aquaculture Center. 1996. Available online: https://www.ncrac.org/files/biblio/SRAC0473.pdf (accessed on 10 January 2022).
- Piper, R.G.; McElwain, I.B.; Orme, L.E.; McCraren, J.P.; Fowler, L.G.; Leonard, R.J. Fish Hatchery Management; U.S. Fish and Wildlife Service: Washington, DC, USA, 1982.
- Sanchez, J.G.; Speare, D.J.; Macnair, N.; Johnson, G. Effects of a Prophylactic Chloramine-T Treatment on Growth Performance and Condition Indices of Rainbow Trout. J. Aquat. Anim. Health 1996, 8, 278–284. [Google Scholar] [CrossRef]
- Gaikowski, M.P.; Rach, J.J.; Ramsay, R.T. Acute toxicity of hydrogen peroxide treatments to selected lifestages of cold-, cool-, and warmwater fish. Aquaculture 1999, 178, 191–207. [Google Scholar] [CrossRef]
- Magondu, E.W.; Rasowo, J.; Oyoo-Okoth, E.; Charo-Karisa, H. Evaluation of sodium chloride (NaCl) for potential prophylactic treatment and its short-term toxicity to African catfish Clarias gariepinus (Burchell 1822) yolk-sac and swim-up fry. Aquaculture 2011, 319, 307–310. [Google Scholar] [CrossRef]
- Vellend, M. Conceptual synthesis in community ecology. Q Rev. Biol. 2010, 85, 183–206. [Google Scholar] [CrossRef] [Green Version]
- Sloan, W.T.; Lunn, M.; Woodcock, S.; Head, I.M.; Nee, S.; Curtis, T.P. Quantifying the roles of immigration and chance in shaping prokaryote community structure. Environ. Microbiol. 2006, 8, 732–740. [Google Scholar] [CrossRef]
- Burns, A.R.; Stephens, W.Z.; Stagaman, K.; Wong, S.; Rawls, J.F.; Guillemin, K.; Bohannan, B.J.M. Contribution of neutral processes to the assembly of gut microbial communities in the zebrafish over host development. ISME J. 2016, 10, 655–664. [Google Scholar] [CrossRef] [Green Version]
- Venkataraman, A.; Bassis, C.M.; Beck, J.M.; Young, V.B.; Curtis, J.L.; Huffnagle, G.B.; Schmidt, T.M. Application of a Neutral Community Model to Assess Structuring of the Human Lung Microbiome. mBio 2015, 6, e02284-14. [Google Scholar] [CrossRef] [Green Version]
- Leibold, M.A.; Holyoak, M.; Mouquet, N.; Amarasekare, P.; Chase, J.M.; Hoopes, M.F.; Holt, R.D.; Shurin, J.B.; Law, R.; Tilman, D.; et al. The metacommunity concept: A framework for multi-scale community ecology. Ecol. Lett. 2004, 7, 601–613. [Google Scholar] [CrossRef]
- Nemergut, D.R.; Schmidt, S.K.; Fukami, T.; O’Neill, S.P.; Bilinski, T.M.; Stanish, L.F.; Knelman, J.E.; Darcy, J.L.; Lynch, R.C.; Wickey, P.; et al. Patterns and processes of microbial community assembly. Microbiol. Mol. Biol. Rev. 2013, 77, 342–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christian, N.; Whitaker, B.K.; Clay, K. Microbiomes: Unifying animal and plant systems through the lens of community ecology theory. Front. Microbiol. 2015, 6, 869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Louca, S.; Polz, M.F.; Mazel, F.; Albright, M.B.N.; Huber, J.A.; O’Connor, M.I.; Ackermann, M.; Hahn, A.S.; Srivastava, D.S.; Crowe, S.A.; et al. Function and functional redundancy in microbial systems. Nat. Ecol. Evol. 2018, 2, 936–943. [Google Scholar] [CrossRef]
- Cho, I.; Blaser, M.J. The human microbiome: At the interface of health and disease. Nat. Rev. Genet. 2012, 13, 260–270. [Google Scholar] [CrossRef] [Green Version]
- Francino, M.P. Antibiotics and the human gut microbiome: Dysbioses and accumulation of resistances. Front. Microbiol. 2016, 6, 1543. [Google Scholar] [CrossRef] [Green Version]
- Langdon, A.; Crook, N.; Dantas, G. The effects of antibiotics on the microbiome throughout development and alternative approaches for therapeutic modulation. Genome Med. 2016, 8, 39. [Google Scholar] [CrossRef] [Green Version]
- Verschuere, L.; Rombaut, G.; Sorgeloos, P.; Verstraete, W. Probiotic Bacteria as Biological Control Agents in Aquaculture. Microbiol. Mol. Biol. Rev. 2000, 64, 655–671. [Google Scholar] [CrossRef] [Green Version]
- Serrano, P.H. Responsible Use of Antibiotics in Aquaculture; Technical Paper; Food & Agriculture Org.: Rome, Italy, 2005. [Google Scholar]
- Dfoirdt, T.; Sorgeloos, P.; Bossier, P. Alternatives to antibiotics for the control of bacterial disease in aquaculture. Curr. Opin. Microbiol. 2011, 14, 251–258. [Google Scholar] [CrossRef]
- Diana, J.S. Aquaculture Production and Biodiversity Conservation. Bioscience 2009, 59, 27–38. [Google Scholar] [CrossRef]
- Cabello, F.C. Heavy use of prophylactic antibiotics in aquaculture: A growing problem for human and animal health and for the environment. Environ. Microbiol. 2006, 8, 1137–1144. [Google Scholar] [CrossRef] [PubMed]
- Mortazavi, A.L. Poppin’ the Prophylactics: An Analysis of Antibiotics in Aquaculture. Columbia Univ. J. Glob. Health 2014, 4, 23–27. [Google Scholar]
- Burka, J.F.; Hammell, K.L.; Horsberg, T.E.; Johnson, G.R.; Rainnie, D.J.; Speare, D.J. Drugs in salmonid aquaculture—A review. J. Vet. Pharmacol. Ther. 2003, 20, 333–349. [Google Scholar] [CrossRef] [PubMed]
- Romero, J.; Ringø, E.; Merrifield, D.L. The Gut Microbiota of Fish. In Aquaculture Nutrition: Gut Health, Probiotics and Prebiotics; Wiley: Chichester, UK, 2014. [Google Scholar]
- Austin, B.; Al-Zahrani, A.M.J. The effect of antimicrobial compounds on the gastrointestinal microflora of rainbow trout, Salmo gairdneri Richardson. J. Fish. Biol. 1988, 33, 1–14. [Google Scholar] [CrossRef]
- Navarrete, P.; Mardones, P.; Opazo, R.; Espejo, R.; Romero, J. Oxytetracycline treatment reduces bacterial diversity of intestinal microbiota of Atlantic salmon. J. Aquat. Anim. Health 2008, 20, 177–183. [Google Scholar] [CrossRef]
- He, S.; Zhou, Z.; Liu, Y.; Cao, Y.; Meng, K.; Shi, P.; Yao, B.; Ringø, E. Effects of the antibiotic growth promoters flavomycin and florfenicol on the autochthonous intestinal microbiota of hybrid tilapia (Oreochromis niloticus ♀ × O. aureus ♂). Arch. Microbiol. 2010, 192, 985–994. [Google Scholar] [CrossRef]
- Liu, Y.; Zhou, Z.; Wu, N.; Tao, Y.; Xu, L.; Cao, Y.; Zhang, Y.; Yao, B. Gibel carp Carassius auratus gut microbiota after oral administration of trimethoprim/sulfamethoxazole. Dis. Aquat. Organ. 2012, 99, 207–213. [Google Scholar] [CrossRef] [Green Version]
- Wang, A.R.; Ran, C.; Ringø, E.; Zhou, Z.G. Progress in fish gastrointestinal microbiota research. Rev Aquac. 2018, 10, 626–640. [Google Scholar] [CrossRef] [Green Version]
- Vadstein, O.; Bergh, Ø.; Gatesoupe, F.-J.; Galindo-Villegas, J.; Mulero, V.; Picchietti, S.; Scapigliati, G.; Makridis, P.; Olsen, Y.; Dierckens, K.; et al. Microbiology and immunology of fish larvae. Rev. Aquac. 2012, 5, S1–S25. [Google Scholar] [CrossRef] [Green Version]
- Holtgren, J.M.; Ogren, S.A.; Paquet, A.J.; Fajfer, S. Design of a portable streamside rearing facility for lake sturgeon. N. Amer. J. Aqua. 2007, 69, 317–323. [Google Scholar] [CrossRef]
- Flagg, T.A.; Nash, C.E. (Eds.) A Conceptual Framework for Conservation Hatchery Strategies for Pacific Salmonids; NOAA Technical Memorandum NMFS; NOAA: Washington, DC, USA, 1999.
- Fujimoto, M.; Crossman, J.A.; Scribner, K.T.; Marsh, T.L. Microbial Community Assembly and Succession on Lake Sturgeon Egg Surfaces as a Function of Simulated Spawning Stream Flow Rate. Microb. Ecol. 2013, 66, 500–511. [Google Scholar] [CrossRef] [PubMed]
- Abdul Razak, S.; Scribner, K.T. Ecological and Ontogenetic Components of Larval Lake Sturgeon Gut Microbiota Assembly, Successional Dynamics, and Ecological Evaluation of Neutral Community Processes. Appl. Environ. Microbiol. 2020, 86, e02662-19. [Google Scholar] [CrossRef] [PubMed]
- Abdul Razak, S.; Griffin, M.J.; Mischke, C.C.; Bosworth, B.G.; Waldbieser, G.C.; Wise, D.J.; Marsh, T.L.; Scribner, K.T. Biotic and abiotic factors influencing channel catfish egg and gut microbiome dynamics during early life stages. Aquaculture 2019, 498, 556–567. [Google Scholar] [CrossRef]
- Crossman, J.A.; Scribner, K.T.; Yen, D.T.; Davis, C.A.; Forsythe, P.S.; Baker, E.A. Gamete and larval collection methods and hatchery rearing environments affect levels of genetic diversity in early life stages of lake sturgeon (Acipenser fulvescens). Aquaculture 2011, 310, 312–324. [Google Scholar] [CrossRef]
- Hastings, R.P.; Bauman, J.M.; Baker, E.A.; Scribner, K.T. Post-hatch dispersal of lake sturgeon (Acipenser fulvescens, Rafinesque, 1817) yolk-sac larvae in relation to substrate in an artificial stream. J. Appl. Ichthyol. 2013, 29, 1208–1213. [Google Scholar] [CrossRef]
- Bauman, J.M.; Woodward, B.M.; Baker, E.A.; Marsh, T.L.; Scribner, K.T. Effects of Family, Feeding Frequency, and Alternate Food Type on Body Size and Survival of Hatchery-Produced and Wild-Caught Lake Sturgeon Larvae. N. Am. J. Aquac. 2016, 78, 136–144. [Google Scholar] [CrossRef]
- Callman, J.L.; Macy, J.M. The predominant anaerobe from the spiral intestine of hatchery-raised sturgeon (Acipenser transmontanus), a new Bacteroides species. Arch. Microbiol. 1984, 140, 57–65. [Google Scholar] [CrossRef]
- Buddington, R.K.; Christofferson, J.P. Digestive and feeding characteristics of the chondrosteans. Environ. Biol. Fishes 1985, 14, 31–41. [Google Scholar] [CrossRef]
- Milligan-Myhre, K.; Charette, J.R.; Phennicie, R.T.; Stephens, W.Z.; Rawls, J.F.; Guillemin, K.; Kim, C.H. Study of host-microbe interactions in zebrafish. In Methods in Cell Biology, 3rd ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2011. [Google Scholar]
- Kozich, J.J.; Westcott, S.L.; Baxter, N.T.; Highlander, S.K.; Schloss, P.D. Development of a dual-index sequencing strategy and curation pipeline for analyzing amplicon sequence data on the miseq illumina sequencing platform. Appl. Environ. Microbiol. 2013, 79, 5112–5120. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.; Guillaume Blanchet, F.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’hara, R. Package ‘Vegan’. Community Ecology Package, Version 2(9). 2015. Available online: https://github.com/vegandevs/vegan (accessed on 10 January 2022).
- Bray, J.R.; Curtis, J.T. An Ordination of the Upland Forest Communities of Southern Wisconsin. Ecol. Monogr. 1957, 27, 325–349. [Google Scholar] [CrossRef]
- Wickham, H. Ggplot2Applied Spatial Data Analysis with R; Springer: New York, NY, USA, 2009. [Google Scholar]
- Anderson, M. A new method for non parametric multivariate analysis of variance. Austral. Ecol. 2001, 26, 32–46. [Google Scholar]
- Anderson, M.J. Distance-based tests for homogeneity of multivariate dispersions. Biometrics 2006, 62, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [Green Version]
- Becattini, S.; Taur, Y.; Pamer, E.G. Antibiotic-Induced Changes in the Intestinal Microbiota and Disease. Trends Mol. Med. 2016, 22, 458–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manichanh, C.; Reeder, J.; Gibert, P.; Varela, E.; Llopis, M.; Antolin, M.; Guigo, R.; Knight, R.; Guarner, F. Reshaping the gut microbiome with bacterial transplantation and antibiotic intake. Genome Res. 2010, 20, 1411–1419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costello, E.K.; Stagaman, K.; Dethlefsen, L.; Bohannan, B.J.M.; Relman, D.A. The Application of Ecological Theory toward an Understanding of the Human Microbiome. Science 2012, 336, 1255–1262. [Google Scholar] [CrossRef] [Green Version]
- Panda, S.; El Khader, I.; Casellas, F.; López Vivancos, J.; García Cors, M.; Santiago, A.; Cuenca, S.; Guarner, F.; Manichanh, C. Short-term effect of antibiotics on human gut microbiota. PLoS ONE 2014, 9, e95476. [Google Scholar] [CrossRef]
- Huttenhower, C.; Gevers, D.; Knight, R.; Abubucker, S.; Badger, J.H.; Chinwalla, A.T.; Creasy, H.H.; Earl, A.M.; FitzGerald, M.G.; Fulton, R.S.; et al. Structure, function and diversity of the healthy human microbiome. Nature 2012, 486, 207–214. [Google Scholar]
- Dave, M.; Higgins, P.D.; Middha, S.; Rioux, K.P. The human gut microbiome: Current knowledge, challenges, and future directions. Transl. Res. 2012, 160, 246–257. [Google Scholar] [CrossRef]
- Goodrich, J.K.; Waters, J.L.; Poole, A.C.; Sutter, J.L.; Koren, O.; Blekhman, R. Human genetics shape the gut microbiome. Cell 2014, 159, 789–799. [Google Scholar] [CrossRef] [Green Version]
- Wilkins, D.; Leung, M.H.Y.; Lee, P.K.H. Indoor air bacterial communities in Hong Kong households assemble independently of occupant skin microbiomes. Environ. Microbiol. 2016, 18, 1754–1763. [Google Scholar] [CrossRef] [PubMed]
- Navarrete, P.; Toledo, I.; Mardones, P.; Opazo, R.; Espejo, R.; Romero, J. Effect of Thymus vulgaris essential oil on intestinal bacterial microbiota of rainbow trout, Oncorhynchus mykiss (Walbaum) and bacterial isolates. Aquac. Res. 2010, 41, e667–e678. [Google Scholar] [CrossRef]
- Romero, J.; Feijoó, C.G.; Navarrete, P. Antibiotics in Aquaculture—Use, Abuse and Alternatives. In Health and Environment in Aquaculture; Carvalho, D.E., Gianmarco, S.D., Reinaldo, J.S., Eds.; Books on Demand: Norderstedt, Germany, 2012; pp. 160–198. [Google Scholar]
- Slama, T.G. Gram-negative antibiotic resistance: There is a price to pay. Crit. Care 2008, 12, S4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasoo, S.; Barreto, J.N.; Tosh, P.K. Emerging Issues in Gram-Negative Bacterial Resistance: An Update for the Practicing Clinician. Mayo Clin. Proc. 2015, 90, 395–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacoby, G.A.; Munoz-Price, L.S. The New β-Lactamases. N. Engl. J. Med. 2005, 352, 380–391. [Google Scholar] [CrossRef] [PubMed]
- Lopetuso, L.R.; Scaldaferri, F.; Petito, V.; Gasbarrini, A. Commensal Clostridia: Leading players in the maintenance of gut homeostasis. Gut Pathog. 2013, 5, 23. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R.S.; Gao, B. Phylogenomic analyses of clostridia and identification of novel protein signatures that are specific to the genus Clostridiumsensu stricto (cluster I). Int. J. Syst. Evol. Microbiol. 2009, 59, 285–294. [Google Scholar] [CrossRef] [Green Version]
- Palmer, C.; Bik, E.M.; DiGiulio, D.B.; Relman, D.A.; Brown, P.O. Development of the human infant intestinal microbiota. PLoS Biol. 2007, 5, e177. [Google Scholar] [CrossRef] [Green Version]
- De Schryver, P.; Vadstein, O. Ecological theory as a foundation to control pathogenic invasion in aquaculture. ISME J. 2014, 8, 2360–2368. [Google Scholar] [CrossRef] [Green Version]
- Gisbert, E.; Sarasquete, M.C.; Williot, P.; Castelló-Orvay, F. Histochemistry of the development of the digestive system of Siberian sturgeon during early ontogeny. J. Fish Biol. 1999, 55, 596–616. [Google Scholar] [CrossRef]
- Berois, N.; Arezo, M.J.; Papa, N.G. Gamete interactions in teleost fish: The egg envelope. Basic studies and perspectives as environmental biomonitor. Biol. Res. 2011, 44, 119–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sevellec, M.; Laporte, M.; Bernatchez, A.; Derome, N.; Bernatchez, L. Evidence for host effect on the intestinal microbiota of whitefish (Coregonus sp.) species pairs and their hybrids. Ecol. Evol. 2019, 9, 11762–11774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarrete, P.; Magne, F.; Araneda, C.; Fuentes, P.; Barros, L.; Opazo, R.; Espejo, R.; Romero, J. PCR-TTGE analysis of 16S rRNA from rainbow trout (Oncorhynchus mykiss) gut microbiota reveals host-specific communities of active bacteria. PLoS ONE 2012, 7, e31335. [Google Scholar]
- Zhao, L.; Wang, G.; Siegel, P.; He, C.; Wang, H.; Zhao, W.; Zhai, Z.; Tian, F.; Zhao, J.; Zhang, H.; et al. Quantitative Genetic Background of the Host Influences Gut Microbiomes in Chickens. Sci. Rep. 2013, 3, 1163. [Google Scholar] [CrossRef] [PubMed]
- Blekhman, R.; Goodrich, J.K.; Huang, K.; Sun, Q.; Bukowski, R.; Bell, J.T.; Spector, T.D.; Keinan, A.; Ley, R.E.; Gevers, D.; et al. Host genetic variation impacts microbiome composition across human body sites. Genome Biol. 2015, 16, 191. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Df | Sum Sq | Mean Sq | F-Model | R2 | Pr (>F) | |
|---|---|---|---|---|---|---|
| Origin (O) | 1 | 0.416 | 0.416 | 1.792 | 0.050 | 0.107 |
| Residuals | 34 | 7.887 | 0.232 | 0.950 | ||
| Total | 35 | 8.302 | 1.000 |
| Df | Sum Sq | Mean Sq | F-Model | R2 | Pr (>F) | |
|---|---|---|---|---|---|---|
| Treatment (T) | 3 | 0.897 | 0.299 | 1.389 | 0.028 | 0.126 |
| Origin (O) | 1 | 1.699 | 1.699 | 7.900 | 0.052 | p < 0.01 |
| Residuals | 139 | 29.900 | 0.215 | 0.920 | ||
| Total | 143 | 32.496 | 1.000 |
| Df | Sum Sq | Mean Sq | F-Model | R2 | Pr (>F) | |
|---|---|---|---|---|---|---|
| Treatment (T) | 3 | 1.108 | 0.369 | 1.913 | 0.076 | 0.012 |
| Family (F) | 1 | 0.402 | 0.402 | 2.084 | 0.028 | 0.057 |
| Treatment (T) × Family (F) | 3 | 0.737 | 0.246 | 1.272 | 0.050 | 0.191 |
| Residuals | 64 | 12.357 | 0.193 | 0.846 | ||
| Total | 71 | 14.603 | 1.000 |
| Df | Sum Sq | Mean Sq | F-Model | R2 | Pr (>F) | |
|---|---|---|---|---|---|---|
| Treatment (T) | 3 | 0.784 | 0.261 | 1.241 | 0.048 | 0.209 |
| Family (F) | 1 | 0.539 | 0.539 | 2.560 | 0.033 | 0.022 |
| Treatment (T) × Family (F) | 3 | 1.390 | 0.463 | 2.202 | 0.086 | 0.007 |
| Residuals | 64 | 13.471 | 0.210 | 0.832 | ||
| Total | 71 | 16.184 | 1.000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdul Razak, S.; Bauman, J.M.; Marsh, T.L.; Scribner, K.T. Changes in Lake Sturgeon Gut Microbiomes Relative to Founding Origin and in Response to Chemotherapeutant Treatments. Microorganisms 2022, 10, 1005. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10051005
Abdul Razak S, Bauman JM, Marsh TL, Scribner KT. Changes in Lake Sturgeon Gut Microbiomes Relative to Founding Origin and in Response to Chemotherapeutant Treatments. Microorganisms. 2022; 10(5):1005. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10051005
Chicago/Turabian StyleAbdul Razak, Shairah, John M. Bauman, Terence L. Marsh, and Kim T. Scribner. 2022. "Changes in Lake Sturgeon Gut Microbiomes Relative to Founding Origin and in Response to Chemotherapeutant Treatments" Microorganisms 10, no. 5: 1005. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10051005