
Diversity, Abundance and Leishmania infantum Infection Rate of Phlebotomine Sandflies in an Area with Low Incidence of Visceral Leishmaniasis in Northern Tunisia

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
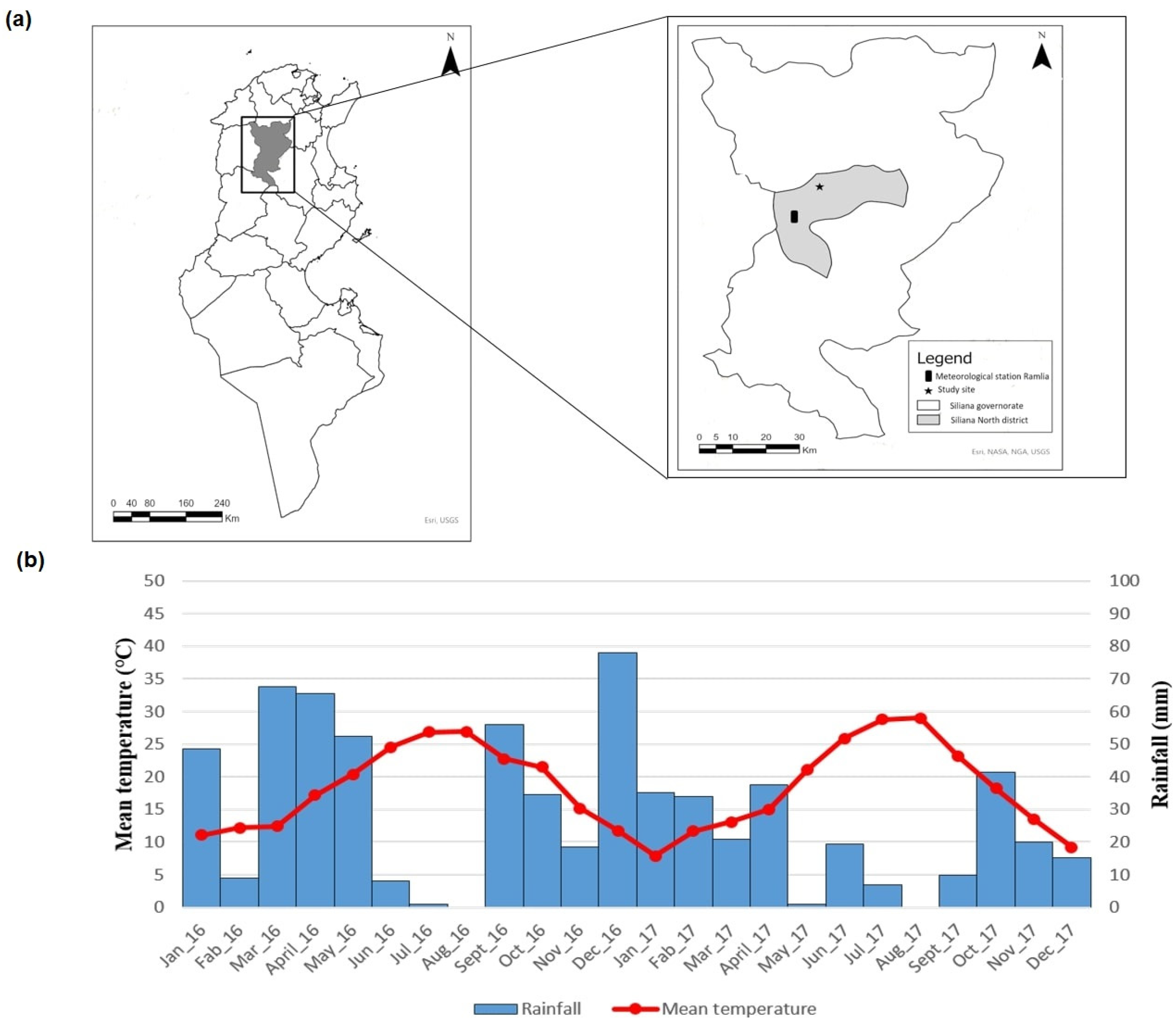
2.1. Study Area
2.2. Sandfly Collection, Habitat, and Physiological State of Female Specimens
2.3. Sandfly Species Identification and Study of Leishmania Infection
2.3.1. Molecular Screening of Leishmania Infection by kDNA qPCR
2.3.2. Identification of Leishmania Species by ITS1 PCR Sequencing
2.3.3. Identification of Sandfly Species by DNA Barcoding
2.4. Data Analysis
2.4.1. Species Diversity
2.4.2. Statistical Analysis
3. Results
3.1. Sandfly Fauna in the Study Site
3.2. Species Diversity
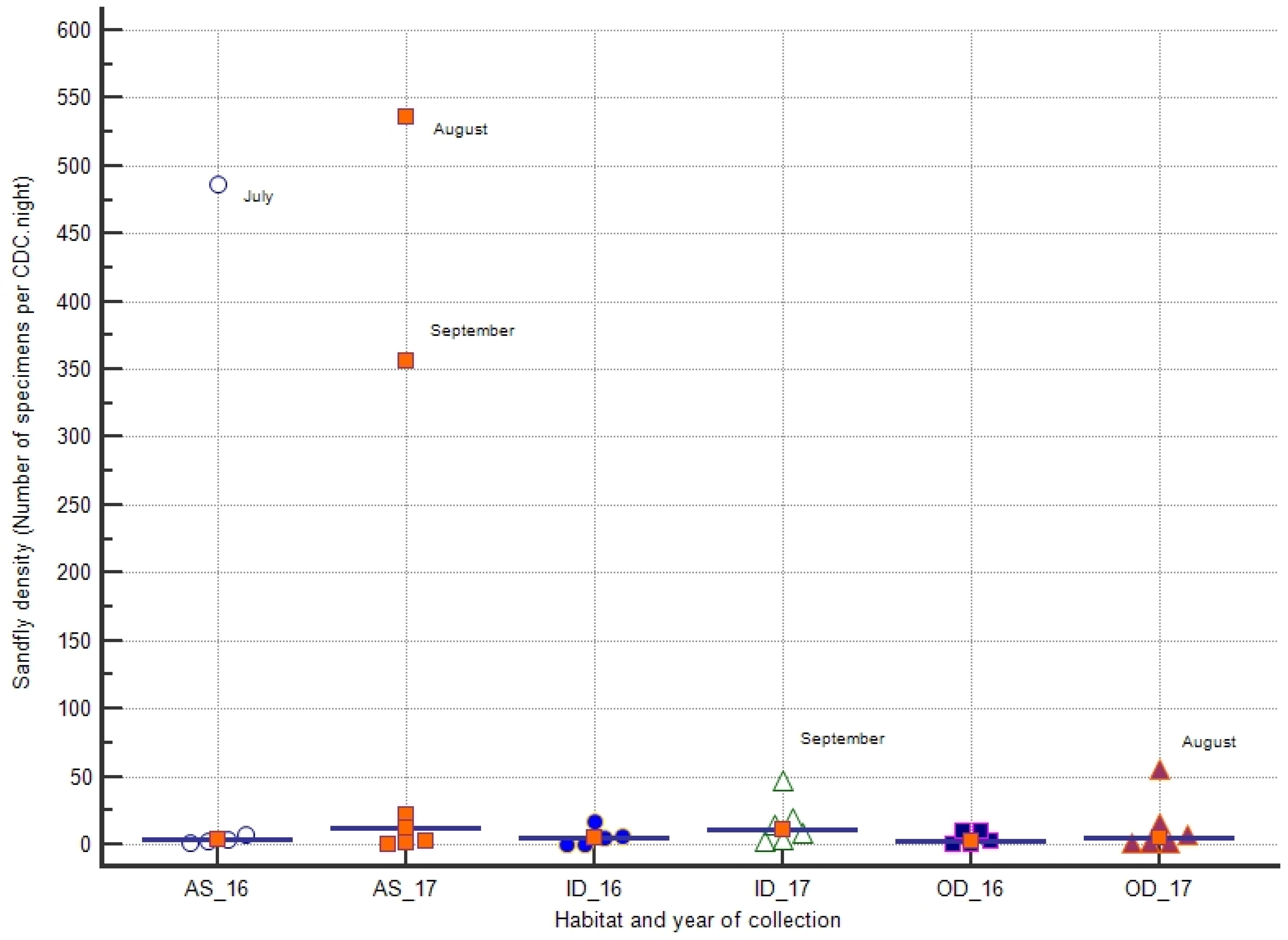
3.3. Sandfly Density
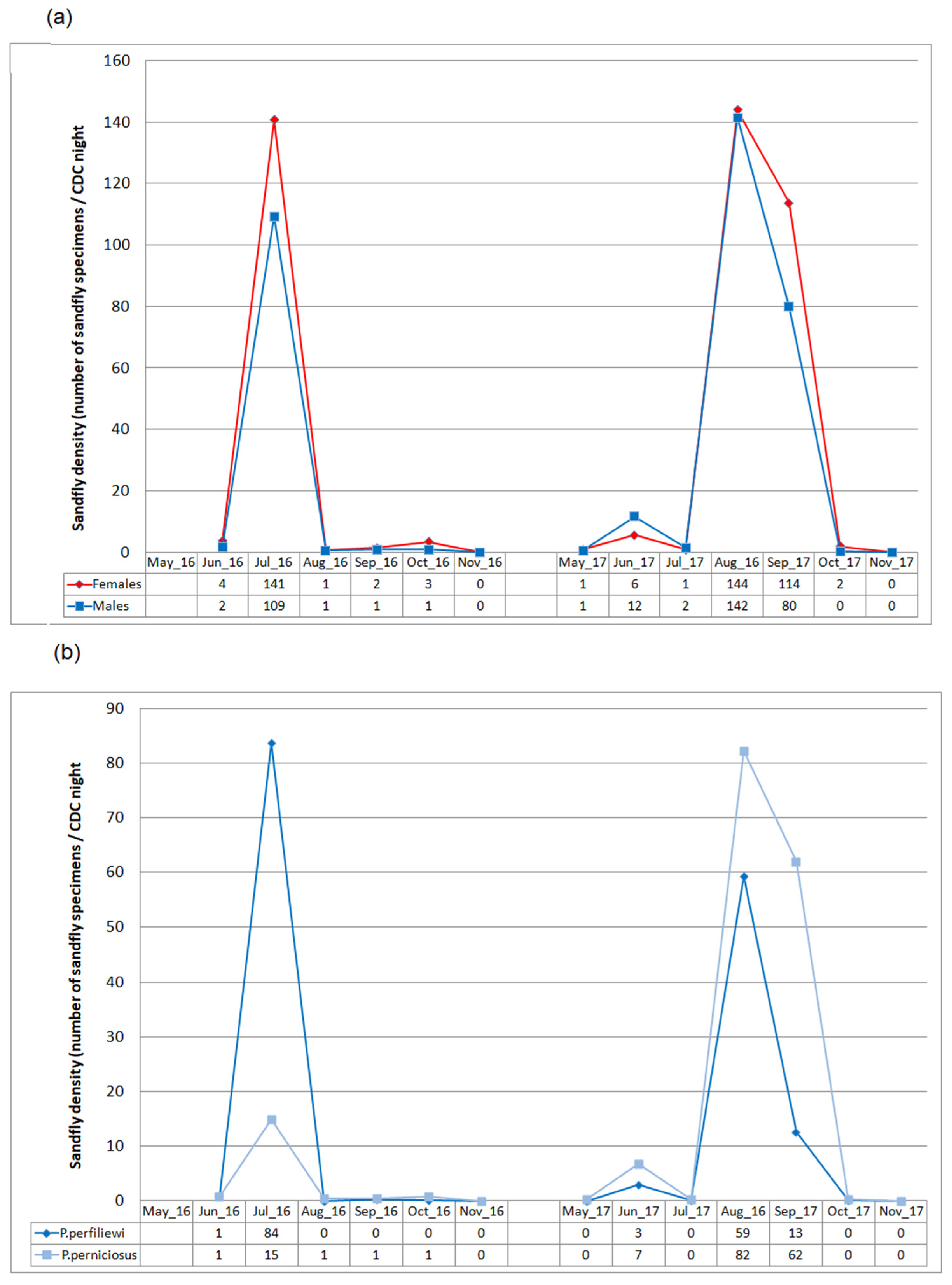
3.4. Seasonal Dynamics
3.5. Leishmania Infection
3.6. Leishmania Infantum Infection and Parasitic Load
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Pratlong, F.; Lami, P.; Ravel, C.; Balard, Y.; Dereure, J.; Serres, G.; El-Baidouri, F.; Dedet, J.P. Geographical distribution and epidemiological features of Old World Leishmania infantum and Leishmania donovani foci, based on the isoenzyme analysis of 2277 strains. Parasitology 2013, 140, 423–434. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization (W.H.O). Control of the Leishmaniases: Report of a Meeting of the WHO Expert Commitee on the Control of Leishmaniases, Geneva, 22–26 March 2010; WHO Technical Report Series; World Health Organization: Geneva, Switzerland, 2010. [Google Scholar]
- Gramiccia, M.; Gradoni, L. The current status of zoonotic leishmaniases and approaches to disease control. Int. J. Parasitol. 2005, 35, 1169–1180. [Google Scholar] [CrossRef]
- Killick-Kendrick, R. Phlebotomine vectors of the leishmaniases: A review. Med. Vet. Entomol. 1990, 4, 1–24. [Google Scholar] [CrossRef]
- Rioux, J.A.; Killick-Kendrick, R.; Leaney, A.J.; Young, C.J.; Turner, D.P.; Lanotte, G.; Bailly, M. Ecology of leishmaniasis in the south of France. 11. Canine leishmaniasis: Successful experimental transmission from dog to dog by the bite of Phlebotomus ariasi Tonnoir, 1921. Ann. Parasitol. Hum. Comp. 1979, 54, 401–407. [Google Scholar] [CrossRef] [Green Version]
- Bettini, S.; Gramiccia, M.; Gradoni, L.; Atzeni, M.C. Leishmaniasis in Sardinia: II. Natural infection of Phlebotomus perniciosus Newstead, 1911, by Leishmania infantum Nicolle, 1908, in the province of Cagliari. Trans. R. Soc. Trop. Med. Hyg. 1986, 80, 458–459. [Google Scholar] [CrossRef]
- Maroli, M.; Gramiccia, M.; Gradoni, L. Natural infection of Phlebotomus perfiliewi with Leishmania infantum in a cutaneous leishmaniasis focus of the Abruzzi region, Italy. Trans. R. Soc. Trop. Med. Hyg. 1987, 8, 596–598. [Google Scholar] [CrossRef]
- Alten, B.; Maia, C.; Afonso, M.O.; Campino, L.; Jiménez, M.; González, E.; Molina, R.; Bañuls, A.L.; Prudhomme, J.; Vergnes, B.; et al. Seasonal dynamics of phlebotomine sandfly species proven vectors of Mediterranean leishmaniasis caused by Leishmania infantum. PLoS Negl. Trop. Dis. 2016, 10, e0004458. [Google Scholar] [CrossRef]
- Aoun, K.; Amri, F.; Chouihi, E.; Haouas, N.; Bedoui, K.; Benikhlef, R.; Ghrab, J.; Babba, H.; Chahed, M.K.; Harrat, Z.; et al. Épidémiologie de Leishmania (L.) infantum, L. major et L. killicki en Tunisie: Résultats et analyse de l’identification de 226 isolats humains et canins et revue de la littérature. Bull. Soc. Pathol. Exot. 2008, 101, 323–328. [Google Scholar] [CrossRef]
- Aoun, K.; Bouratbine, A. Cutaneous leishmaniasis in North Africa: A review. Parasite 2014, 21, 14. [Google Scholar] [CrossRef] [Green Version]
- Aoun, K.; Jeddi, F.; Amri, F.; Ghrab, J.; Bouratbine, A. Current epidemiological data on visceral leishmaniasis in Tunisia. Med. Mal. Infect. 2009, 39, 775–779. [Google Scholar] [CrossRef]
- Ben-Ahmed, K.; Aoun, K.; Jeddi, F.; Ghrab, J.; El-Aroui, M.A.; Bouratbine, A. Visceral leishmaniasis in Tunisia: Spatial distribution and association with climatic factors. Am. J. Trop. Med. Hyg. 2009, 81, 40–45. [Google Scholar] [CrossRef]
- Benabid, M.; Ghrab, J.; Rhim, A.; Ben-Romdhane, R.; Aoun, K.; Bouratbine, A. Temporal dynamics and Leishmania infantum infection prevalence of Phlebotomus perniciosus (Diptera, Phlebotominae) in highly endemic areas of visceral leishmaniasis in Tunisia. PLoS ONE 2017, 12, 0184700. [Google Scholar] [CrossRef] [Green Version]
- Ben-Salah, A.; Ben-Ismail, R.; Amri, F.; Chlif, S.; Ben-Rzig, F.; Kharrat, H.; Hadhri, H.; Hassouna, M.; Dellagi, K. Investigation of the spread of human visceral leishmaniasis in central Tunisia. Trans. R. Soc. Trop. Med. Hyg. 2000, 94, 382–386. [Google Scholar] [CrossRef]
- Barhoumi, W.; Fares, W.; Cherni, S.; Derbali, M.; Dachraoui, K.; Chelbi, I.; Ramalho-Ortigao, M.; Beier, J.C.; Zhioua, E. Changes of sand fly populations and Leishmania infantum infection rates in an irrigated village located in arid Central Tunisia. Int. J. Environ. Res. Public Health 2016, 13, 329. [Google Scholar] [CrossRef] [Green Version]
- Remadi, L.; Chargui, N.; Jimenez, M.; Molina, R.; Haouas, N.; González, E.; Chaabane-Banaouas, R.; Ben Salah, E.; Haddaji, M.; Chaabouni, Y.; et al. Molecular detection and identification of Leishmania DNA and blood meal analysis in Phlebotomus (Larroussius) species. PLoS Negl. Trop. Dis. 2020, 14, e0008077. [Google Scholar] [CrossRef]
- Chaouch, M.; Chaabane, A.; Ayari, C.; Ben-Othman, S.; Sereno, D.; Chemkhi, J.; BenAbderrazak, S. Investigation of natural infection of Phlebotomine (Diptera: Psychodidae) by Leishmania in Tunisian endemic regions. Parasite Epidemiol. Control 2021, 14, e00212. [Google Scholar] [CrossRef]
- Ghrab, J.; Rhim, A.; Bach-Hamba, D.; Chahed, M.K.; Aoun, K.; Nouira, S.; Bouratbine, A. Phlebotominae (Diptera: Psychodidae) of human leishmaniosis sites in Tunisia. Parasite 2006, 13, 23–33. [Google Scholar] [CrossRef] [Green Version]
- Galluzzi, L.; Ceccarelli, M.; Diotallevi, A.; Menotta, M.; Magnani, M. Real-time PCR applications for diagnosis of leishmaniasis. Parasites Vectors 2018, 11, 273. [Google Scholar] [CrossRef]
- Myskova, J.; Votypka, J.; Volf, P. Leishmania in sandflies: Comparison of quantitative polymerase chain reaction with other techniques to determine the intensity of infection. J. Med. Entomol. 2008, 45, 133–138. [Google Scholar] [CrossRef]
- Gonzalez, E.; Alvarez, A.; Ruiz, S.; Molina, R.; Jimenez, M. Detection of high Leishmania infantum loads in Phlebotomus perniciosus captured in the leishmaniasis focus of southwestern Madrid region (Spain) by real time PCR. Acta Trop. 2017, 171, 68–73. [Google Scholar] [CrossRef]
- Dolmatova, A.V.; Denima, N.A. Les phlébotomes (Phlebotominae) et Maladies qu’ils Transmettent; Documentation Technique; O.R.S.T.O.M.: Paris, France, 1971; p. 169. [Google Scholar]
- Torina, A.; Sole, M.; Reale, S.; Vitale, F.; Caracappa, S. Use of phlebotomine sandflies as indicator of Leishmania prevalence in an endemic area. Ann. N. Y. Acad. Sci. 2008, 1149, 355–357. [Google Scholar] [CrossRef]
- Croset, H.; Rioux, J.A.; Maistre, M.; Bayar, N. Les Phlébotomes de Tunisie (Diptera, Phlebotomidae). Mise au point systématique, chorologique et éthologique. Ann. Parasitol. Hum. Comp. 1978, 53, 711–749. [Google Scholar] [CrossRef] [Green Version]
- Ready, P.D.; Lainson, R.; Shaw, J.J.; Souza, A.A. DNA probes for distinguishing Psychodopygus wellcomei from Psychodopygus complexus (Diptera: Psychodidae). Mem. Inst. Oswaldo Cruz 1991, 86, 41–49. [Google Scholar] [CrossRef] [Green Version]
- Mary, C.; Faraut, F.; Lascombe, L.; Dumon, H. Quantification of Leishmania infantum DNA by a real-time PCR assay with high sensitivity. J. Clin. Microbiol. 2004, 42, 5249–5255. [Google Scholar] [CrossRef] [Green Version]
- Schönian, G.; Nasereddin, A.; Dinse, N.; Schweynoch, C.; Schallig, H.D.; Presber, W.; Jaffe, C.L. PCR diagnosis and characterization of Leishmania in local and imported clinical samples. Diagn. Microbiol. Infect. Dis. 2003, 47, 349–358. [Google Scholar] [CrossRef]
- Kumar, N.P.; Srinivasan, R.; Jambulingam, P. DNA barcoding for identification of sandflies (Diptera: Psychodidae) in India. Mol. Ecol. Res. 2012, 12, 414–420. [Google Scholar] [CrossRef]
- De Jong, T.M. A Comparison of three diversity indices based on their components of richness and evenness. Oikos 1975, 26, 222–227. [Google Scholar] [CrossRef]
- Mouillot, D.; Leprêtre, A.A. Comparison of species diversity estimators. Rese. Popul. Ecol. 1999, 41, 203–215. [Google Scholar]
- Prudhomme, J.; Rahola, N.; Toty, C.; Cassan, C.; Roiz, D.; Vergnes, B.; Thierry, M.; Rioux, J.A.; Alten, B.; Sereno, D.; et al. Ecology and spatiotemporal dynamics of sandflies in the Mediterranean Languedoc region (Roquedur area, Gard, France). Parasites Vectors 2015, 8, 642. [Google Scholar] [CrossRef] [Green Version]
- Alexander, B. Sampling methods for phlebotomine sandflies. Med. Vet. Entomol. 2000, 14, 109–122. [Google Scholar] [CrossRef]
- Rispail, P.; Dereure, J.; Jarry, D. Risk zones of human Leishmaniases in the Western Mediterranean basin: Correlations between vector sandflies, bioclimatology and phytosociology. Mem. Inst. Oswaldo Cruz 2002, 97, 477–483. [Google Scholar] [CrossRef] [Green Version]
- Norouzi, B.; Hanafi-Bojd, A.A.; Moin-Vaziri, V.; Noorallahi, A.; Azari-Hamidian, S. Ecology of sand flies (Diptera: Psychodidae, Phlebotominae) in a new focus of leishmaniasis in northern Iran. Acta Trop. 2020, 212, 105649. [Google Scholar] [CrossRef]
- Barhoumi, W.; Qualls, W.A.; Archer, R.S.; Fuller, D.O.; Chelbi, I.; Cherni, S.; Derbali, M.; Arheart, K.L.; Zhioua, E.; Beier, J.C. Irrigation in the arid regions of Tunisia impacts the abundance and apparent density of sand fly vectors of Leishmania infantum. Acta Trop. 2015, 141, 73–78. [Google Scholar] [CrossRef] [Green Version]
- Alcover, M.M.; Ballart, C.; Martín-Sánchez, J.; Serra, T.; Castillejo, S.; Portús, M.; Gállego, M. Factors influencing the presence of sandflies in Majorca (Balearic Islands, Spain) with special reference to Phlebotomus pernicious, vector of Leishmania infantum. Parasites Vectors 2014, 7, 421. [Google Scholar] [CrossRef] [Green Version]
- Rossi, E.; Bongiorno, G.; Ciolli, E.; Di Muccio, T.; Scalone, A.; Gramiccia, M.; Gradoni, L.; Maroli, M. Seasonal phenology, host-blood feeding preferences and natural Leishmania infection of Phlebotomus perniciosus (Diptera, Psychodidae) in a high-endemic focus of canine leishmaniasis in Rome province, Italy. Acta Trop. 2008, 105, 158–165. [Google Scholar] [CrossRef]
- Cazan, C.D.; Păstrav, I.R.; Györke, A.; Oguz, G.; Alten, B.; Mihalca, A.D. Seasonal dynamics of a population of Phlebotomus (Larroussius) perfiliewi Parrot, 1930 (Diptera: Psychodidae) in North-Eastern Romania. Parasitol. Res. 2019, 118, 1371–1384. [Google Scholar] [CrossRef]
- Tabbabi, A.; Ghrab, J.; Aoun, K.; Ready, P.D.; Bouratbine, A. Habitats of the sandfly vectors of Leishmania tropica and L. major in a mixed focus of cutaneous leishmaniasis in southeast Tunisia. Acta Trop. 2011, 119, 131–137. [Google Scholar] [CrossRef]
- Risueño, J.; Muñoz, C.; Pérez-Cutillas, P.; Goyena, E.; Gonzálvez, M.; Ortuño, M.; Bernal, L.J.; Ortiz, J.; Alten, B.; Berriatua, E. Understanding Phlebotomus perniciosus abundance in south-east Spain: Assessing the role of environmental and anthropic factors. Parasites Vectors 2017, 10, 189. [Google Scholar] [CrossRef] [Green Version]
- Croset, H.; Rioux, J.A.; Juminer, B.; Tour, S. Annual fluctuations of populations of Phlebotomus perniciosus Newstead, 1911, Phlebotomus perfiliewi Parrot, 1930 and Sergentomyia minuta parroti (Adler and Theodor, 1927)(Diptera-Psychodidae) in northern Tunisia. Arch. Inst. Pasteur Tunis 1970, 47, 43–56. [Google Scholar]
- Ballart, C.; Guerrero, I.; Castells, X.; Barón, S.; Castillejo, S.; Alcover, M.M.; Portús, M.; Gállego, M. Importance of individual analysis of environmental and climatic factors affecting the density of Leishmania vectors living in the same geographical area: The example of Phlebotomus ariasi and P. perniciosus in northeast Spain. Geosp. Health 2014, 8, 389–403. [Google Scholar] [CrossRef] [Green Version]
- Boutsini, S.; Athanasiou, L.V.; Spanakos, G.; Ntousi, D.; Dotsika, E.; Bisia, M.; Papadopoulos, E. Phlebotomine sandflies and factors associated with their abundance in the leishmaniasis endemic area of Attiki, Greece. Parasitol. Res. 2018, 117, 107–113. [Google Scholar] [CrossRef]
- Bennai, K.; Tahir, D.; Lafri, I.; Bendjaballah-Laliam, A.; Bitam, I.; Parola, P. Molecular detection of Leishmania infantum DNA and host blood meal identification in Phlebotomus in a hypoendemic focus of human leishmaniasis in northern Algeria. PLoS Negl. Trop. Dis. 2018, 12, e0006513. [Google Scholar] [CrossRef]
- Marcelino, A.P.; de Souza Filho, J.A.; e Bastos, C.D.V.; Ribeiro, S.R.; Medeiros, F.A.C.; Reis, I.A.; da Rocha, A.C.V.M.; Barbosa, J.R.; Paz, G.F.; Gontijo, C.M.F. Comparative PCR-based diagnosis for the detection of Leishmania infantum in naturally infected dogs. Acta Trop. 2020, 207, 105495. [Google Scholar] [CrossRef]
- Bensoussan, E.; Nasereddin, A.; Jonas, F.; Schnur, L.F.; Jaffe, C.L. Comparison of PCR assays for diagnosis of cutaneous leishmaniasis. J. Clin. Microbiol. 2006, 44, 1435–1439. [Google Scholar] [CrossRef] [Green Version]
- González, E.; Molina, R.; Jiménez, M. Rabbit trypanosome detection in Phlebotomus perniciosus sand flies from the leishmaniasis outbreak in Madrid, Spain. Acta Trop. 2018, 187, 201–206. [Google Scholar] [CrossRef]
- Svobodova, M.; Radrova, J. Phlebotomine Sandflies-Potential Vectors of Avian Trypanosomes. Acta Protozool. 2018, 57, 53–59. [Google Scholar]
- Kocher, A.; Valiere, S.; Banuls, A.L.; Murienne, J. High-throughput sequencing of kDNA amplicons for the analysis of Leishmania minicircles and identification of Neotropical species. Parasitology 2018, 145, 585–594. [Google Scholar] [CrossRef]
- Bussotti, G.; Benkahla, A.; Jeddi, F.; Souiaï, O.; Aoun, K.; Späth, G.F.; Bouratbine, A. Nuclear and mitochondrial genome sequencing of North-African Leishmania infantum isolates from cured and relapsed visceral leishmaniasis patients reveals variations correlating with geography and phenotype. Microb. Genom. 2020, 6, mgen000444. [Google Scholar] [CrossRef]
- Killick-Kendrick, R.; Rioux, J.A. Mark-release-recapture of sandflies fed on leishmanial dogs: The natural life-cycle of Leishmania infantum in Phlebotomus ariasi. Parassitologia 2002, 44, 67–71. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Habitat | Number CDC.night | Number Males | Number Females | Number Sandflies | Sex Ratio |
|---|---|---|---|---|---|---|
| 2016 | ID | 12 | 22 | 37 | 59 | 0.59 |
| OD | 12 | 13 | 27 | 40 | 0.48 | |
| AS | 24 | 871 | 1136 | 2007 | 0.77 | |
| Total | 48 | 906 | 1200 | 2106 | 0.75 | |
| 2017 | ID | 14 | 111 | 78 | 189 | 1.42 |
| OD | 14 | 60 | 104 | 164 | 0.58 | |
| AS | 28 | 1712 | 1954 | 3666 | 0.88 | |
| Total | 56 | 1883 | 2136 | 4019 | 0.88 | |
| Total | 104 | 2789 | 3336 | 6125 | 0.84 |
| Year | Habitat | Number Females | Number Unfed (%) | Number Fed (%) | Number Gravid (%) | |
|---|---|---|---|---|---|---|
| Fresh BM | Digested BM | |||||
| 2016 | ID | 37 | 30 (81%) | 6 (16.3%) | 1 (2.7%) | |
| 1 | 5 | |||||
| OD | 27 | 23 (85.2%) | 4 (14.8%) | 0 (0%) | ||
| 1 | 3 | |||||
| AS | 1136 | 888 (78.2%) | 246 (21.6%) | 2 (0.2%) | ||
| 123 | 123 | |||||
| Total | 1200 | 941 (78.4%) | 256 (21.3%) | 3 (0.3%) | ||
| 125 | 131 | |||||
| 2017 | ID | 78 | 49 (62.8%) | 27 (34.6%) | 2 (2.6%) | |
| 11 | 16 | |||||
| OD | 104 | 86 (82.7%) | 15 (14.4%) | 3 (2.9%) | ||
| 7 | 8 | |||||
| AS | 1954 | 1499(76.7%) | 437 (22.4%) | 18 (0.9%) | ||
| 267 | 170 | |||||
| Total | 2136 | 1634 (76.5%) | 479 (22.4%) | 23 (1.1%) | ||
| 285 | 194 | |||||
| Year | Habitat | Abundance (pi) | Diversity Indices | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| P. perf | P. pern | P. long | P. pap | P. serg | S. ant | S | H’ | H’/H’max | D | D/Dmax | ||
| 2016 | ID | 10 (0.45) | 5 (0.23) | - | 7 (0.32) | - | - | 3 | 1.54 | 0.96 | 0.67 | 1.00 |
| OD | 6 (0.46) | 7 (0.54) | - | - | - | - | 2 | 1.00 | 1.00 | 0.54 | 1.00 | |
| AS | 665 (0.76) | 127 (0.15) | - | - | 40 (0.05) | 39 (0.04) | 4 | 1.11 | 0.55 | 0.39 | 0.52 | |
| 2017 | ID | 1 (0.01) | 100 (0.90) | - | 2 (0.02) | 5 (0.05) | 3 (0.03) | 5 | 0.65 | 0.28 | 0.19 | 0.23 |
| OD | 57 (0.95) | 1 (0.02) | - | - | - | 2 (0.03) | 3 | 0.33 | 0.21 | 0.10 | 0.15 | |
| AS | 542 (0.32) | 1115 (0.65) | 3 (0.00) | - | 3(0.00) | 49 (0.03) | 5 | 1.11 | 0.48 | 0.48 | 0.59 | |
| Number of Specimens in 2016 | Number of Specimens in 2017 | ||||||
|---|---|---|---|---|---|---|---|
| Collected | DNA | + kDNA qPCR | Collected | DNA | + kDNA qPCR (Infection Rate) | ||
| Habitat | ID | 37 | 32 | 0 | 78 | 73 | 1 (1.36%) |
| OD | 27 | 24 | 0 | 104 | 103 | 0 | |
| AS | 1136 | 255 | 0 | 1954 | 391 | 3 (0.76%) | |
| Month of capture | May | - | - | - | 7 | 7 | 0 |
| June | 28 | 28 | 0 | 42 | 42 | 0 | |
| July | 1128 | 239 | 0 | 7 | 7 | 0 | |
| August | 5 | 5 | 0 | 1151 | 275 | 0 | |
| September | 12 | 12 | 0 | 912 | 223 | 4 (0.44%) | |
| October | 27 | 27 | 0 | 17 | 13 | 0 | |
| November | 0 | 0 | - | 0 | 0 | 0 | |
| Physiological state | Unfed | 941 | 178 | 0 | 1634 | 304 | 0 |
| Fed | 256 | 130 | 0 | 479 | 240 | 2 (0.83%) | |
| Gravid | 3 | 3 | 0 | 23 | 23 | 2 (8.69%) | |
| Blood meal | Fresh | 125 | 48 | 0 | 285 | 87 | 2 (2.29%) |
| Digested | 131 | 82 | 0 | 194 | 153 | 0 | |
| Total female specimens | 1200 | 311 | 0 | 2136 | 567 | 4 (0.70%) | |
| Code | Habitat | Physiological State | Month of Capture | Estimated Number of Parasites per Sandfly Female Specimen | ITS1 PCR Result | Leishmania Species |
|---|---|---|---|---|---|---|
| Dj033E10 | AS | Gravid | Sept 2017 | 2 parasites | Negative | |
| Dj033F4 | AS | Gravid | Sept 2017 | 38,000 parasites | Positive | L. infatum |
| Dj032B1 | AS | Fed | Sept 2017 | 4 parasites | Negative | |
| Dj030F1 | ID | Fed | Sept 2017 | 18 parasites | Negative |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weslati, M.; Ghrab, J.; Benabid, M.; Souissi, O.; Aoun, K.; Bouratbine, A. Diversity, Abundance and Leishmania infantum Infection Rate of Phlebotomine Sandflies in an Area with Low Incidence of Visceral Leishmaniasis in Northern Tunisia. Microorganisms 2022, 10, 1012. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10051012
Weslati M, Ghrab J, Benabid M, Souissi O, Aoun K, Bouratbine A. Diversity, Abundance and Leishmania infantum Infection Rate of Phlebotomine Sandflies in an Area with Low Incidence of Visceral Leishmaniasis in Northern Tunisia. Microorganisms. 2022; 10(5):1012. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10051012
Chicago/Turabian StyleWeslati, Marwa, Jamila Ghrab, Meriem Benabid, Olfa Souissi, Karim Aoun, and Aïda Bouratbine. 2022. "Diversity, Abundance and Leishmania infantum Infection Rate of Phlebotomine Sandflies in an Area with Low Incidence of Visceral Leishmaniasis in Northern Tunisia" Microorganisms 10, no. 5: 1012. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10051012