
Distinct Elevational Patterns and Their Linkages of Soil Bacteria and Plant Community in An Alpine Meadow of the Qinghai–Tibetan Plateau


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites and Sampling
2.2. Measurement of Soil Physicochemical Characteristics
2.3. Climate Data
2.4. Soil Microbial DNA Extraction, Purification, and Quantification
2.5. Illumina Sequencing of Bacterial 16S rRNA Gene Amplicons and Data Processing
2.6. GeoChip Hybridization and Data Processing
2.7. Statistical Analysis
3. Results
3.1. Climate and Soil Physicochemical Characteristics
3.2. Distribution Pattern of Plant Diversity along Elevation Gradient
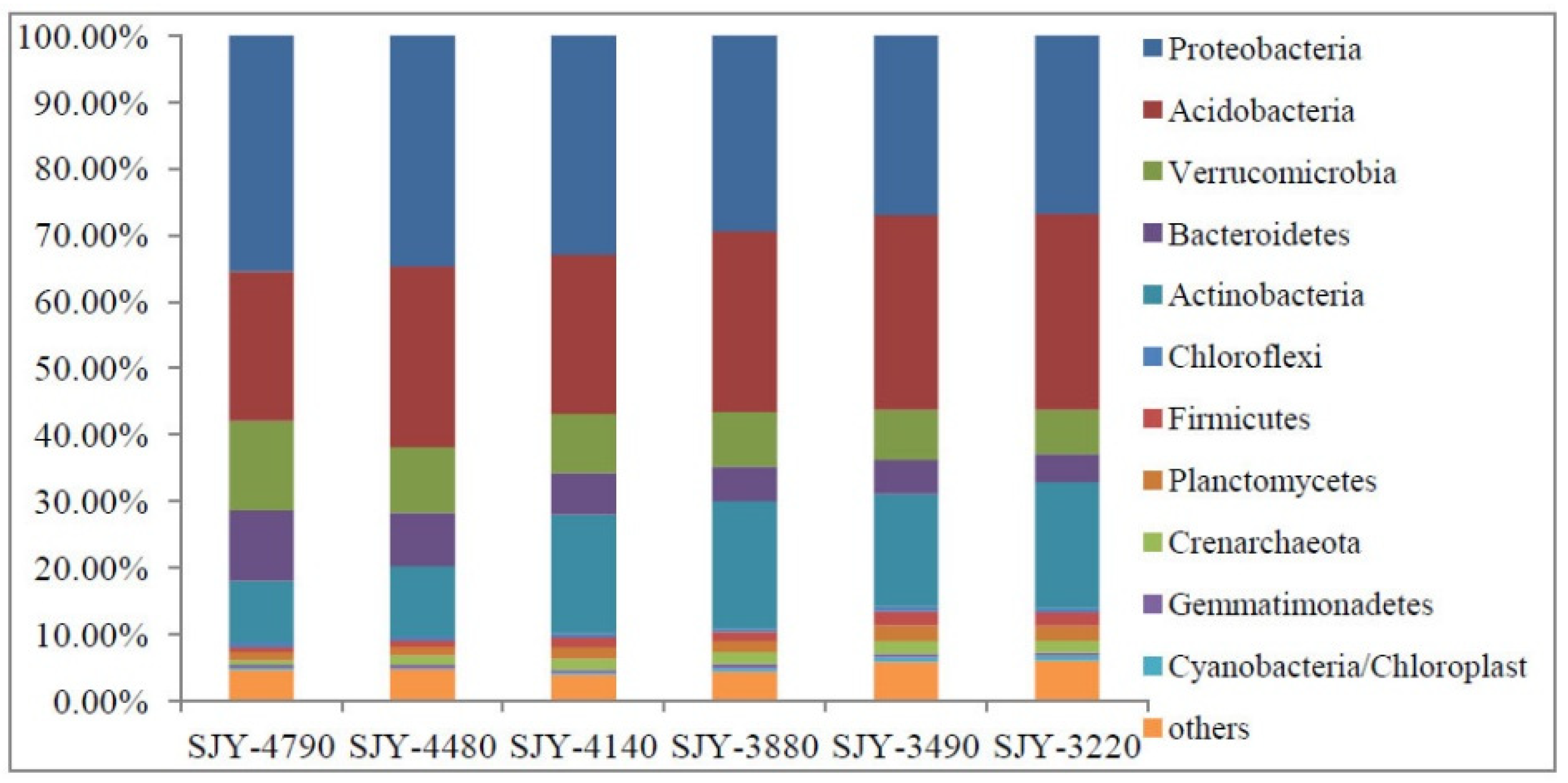
3.3. Distribution Pattern of Soil Community Structure and Bacterial Diversity along Elevation Gradient
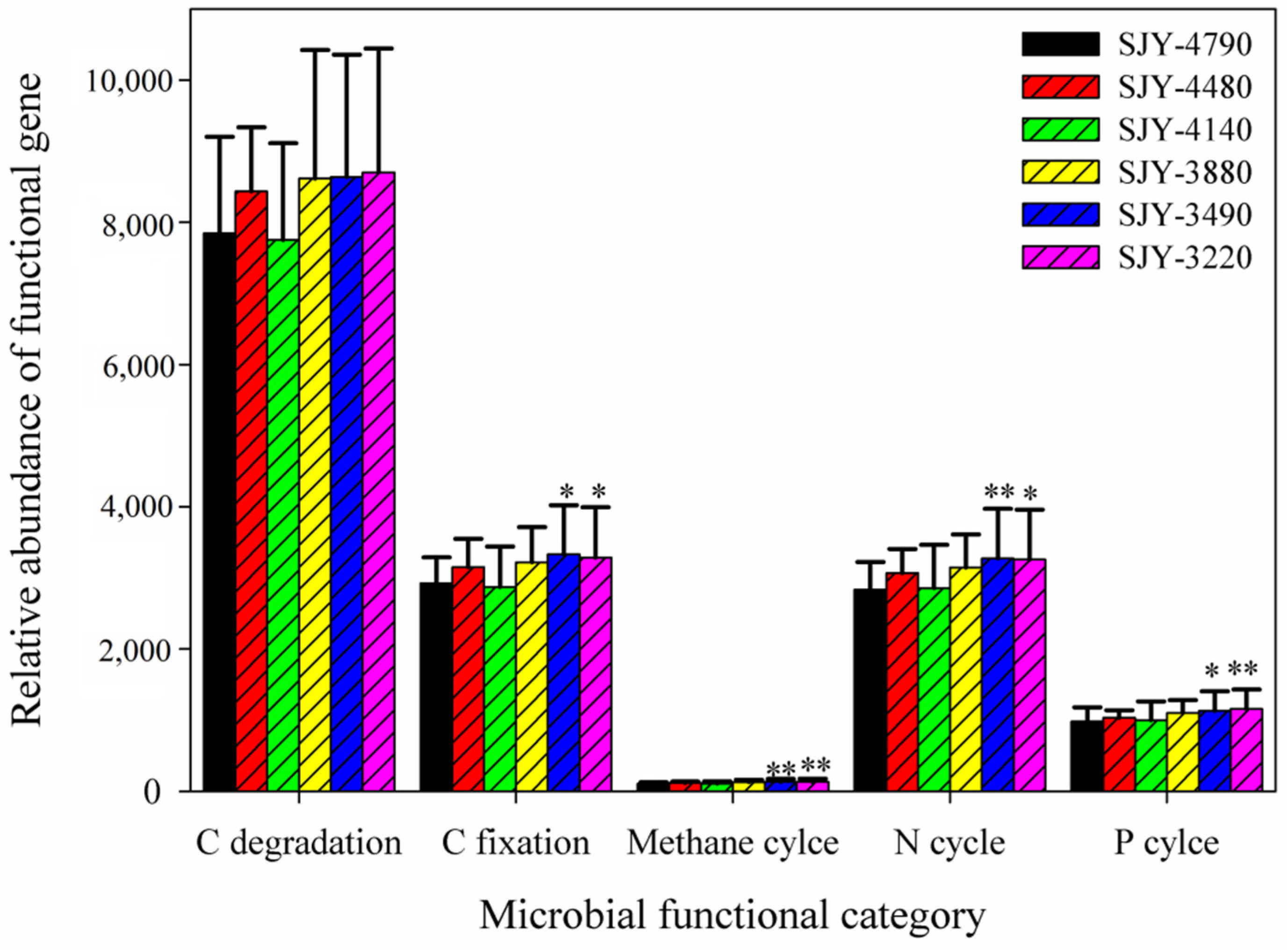
3.4. Soil Microbial Functional Genes Involved in C, N, and P Cycles
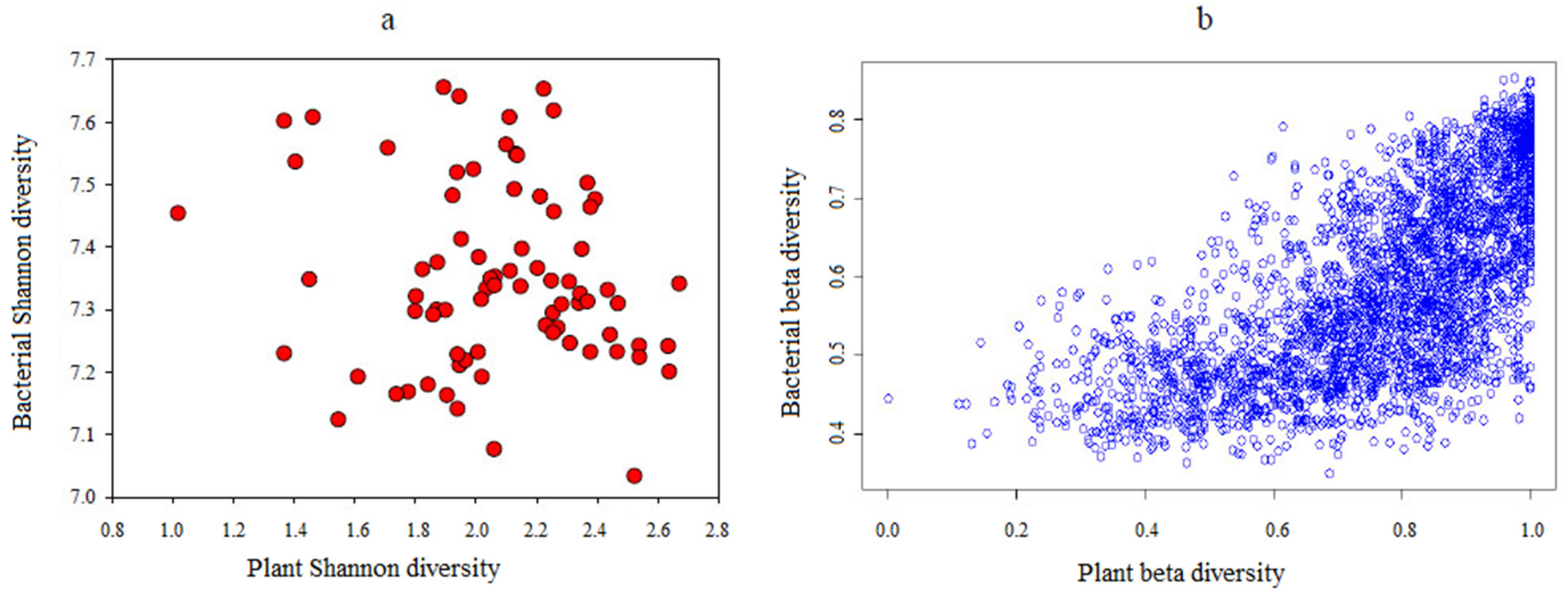
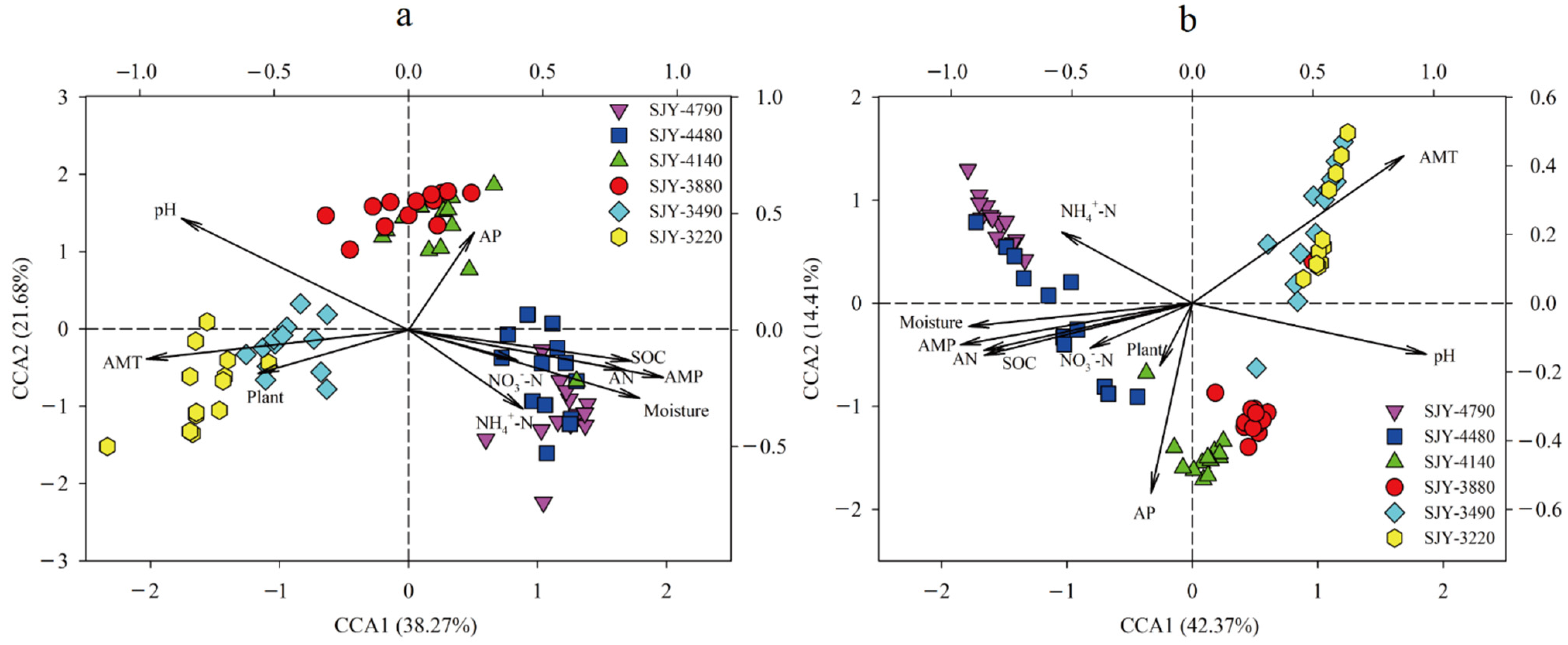
3.5. The Linkages among Plants, Soil Bacterial Diversity, and Environmental Factors
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Singh, B.K.; Bardgett, R.D.; Smith, P.; Reay, D.S. Microorganisms and climate change: Terrestrial feedbacks and mitigation options. Nat. Rev. Microbiol. 2010, 8, 779–790. [Google Scholar] [CrossRef] [PubMed]
- Prober, S.M.; Leff, J.W.; Bates, S.T.; Borer, E.T.; Firn, J.; Harpole, W.S.; Lind, E.M.; Seabloom, E.W.; Adler, P.B.; Bakker, J.D.; et al. Plant diversity predicts beta but not alpha diversity of soil microbes across grasslands worldwide. Ecol. Lett. 2015, 18, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Teste, F.P.; Kardol, P.; Turner, B.L.; Wardle, D.A.; Zemunik, G.; Renton, M.; Laliberté, E. Plant-soil feedback and the maintenance of diversity in Mediterranean-climate shrublands. Science 2017, 355, 173–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Xu, H.; Wu, S.; Baoyin, T. Plant and soil properties mediate the response of soil microbial communities to moderate grazing in a semiarid grassland of northern China. J. Environ. Manag. 2021, 284, 112005. [Google Scholar] [CrossRef] [PubMed]
- Hiiesalu, I.; Pärtel, M.; Davison, J.; Gerhold, P.; Metsis, M.; Moora, M.; Öpik, M.; Vasar, M.; Zobel, M.; Wilson, S.D. Species richness of arbuscular mycorrhizal fungi: Associations with grassland plant richness and biomass. New Phytol. 2014, 203, 233–244. [Google Scholar] [CrossRef]
- Falkowski, P.G.; Fenchel, T.; Delong, E.F. The microbial engines that drive Earth’s biogeochemical cycles. Science 2008, 320, 1034–1039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Classen, A.T.; Sundqvist, M.K.; Henning, J.A.; Newman, G.S.; Moore, J.A.M.; Cregger, M.A.; Moorhead, L.C.; Patterson, C.M. Direct and indirect effects of climate change on soil microbial and soil microbial–plant interactions: What lies ahead? Ecosphere 2016, 6, 1–21. [Google Scholar] [CrossRef]
- Xu, M.; Li, X.; Kuyper, T.W.; Xu, M.; Li, X.; Zhang, J. High microbial diversity stabilizes the responses of soil organic carbon decomposition to warming in the subsoil on the Tibetan Plateau. Glob. Change Biol. 2021, 27, 2061–2075. [Google Scholar] [CrossRef]
- Van der Heijden, M.G.; Bardgett, R.D.; van Straalen, N.M. The unseen majority: Soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecol. Lett. 2008, 11, 296–310. [Google Scholar] [CrossRef]
- Van der Heijden, M.G.; de Bruin, S.; Luckerhoff, L.; van Logtestijn, R.S.; Schlaeppi, K. A widespread plant-fungal-bacterial symbiosis promotes plant biodiversity, plant nutrition and seedling recruitment. ISME J. 2016, 10, 389–399. [Google Scholar] [CrossRef] [Green Version]
- Millard, P.; Singh, B.K. Does grassland vegetation drive soil microbial diversity? Nutr. Cycl. Agroecosyst. 2010, 88, 147–158. [Google Scholar] [CrossRef]
- Gao, C.; Shi, N.N.; Liu, Y.X.; Peay, K.G.; Zheng, Y.; Ding, Q.; Mi, X.C.; Ma, K.P.; Wubet, T.; Buscot, F.; et al. Host plant genus-level diversity is the best predictor of ectomycorrhizal fungal diversity in a Chinese subtropical forest. Mol. Ecol. 2013, 22, 3403–3414. [Google Scholar] [CrossRef] [PubMed]
- Deutsch, C.A.; Tewksbury, J.J.; Huey, R.B.; Sheldon, K.S.; Ghalambor, C.K.; Haak, D.C.; Martin, P.R. Impacts of climate warming on terrestrial ectotherms across latitude. Proc. Natl. Acad. Sci. USA 2008, 105, 6668–6672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Y.; Si, G.; Wang, J.; Luo, T.; Zhang, G. Bacterial community in alpine grasslands along an altitudinal gradient on the Tibetan Plateau. FEMS Microbiol. Ecol. 2014, 87, 121–132. [Google Scholar] [CrossRef] [Green Version]
- Yashiro, E.; Pinto-Figueroa, E.; Buri, A.; Spangenberg, J.E.; Adatte, T.; Niculita-Hirzel, H.; Guisan, A.; van der Meer, J.R. Local Environmental Factors Drive Divergent Grassland Soil Bacterial Communities in the Western Swiss Alps. Appl. Environ. Microbiol. 2016, 82, 6303–6316. [Google Scholar] [CrossRef] [Green Version]
- Yashiro, E.; Pinto-Figueroa, E.; Buri, A.; Spangenberg, J.E.; Adatte, T.; Niculita-Hirzel, H.; Guisan, A.; van der Meer, J.R. Meta-scale mountain grassland observatories uncover commonalities as well as specific interactions among plant and non-rhizosphere soil bacterial communities. Sci. Rep. 2018, 8, 5758. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Freeman, C.; Ostle, N.J. Microbial contributions to climate change through carbon cycle feedbacks. ISME J. 2008, 2, 805–814. [Google Scholar] [CrossRef] [Green Version]
- Dieleman, W.I.; Vicca, S.; Dijkstra, F.A.; Hagedorn, F.; Hovenden, M.J.; Larsen, K.S.; Morgan, J.A.; Volder, A.; Beier, C.; Dukes, J.S.; et al. Simple additive effects are rare: A quantitative review of plant biomass and soil process responses to combined manipulations of CO2 and temperature. Glob. Change Biol. 2012, 18, 2681–2693. [Google Scholar] [CrossRef]
- Nie, M.; Pendall, E.; Bell, C.; Gasch, C.K.; Raut, S.; Tamang, S.; Wallenstein, M.D. Positive climate feedbacks of soil microbial communities in a semi-arid grassland. Ecol. Lett. 2013, 16, 234–241. [Google Scholar] [CrossRef]
- Melillo, J.M.; Butler, S.; Johnson, J.; Mohan, J.; Steudler, P.; Lux, H.; Burrows, E.; Bowles, F.; Smith, R.; Scott, L.; et al. Soil warming, carbon-nitrogen interactions, and forest carbon budgets. Proc. Natl. Acad. Sci. USA 2011, 108, 9508–9512. [Google Scholar] [CrossRef] [Green Version]
- Margesin, R.; Minerbi, S.; Schinner, F. Long-term monitoring of soil microbiological activities in two forest sites in South Tyrol in the Italian alps. Microbes Environ. 2014, 29, 277–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero-Olivares, A.L.; Allison, S.D.; Treseder, K.K. Soil microbes and their response to experimental warming over time: A meta-analysis of field studies. Soil Biol. Biochem. 2017, 107, 32–40. [Google Scholar] [CrossRef] [Green Version]
- Bryant, J.A.; Lamanna, C.; Morlon, H.; Kerkhoff, A.J.; Enquist, B.J.; Green, J.L. Colloquium paper: Microbes on mountainsides: Contrasting elevational patterns of bacterial and plant diversity. Proc. Natl. Acad. Sci. USA 2008, 105 (Suppl. S1), 11505–11511. [Google Scholar] [CrossRef] [Green Version]
- Nottingham, A.T.; Whitaker, J.; Turner, B.L.; Salinas, N.; Zimmermann, M.; Malhi, Y.; Meir, P. Climate Warming and Soil Carbon in Tropical Forests: Insights from an Elevation Gradient in the Peruvian Andes. Bioscience 2015, 65, 906–921. [Google Scholar] [CrossRef]
- Colwell, R.K.; Brehm, G.; Cardelús, C.L.; Gilman, A.C.; Longino, J.T. Global warming, elevational range shifts, and lowland biotic attrition in the wet tropics. Science 2008, 322, 258–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malhi, Y.; Silman, M.; Salinas, N.; Bush, M.; Meir, P.; Saatchi, S. Introduction: Elevation gradients in the tropics: Laboratories for ecosystem ecology and global change research. Glob. Change Biol. 2010, 16, 3171–3175. [Google Scholar] [CrossRef]
- Wang, S.; Duan, J.; Xu, G.; Wang, Y.; Zhang, Z.; Rui, Y.; Luo, C.; Xu, B.; Zhu, X.; Chang, X.; et al. Effects of warming and grazing on soil N availability, species composition, and ANPP in an alpine meadow. Ecology 2012, 93, 2365–2376. [Google Scholar] [CrossRef]
- Qiu, J. China: The third pole. Nature 2008, 454, 393–396. [Google Scholar] [CrossRef] [Green Version]
- Kang, L.; Han, X.; Zhang, Z.; Sun, O.J. Grassland ecosystems in China: Review of current knowledge and research advancement. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2007, 362, 997–1008. [Google Scholar] [CrossRef]
- Zhou, H.; Zhao, X.; Tang, Y.; Song, G.; Li, Z. Alpine grassland degradation and its control in the source region of the Yangtze and Yellow Rivers, China. Grassl. Sci. 2005, 51, 191–203. [Google Scholar] [CrossRef]
- Genxu, W.; Ju, Q.; Guodong, C.; Yuanmin, L. Soil organic carbon pool of grassland soils on the Qinghai-Tibetan Plateau and its global implication. Sci. Total Environ. 2002, 291, 207–217. [Google Scholar] [CrossRef]
- Dunne, J.A.; Saleska, S.R.; Fischer, M.L.; Harte, J. Integrating experimental and gradient methods in ecological climate change research. Ecology 2004, 85, 904–916. [Google Scholar] [CrossRef]
- Blois, J.L.; Williams, J.W.; Fitzpatrick, M.C.; Jackson, S.T.; Ferrier, S. Space can substitute for time in predicting climate-change effects on biodiversity. Proc. Natl. Acad. Sci. USA 2013, 110, 9374–9379. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Zhu, Q.; Peng, C.; Wu, N.; Wang, Y.; Fang, X.; Gao, Y.; Zhu, D.; Yang, G.; Tian, J.; et al. The impacts of climate change and human activities on biogeochemical cycles on the Qinghai-Tibetan Plateau. Glob. Change Biol. 2013, 19, 2940–2955. [Google Scholar] [CrossRef]
- Zhang, Y.; Lu, Z.; Liu, S.; Yang, Y.; He, Z.; Ren, Z.; Zhou, J.; Li, D. Geochip-based analysis of microbial communities in alpine meadow soils in the Qinghai-Tibetan plateau. BMC Microbiol. 2013, 13, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hui, L.U.; Cong, J.; Liu, X.; Wang, X.L.; Tang, J.; Di-Qiang, L.I.; Zhang, Y.G. Plant diversity patterns along altitudinal gradients in alpine meadows in the Three River Headwater Region, China. Acta Pratacult. Sin. 2015, 24, 197–204. [Google Scholar]
- Semenov, M.V.; Nikitin, D.A.; Stepanov, A.L.; Semenov, V.M. The Structure of Bacterial and Fungal Communities in the Rhizosphere and Root-Free Loci of Gray Forest Soil. Eurasian Soil Sci. 2019, 52, 319–332. [Google Scholar] [CrossRef]
- Cong, J.; Yang, Y.; Liu, X.; Lu, H.; Liu, X.; Zhou, J.; Li, D.; Yin, H.; Ding, J.; Zhang, Y. Analyses of soil microbial community compositions and functional genes reveal potential consequences of natural forest succession. Sci. Rep. 2015, 5, 10007. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. S1), 4516–4522. [Google Scholar] [CrossRef] [Green Version]
- Ding, J.; Zhang, Y.; Deng, Y.; Cong, J.; Lu, H.; Sun, X.; Yang, C.; Yuan, T.; Van Nostrand, J.D.; Li, D.; et al. Integrated metagenomics and network analysis of soil microbial community of the forest timberline. Sci. Rep. 2015, 5, 7994. [Google Scholar] [CrossRef] [Green Version]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tu, Q.; Yu, H.; He, Z.; Deng, Y.; Wu, L.; Van Nostrand, J.D.; Zhou, A.; Voordeckers, J.; Lee, Y.J.; Qin, Y.; et al. GeoChip 4: A functional gene-array-based high-throughput environmental technology for microbial community analysis. Mol. Ecol. Resour. 2014, 14, 914–928. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Lian, J.; Wang, L.; Wang, X.; Luo, Y.; Wang, W.; Wu, F.; Zhao, J.; Ding, Y.; Ma, J.; et al. Variation in Soil Microbial Communities Along an Elevational Gradient in Alpine Meadows of the Qilian Mountains, China. Front. Microbiol. 2021, 12, 684386. [Google Scholar] [CrossRef]
- Shen, C.; Xiong, J.; Zhang, H.; Feng, Y.; Lin, X.; Li, X.; Liang, W.; Chu, H. Soil pH drives the spatial distribution of bacterial communities along elevation on Changbai Mountain. Soil Biol. Biochem. 2013, 57, 204–211. [Google Scholar] [CrossRef]
- Guo, Y.; Ren, C.; Yi, J.; Doughty, R.; Zhao, F. Contrasting Responses of Rhizosphere Bacteria, Fungi and Arbuscular Mycorrhizal Fungi Along an Elevational Gradient in a Temperate Montane Forest of China. Front. Microbiol. 2020, 11, 2042. [Google Scholar] [CrossRef] [PubMed]
- Putten, V.D.; Wim, H. Climate Change, Aboveground-Belowground Interactions, and Species’ Range Shifts. Annu. Rev. Ecol. Evol. Syst. 2012, 43, 365–383. [Google Scholar] [CrossRef]
- Zhang, Y.; Dong, S.; Gao, Q.; Liu, S.; Zhou, H.; Ganjurjav, H.; Wang, X. Climate change and human activities altered the diversity and composition of soil microbial community in alpine grasslands of the Qinghai-Tibetan Plateau. Sci. Total Environ. 2016, 562, 353–363. [Google Scholar] [CrossRef]
- Salas-Morales, S.H.; Meave, J.A. Elevational patterns in the vascular flora of a highly diverse region in southern Mexico. Plant Ecol. 2012, 213, 1209–1220. [Google Scholar] [CrossRef]
- Nyman, T.; Linder, H.P.; Peña, C.; Malm, T.; Wahlberg, N. Climate-driven diversity dynamics in plants and plant-feeding insects. Ecol. Lett. 2012, 15, 889–898. [Google Scholar] [CrossRef]
- Liu, S.; Kruse, S.; Scherler, D.; Ree, R.H.; Zimmermann, H.H.; Stoof-Leichsenring, K.R.; Epp, L.S.; Mischke, S.; Herzschuh, U. Sedimentary ancient DNA reveals a threat of warming-induced alpine habitat loss to Tibetan Plateau plant diversity. Nat. Commun. 2021, 12, 2995. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Liu, C.; Wang, W.; Zhou, H.; Xue, Y.; Xu, J.; Xue, P.; Yan, H. Effects of Different Grazing Disturbances on the Plant Diversity and Ecological Functions of Alpine Grassland Ecosystem on the Qinghai-Tibetan Plateau. Front. Plant Sci. 2021, 12, 765070. [Google Scholar] [CrossRef]
- Meadow, J.F.; Zabinski, C.A. Linking symbiont community structures in a model arbuscular mycorrhizal system. New Phytol. 2012, 194, 800–809. [Google Scholar] [CrossRef] [PubMed]
- Milcu, A.; Allan, E.; Roscher, C.; Jenkins, T.; Meyer, S.T.; Flynn, D.; Bessler, H.; Buscot, F.; Engels, C.; Gubsch, M.; et al. Functionally and phylogenetically diverse plant communities key to soil biota. Ecology 2013, 94, 1878–1885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhelezova, A.; Chernov, T.; Nikitin, D.; Tkhakakhova, A.; Ksenofontova, N.; Zverev, A.; Kutovaya, O.; Semenov, M. Seasonal Dynamics of Soil Bacterial Community under Long-Term Abandoned Cropland in Boreal Climate. Agronomy 2022, 12, 519. [Google Scholar] [CrossRef]
- Hawkes, C.V.; Keitt, T.H. Resilience vs. historical contingency in microbial responses to environmental change. Ecol. Lett. 2015, 18, 612–625. [Google Scholar] [CrossRef]
- Zhou, J.Z.; Kai, X.; Xie, J.P.; Ye, D.; Luo, Y.Q. Microbial mediation of carbon-cycle feedbacks to climate warming. Nat. Clim. Change 2011, 2, 106–110. [Google Scholar] [CrossRef]
- Cheng, Y.T.; Zhang, L.; He, S.Y. Plant-Microbe Interactions Facing Environmental Challenge. Cell Host Microbe 2019, 26, 183–192. [Google Scholar] [CrossRef]
- Berg, M.P.; Kiers, E.T.; Driessen, G.; Van Der Heijden, M.; Kooi, B.W.; Kuenen, F.; Liefting, M.; Verhoef, H.A.; Ellers, J. Adapt or disperse: Understanding species persistence in a changing world. Glob. Change Biol. 2020, 16, 587–598. [Google Scholar] [CrossRef]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an ecological classification of soil bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar] [CrossRef]
- Djukic, I.; Zehetner, F.; Watzinger, A.; Horacek, M.; Gerzabek, M.H. In situ carbon turnover dynamics and the role of soil microorganisms therein: A climate warming study in an Alpine ecosystem. FEMS Microbiol. Ecol. 2013, 83, 112–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Lv, W.; Jiang, L.; Zhang, L.; Wang, S.; Wang, Q.; Xue, K.; Li, B.; Liu, P.; Hong, H.; et al. Microbial community responses reduce soil carbon loss in Tibetan alpine grasslands under short-term warming. Glob. Change Biol. 2019, 25, 3438–3449. [Google Scholar] [CrossRef] [PubMed]
- Rousk, J.; Smith, A.R.; Jones, D.L. Investigating the long-term legacy of drought and warming on the soil microbial community across five European shrubland ecosystems. Glob. Change Biol. 2013, 19, 3872–3884. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.; Shi, Y.; Fan, K.; He, J.S.; Adams, J.M.; Ge, Y.; Chu, H. Soil pH dominates elevational diversity pattern for bacteria in high elevation alkaline soils on the Tibetan Plateau. FEMS Microbiol. Ecol. 2019, 95, fiz003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freeman, C.; Nevison, G.B.; Kang, H.; Hughes, S.; Reynolds, B.; Hudson, J.A. Contrasted effects of simulated drought on the production and oxidation of methane in a mid-Wales wetland. Soil Biol. Biochem. 2002, 34, 61–67. [Google Scholar] [CrossRef]
- Chen, Q.L.; Ding, J.; Zhu, Y.G.; He, J.Z.; Hu, H.W. Soil bacterial taxonomic diversity is critical to maintaining the plant productivity. Environ. Int. 2020, 140, 105766. [Google Scholar] [CrossRef]
- Ke, P.J.; Miki, T.; Ding, T.S. The soil microbial community predicts the importance of plant traits in plant-soil feedback. New Phytol. 2015, 206, 329–341. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, D.A.; da Silva, T.F.; Pylro, V.S.; Salles, J.F.; Andreote, F.D.; Dini-Andreote, F. Soil Microbial Diversity Affects the Plant-Root Colonization by Arbuscular Mycorrhizal Fungi. Microb. Ecol. 2021, 82, 100–103. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Maestre, F.T.; Reich, P.B.; Jeffries, T.C.; Gaitan, J.J.; Encinar, D.; Berdugo, M.; Campbell, C.D.; Singh, B.K. Microbial diversity drives multifunctionality in terrestrial ecosystems. Nat. Commun. 2016, 7, 10541. [Google Scholar] [CrossRef] [Green Version]
- García-Palacios, P.; Vandegehuchte, M.L.; Shaw, E.A.; Dam, M.; Post, K.H.; Ramirez, K.S.; Sylvain, Z.A.; de Tomasel, C.M.; Wall, D.H. Are there links between responses of soil microbes and ecosystem functioning to elevated CO2, N deposition and warming? A global perspective. Glob. Change Biol. 2015, 21, 1590–1600. [Google Scholar] [CrossRef]
- Nikitin, D.A.; Semenov, M.V.; Chernov, T.I.; Ksenofontova, N.A.; Zhelezova, A.D.; Ivanova, E.A.; Khitrov, N.B.; Stepanov, A.L. Microbiological Indicators of Soil Ecological Functions: A Review. Eurasian Soil Sci. 2022, 55, 221–234. [Google Scholar] [CrossRef]
- Li, Z.; Zeng, Z.; Song, Z.; Wang, F.; Tian, D.; Mi, W.; Huang, X.; Wang, J.; Song, L.; Yang, Z.; et al. Vital roles of soil microbes in driving terrestrial nitrogen immobilization. Glob. Change Biol. 2021, 27, 1848–1858. [Google Scholar] [CrossRef] [PubMed]
- Mehnaz, K.R.; Keitel, C.; Dijkstra, F.A. Phosphorus availability and plants alter soil nitrogen retention and loss. Sci. Total Environ. 2019, 671, 786–794. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Wang, P. Afforestation suppresses soil nitrogen availability and soil multifunctionality on a subtropical grassland. Sci. Total Environ. 2021, 761, 143663. [Google Scholar] [CrossRef]
- Zhang, C.; Dong, W.; Manevski, K.; Hu, W.; Timilsina, A.; Chen, X.; Zhang, X.; Hu, C. Long-term warming and nitrogen fertilization affect C-, N- and P-acquiring hydrolase and oxidase activities in winter wheat monocropping soil. Sci. Rep. 2021, 11, 18542. [Google Scholar] [CrossRef]
- DeForest, J.L.; Smemo, K.A.; Burke, D.J.; Elliott, H.L.; Becker, J.C. Soil microbial responses to elevated phosphorus and pH in acidic temperate deciduous forests. Biogeochemistry 2012, 109, 189–202. [Google Scholar] [CrossRef]
- Zhang, B.; Xue, K.; Zhou, S.; Che, R.; Du, J.; Tang, L.; Pang, Z.; Wang, F.; Wang, D.; Cui, X.; et al. Phosphorus mediates soil prokaryote distribution pattern along a small-scale elevation gradient in Noijin Kangsang Peak, Tibetan Plateau. FEMS Microbiol. Ecol. 2019, 95, fiz076. [Google Scholar] [CrossRef]
- Luo, G.; Xue, C.; Jiang, Q.; Xiao, Y.; Zhang, F.; Guo, S.; Shen, Q.; Ling, N. Soil Carbon, Nitrogen, and Phosphorus Cycling Microbial Populations and Their Resistance to Global Change Depend on Soil C:N:P Stoichiometry. mSystems 2020, 5, e00162-20. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site Name | Location | Elevation | Latitude, Longitude | Slope | Aspect | Dominant Plant Species | MAT of the Warmest Quarter (°C) | MAP (mm) |
|---|---|---|---|---|---|---|---|---|
| SJY-4790 | Maduo County, Qinghai Province | 4790 m | 34°08′16″ N 97°40′22″ E | 15° | N | Kobresia tibetica, Kobresia pygmaea | 4.90 | 417.00 |
| SJY-4480 | Maduo County, Qinghai Province | 4480 m | 34°22′15″ N 97°56′57″ E | 18° | 30°, NW | Kobresia tibetica | 6.20 | 386.00 |
| SJY-4140 | Maduo County, Qinghai Province | 4140 m | 35°24′28″ N 99°21′6″ E | 5° | 20°, NW | Kobresia pygmaea, Kobresia humilis | 6.30 | 372.00 |
| SJY-3880 | Xinghai County, Qinghai Province | 3880 m | 35°41′26″ N 99°33′1″ E | 15° | 30°, NW | Kobresia pygmaea, Stipa capillata | 7.40 | 354.00 |
| SJY-3490 | Xinghai County, Qinghai Province | 3490 m | 35°40′10″ N 99°55′13″ E | 5° | N | Poa annua, Stipa capillata | 10.00 | 344.00 |
| SJY-3220 | Xinghai County, Qinghai Province | 3220 m | 35°56′6″ N 100°5′27″ E | 5° | N | Elymus nutans, Stipa capillata | 11.60 | 319.00 |
| Site Name | Plant Shannon Index | Plant Pielou Index | Bacterial Shannon Index | Bacterial Simpson Index |
|---|---|---|---|---|
| SJY-4790 | 1.97 ± 0.26 c | 0.79 ± 0.09 ab | 7.27 ± 0.10 a | 448 ± 102 a |
| SJY-4480 | 2.34 ± 0.23 a | 0.84 ± 0.07 a | 7.26 ± 0.12 a | 483 ± 109 a |
| SJY-4140 | 2.27 ± 0.25 ab | 0.85 ± 0.08 a | 7.35 ± 0.09 a | 518 ± 112 a |
| SJY-3880 | 1.72 ± 0.30 d | 0.74 ± 0.10 b | 7.27 ± 0.12 a | 476 ± 87 a |
| SJY-3490 | 2.09 ± 0.14b | 0.85 ± 0.04a | 7.50 ± 0.08b | 755 ± 145b |
| SJY-3220 | 2.05 ± 0.34 bc | 0.86 ± 0.08 a | 7.45 ± 0.16 b | 702 ± 185 b |
| Environmental Factors | Plant Beta Diversity | Bacterial Beta Diversity | ||
|---|---|---|---|---|
| r | p | r | p | |
| Site elevation | 0.496 | <0.001 | 0.670 | <0.001 |
| Mean annual temperature | 0.410 | 0.001 | 0.437 | 0.001 |
| Mean annual precipitation | 0.373 | 0.001 | 0.624 | 0.001 |
| Soil moisture | 0.164 | 0.001 | 0.452 | 0.001 |
| Soil pH | 0.255 | 0.001 | 0.634 | 0.001 |
| Soil organic carbon | 0.036 | 0.107 | 0.165 | 0.001 |
| Available nitrogen | −0.017 | 0.735 | 0.094 | 0.008 |
| Available phosphorus | 0.089 | 0.010 | 0.030 | 0.268 |
| Soil NH4+- N | −0.103 | 0.999 | 0.150 | 0.003 |
| Soil NO3−- N | −0.039 | 0.864 | −0.040 | 0.778 |
| Bacteria Shannon index | 0.176 | 0.004 | - | - |
| Plant Shannon index | - | - | 0.060 | 0.069 |
| Bacterial beta diversity | 0.493 | 0.001 | - | |
| Plant beta diversity | - | - | 0.493 | 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cong, J.; Cong, W.; Lu, H.; Zhang, Y. Distinct Elevational Patterns and Their Linkages of Soil Bacteria and Plant Community in An Alpine Meadow of the Qinghai–Tibetan Plateau. Microorganisms 2022, 10, 1049. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10051049
Cong J, Cong W, Lu H, Zhang Y. Distinct Elevational Patterns and Their Linkages of Soil Bacteria and Plant Community in An Alpine Meadow of the Qinghai–Tibetan Plateau. Microorganisms. 2022; 10(5):1049. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10051049
Chicago/Turabian StyleCong, Jing, Wei Cong, Hui Lu, and Yuguang Zhang. 2022. "Distinct Elevational Patterns and Their Linkages of Soil Bacteria and Plant Community in An Alpine Meadow of the Qinghai–Tibetan Plateau" Microorganisms 10, no. 5: 1049. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10051049