
Effects of Humic Substances on the Growth of Pseudomonas plecoglossicida 2,4-D and Wheat Plants Inoculated with This Strain

Abstract
:1. Introduction
2. Materials and Methods
2.1. Extraction of Humic and Fulvic Acids and Their Spectral Characteristics
2.2. Bacterial Strain and Culture Media
2.3. Estimation of Root Colonization
2.4. Experiments with Plants
3. Results
3.1. Spectral Characteristics of Humic and Fulvic Fractions
3.2. Effects of Humic Substances on Proliferation of Bacteria In Vitro
3.3. Root Colonization by Bacteria
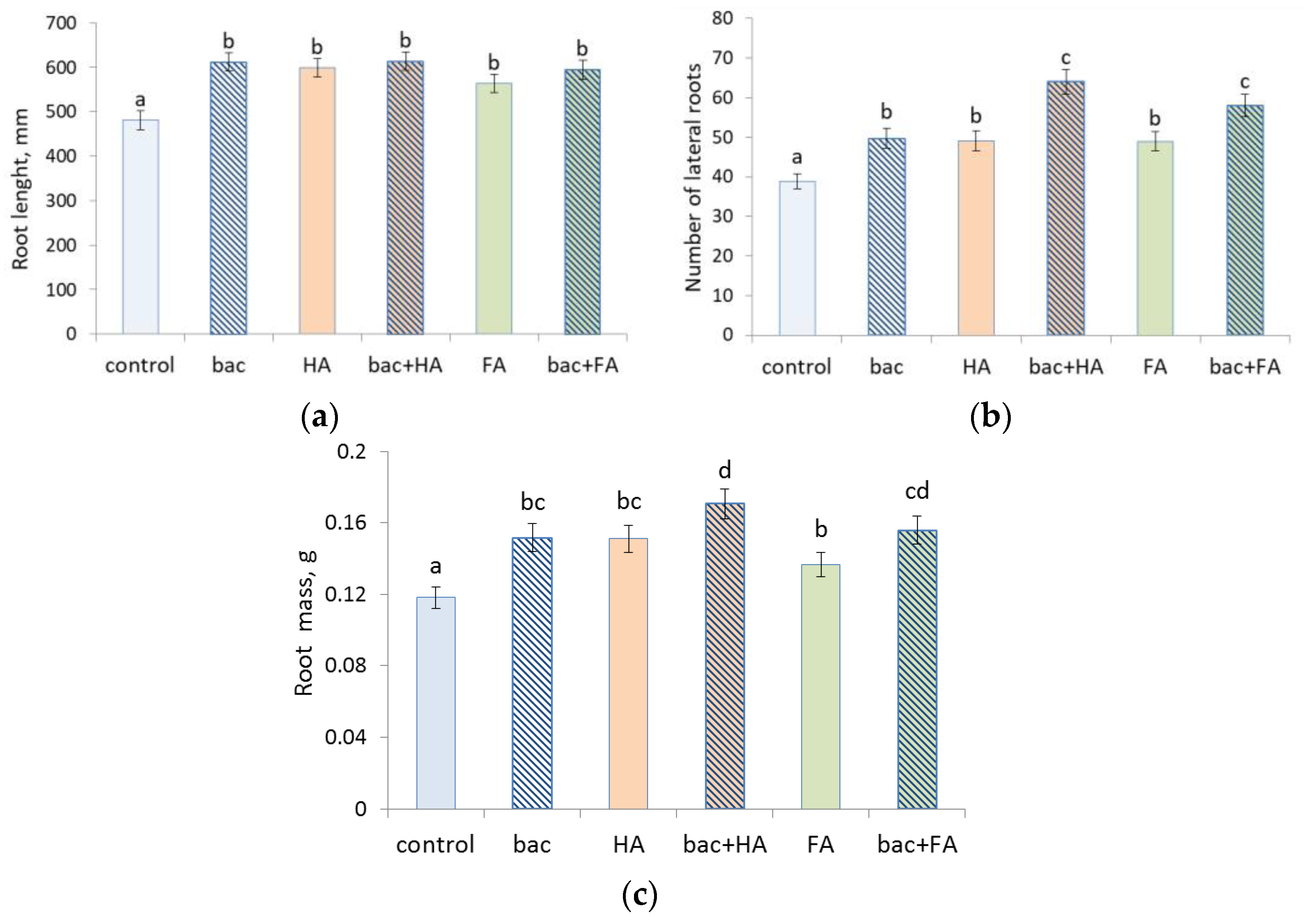
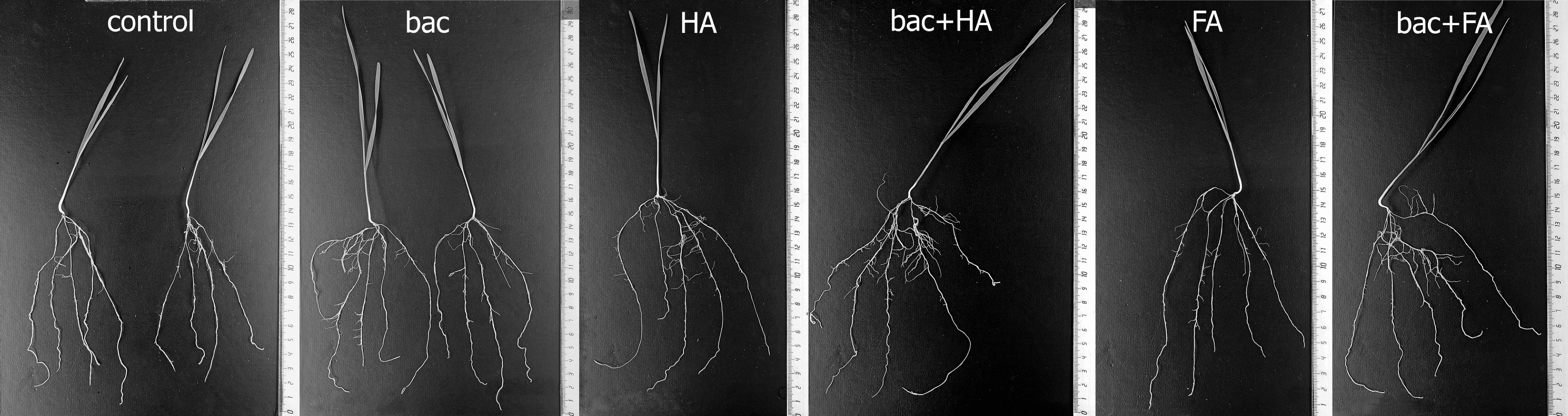
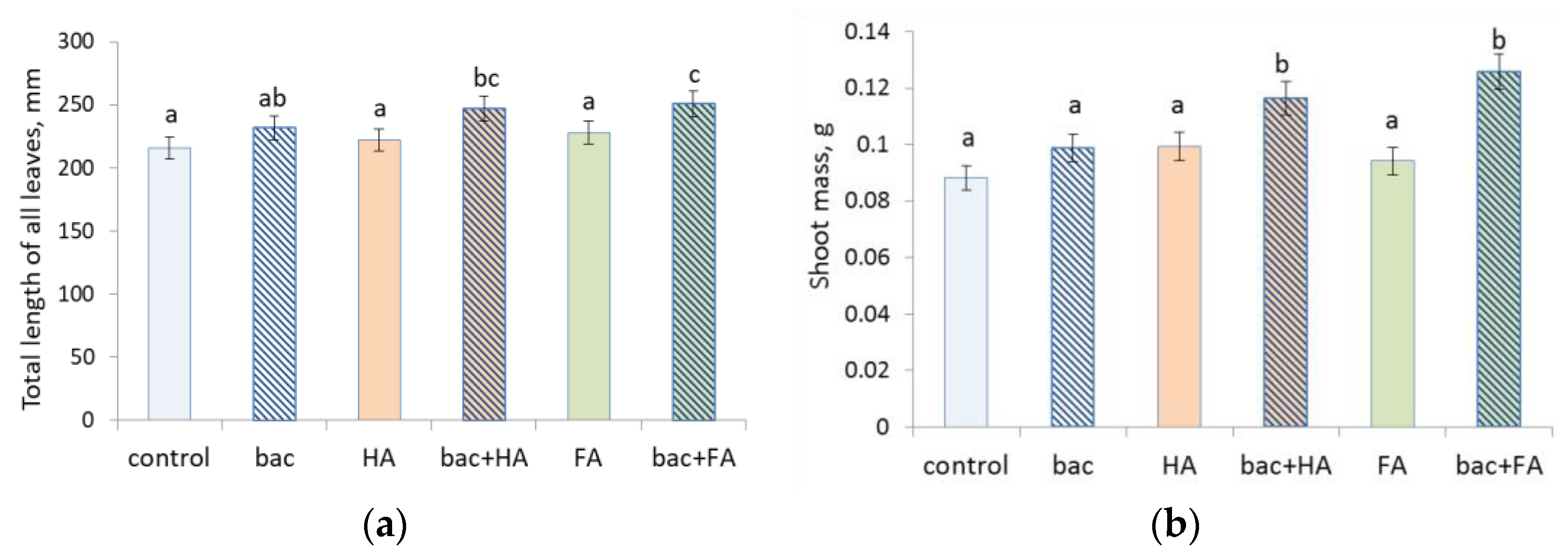
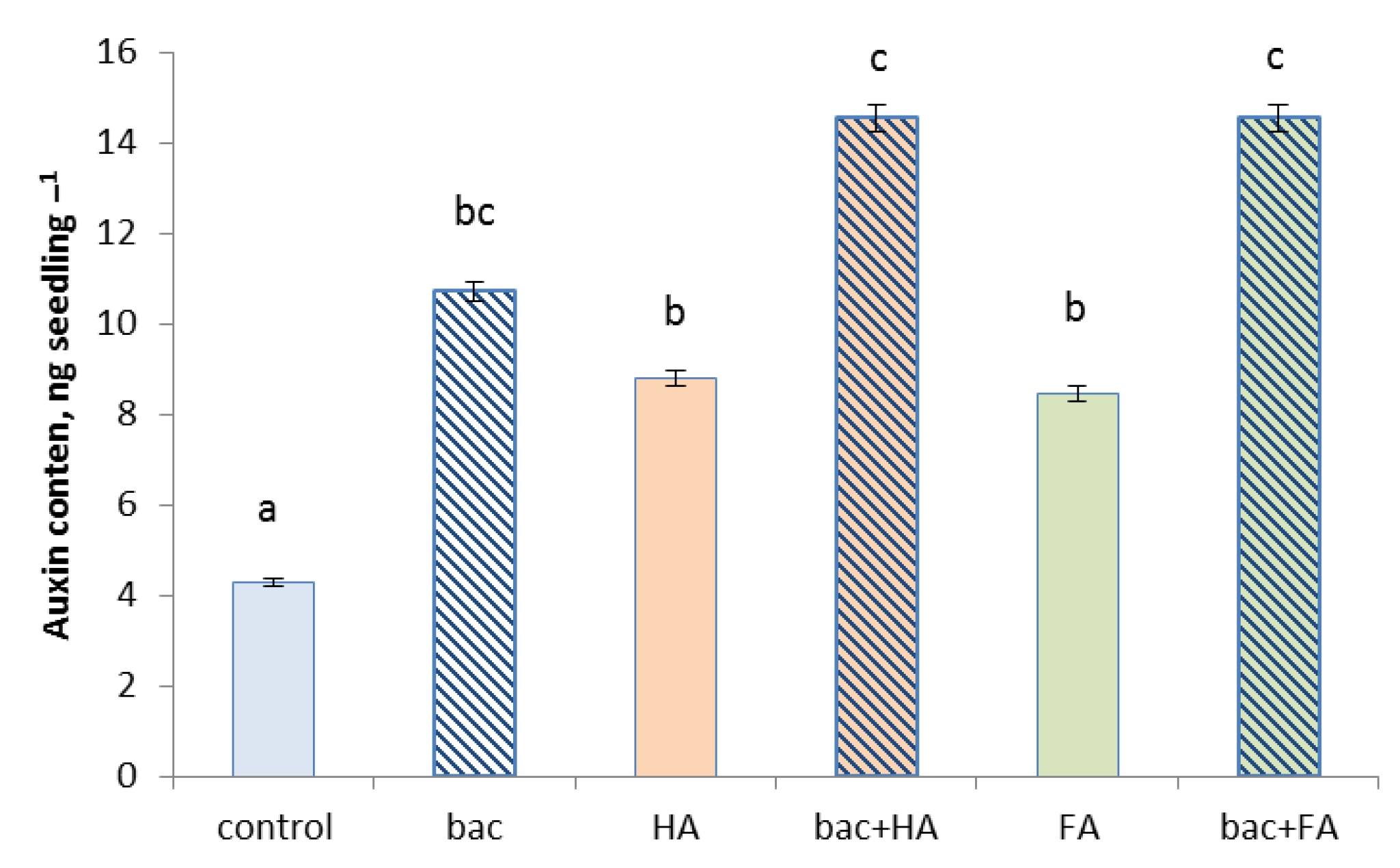
3.4. Treatment Effects on Plants
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time of Incubation, Days | Additives to the Medium | |||
|---|---|---|---|---|
| No Additives (Control) | HA | FA | HSs | |
| 0 | (3.91 ± 0.48) × 105 a | (4.15 ± 0.42) × 105 a | (4.03 ± 0.37) × 105 a | (4.17 ± 0.33) × 105 a |
| 1 | (1.42 ± 0.22) × 107 a | (3.02 ± 0.51) × 107 a | (1.81 ± 0.28) × 107 a | (2.50 ± 0.40) × 107 a |
| 2 | (1.03 ± 0.18) × 107 a | (1.10 ± 0.23) × 107 a | (8.36 ± 0.92) × 106 a | (1.14 ± 0.19) × 107 a |
| 7 | (5.81 ± 0.55) × 106 a | (8.05 ± 0.72) × 106 a | (5.07 ± 0.66) × 106 a | (6.10 ± 0.69) × 106 a |
References
- Ruzzi, M.; Aroca, R. Plant growth-promoting rhizobacteria act as biostimulants in horticulture. Sci. Hortic. 2015, 196, 124–134. [Google Scholar] [CrossRef]
- Backer, R.; Rokem, J.S.; Ilangumaran, G.; Lamont, J.; Praslickova, D.; Ricci, E.; Subramanian, S.; Smith, D.L. Plant Growth-Promoting Rhizobacteria: Context, Mechanisms of Action, and Roadmap to Commercialization of Biostimulants for Sustainable Agriculture. Front. Plant Sci. 2018, 9, 1473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kudoyarova, G.; Arkhipova, T.; Korshunova, T.; Bakaeva, M.; Loginov, O.; Dodd, I.C. Phytohormone mediation of interactions between plants and non-symbiotic growth promoting bacteria under edaphic stresses. Front. Plant Sci. 2019, 10, 1368. [Google Scholar] [CrossRef] [PubMed]
- Canellas, L.P.; Olivares, F.L.; Aguiar, N.O.; Jones, D.L.; Nebbioso, A.; Mazzei, P. Humic and fulvic acids as biostimulants in horticulture. Sci. Hortic. 2015, 196, 15–27. [Google Scholar] [CrossRef]
- Olaetxea, M.; De Hita, D.; Garcia, C.A.; Fuentes, M.; Baigorri, R.; Mora, V. Hypothetical framework integrating the main mechanisms involved in the promoting action of rhizospheric humic substances on plant root and shoot-growth. Appl. Soil Ecol. 2018, 123, 521–537. [Google Scholar] [CrossRef]
- Nazarov, A.M.; Garankov, I.N.; Tuktarova, I.O.; Salmanova, E.R.; Arkhipova, T.N.; Ivanov, I.I.; Feoktistova, A.V.; Prostyakova, Z.G.; Kudoyarova, G.R. Hormone balance and shoot growth in wheat (Triticum durum Desf.) plants as influenced by sodium humates of the granulated organic fertilizer. Sel’skokhozyaistvennaya Biol. 2020, 55, 945–955. [Google Scholar] [CrossRef]
- Lopes, M.J.S.; Dias-Filho, M.B.; Gurgel, E.S.C. Successful plant growth-promoting microbes: Inoculation methods and abiotic factors. Front. Sustain. Food Syst. 2021, 5, 606454. [Google Scholar] [CrossRef]
- Bakhshandeh, E.; Gholamhosseini, M.; Yaghoubian, Y.; Pirdashti, H. Plant growth promoting microorganisms can improve germination, seedling growth and potassium uptake of soybean under drought and salt stress. Plant Growth Regul. 2020, 90, 123–136. [Google Scholar] [CrossRef]
- Czarnes, S.; Mercier, P.-E.; Lemoine, D.G.; Hamzaoui, J.; Legendre, L. Impact of soil water content on maize responses to the plant growth-promoting rhizobacterium Azospirillum lipoferum CRT1. J. Agro. Crop Sci. 2020, 206, 505–516. [Google Scholar] [CrossRef]
- Mukhtar, T.; Rehman, S.; Smith, D.; Sultan, T.; Seleiman, M.F.; Alsadon, A.A. Mitigation of heat stress in Solanum lycopersicum L. by ACC-deaminase and exopolysaccharide producing Bacillus cereus: Effects on biochemical profiling. Sustainability 2020, 12, 2159. [Google Scholar] [CrossRef] [Green Version]
- El-Sayed, S.Y.S.; Hagab, R.H. Effect of organic acids and plant growth promoting rhizobacteria (PGPR) on biochemical content and productivity of wheat under saline soil conditions. Middle East J. Agric. Res. 2020, 9, 227–242. [Google Scholar] [CrossRef]
- Ullah, A.; Ali, M.; Shahzad, K.; Ahmad, F.; Iqbal, S.; Rahman, M.H.U.; Ahmad, S.; Iqbal, M.M.; Danish, S.; Fahad, S.; et al. Impact of seed dressing and soil application of potassium humate on cotton plants productivity and fiber quality. Plants 2020, 9, 1444. [Google Scholar] [CrossRef] [PubMed]
- Olaetxea, M.; Mora, V.; Bacaicoa, E.; Garnica, M.; Fuentes, M.; Casanova, E.; Zamarreño, A.M.; Iriarte, J.C.; Etayo, D.; Ederra, I. Abscisic acid regulation of root hydraulic conductivity and aquaporin gene expression is crucial to the plant shoot growth enhancement caused by rhizosphere humic acids. Plant Physiol. 2015, 169, 2587–2596. [Google Scholar] [CrossRef] [PubMed]
- Canellas, L.P.; Olivares, F.L.; Okorokova-Façanha, A.L.; Façanha, A.R. Humic acids isolated from earthworm compost enhance root elongation, lateral root emergence, and plasma membrane H+-ATPase activity in maize roots. Plant Physiol. 2002, 130, 1951–1957. [Google Scholar] [CrossRef] [Green Version]
- de Andrade da Silva, M.S.R.; de Melo Silveira Dos Santos, B.; de Andrade da Silva, C.S.R.; de Andrade da Silva, C.S.R.; de Sousa Antunes, L.F.; Dos Santos, R.M.; Santos, C.H.B.; Rigobelo, E.C. Humic substances in combination with plant growth-promoting bacteria as an alternative for sustainable agriculture. Front. Microbiol. 2021, 12, 719653. [Google Scholar] [CrossRef]
- Olivares, F.L.; Busato, J.G.; de Paula, A.M.; da Silva Lima, L.; Aguiar, N.O.; Canellas, L.P. Plant growth promoting bacteria and humic substances: Crop promotion and mechanisms of action. Chem. Biol. Technol. Agric. 2017, 4, 30. [Google Scholar] [CrossRef] [Green Version]
- Tikhonov, V.V.; Yakushev, A.V.; Zavgorodnyaya, Y.A.; Byzov, B.A.; Demin, V.V. Effects of humic acids on the growth of bacteria. Eurasian J. Soil Sci. 2010, 43, 305–313. [Google Scholar] [CrossRef]
- Klucáková, M. Size and charge evaluation of standard humic and fulvic acids as crucial factors to determine their environmental behavior and impact. Front. Chem. 2018, 6, 235. [Google Scholar] [CrossRef]
- Tang, C.; Li, Y.; Song, J.; Antonietti, M.; Yang, F. Artificial humic substances improve microbial activity for binding CO2. iScience 2021, 24, 102647. [Google Scholar] [CrossRef]
- Arkhipova, T.; Martynenko, E.; Sharipova, G.; Kuzmina, L.; Ivanov, I.; Garipova, M.; Kudoyarova, G. Effects of plant growth promoting rhizobacteria on the content of abscisic acid and salt resistance of wheat plants. Plants 2020, 9, 1429. [Google Scholar] [CrossRef]
- Chetverikov, S.; Vysotskaya, L.; Kuzina, E.; Arkhipova, T.; Bakaeva, M.; Rafikova, G.; Korshunova, T.; Chetverikova, D.; Hkudaygulov, G.; Kudoyarova, G. Effects of association of barley plants with hydrocarbon-degrading bacteria on the content of soluble organic compounds in clean and oil-contaminated sand. Plants 2021, 10, 75. [Google Scholar] [CrossRef] [PubMed]
- Chetverikov, S.P.; Sharipov, D.A.; Korshunova, T.Y.; Loginov, O.N. Degradation of perfluorooctanyl sulfonate by strain Pseudomonas plecoglossicida 2.4-D. Appl. Biochem. Microbiol. 2017, 53, 533–538. [Google Scholar] [CrossRef]
- Bakaeva, M.; Kuzina, E.; Vysotskaya, L.; Kudoyarova, G.; Arkhipova, T.; Rafikova, G.; Chetverikov, S.; Korshunova, T.; Chetverikova, D.; Loginov, O. Capacity of Pseudomonas strains to degrade hydrocarbons, produce auxins and maintain plant growth under normal conditions and in the presence of petroleum contaminants. Plants 2020, 9, 379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kholodov, V.A.; Konstantinov, A.I.; Kudryavtsev, A.V. Structure of humic acids in zonal soils from 13C NMR data. Eurasian Soil Sci. 2011, 44, 976. [Google Scholar] [CrossRef]
- Machado, W.; Franchini, J.C.; Guimarães, M.F.; Filho, J.T. Spectroscopic characterization of humic and fulvic acids in soil aggregates. Brazil. Heliyon. 2020, 6, e04078. [Google Scholar] [CrossRef] [PubMed]
- Volkov, D.S.; Rogova, O.B.; Proskurnin, M.A. Temperature dependences of IR spectra of humic substances of brown coal. Agronomy 2021, 11, 1822. [Google Scholar] [CrossRef]
- Giovanela, M.; Crespo, J.S.; Antunes, M.; Adametti, D.S.; Fernandes, A.N. Chemical and spectroscopic characterization of humic acids extracted from the bottom sediments of a Brazilian subtropical microbasin. J. Mol. Struct. 2010, 981, 111–119. [Google Scholar] [CrossRef] [Green Version]
- Yakimenko, O.; Khundzhua, D.; Izosimov, A.; Yuzhakov, V.; Patsaeva, S. Source indicator of commercial humic products: UV-Vis and fluorescence proxies. J. Soils Sediments 2018, 18, 1279–1291. [Google Scholar] [CrossRef]
- Gosteva, O.Y.; Izosimov, A.A.; Patsaeva, S.V.; Yuzhakov, V.I.; Yakimenko, O.S. Fluorescence of aqueous solutions of commercial humic products. J. Appl. Spectrosc. 2012, 78, 6. [Google Scholar] [CrossRef]
- Tikhonov, V.V.; Orlov, D.S.; Lisovitskaya, O.V.; Zavgorodnyaya, Y.A.; Byzov, B.A.; Demin, V.V. Sorption of humic acids by bacteria. Microbiology 2013, 82, 707–712. [Google Scholar] [CrossRef]
- Moura, M.N.; Martín, F.J.; Burguillo, M.J. A comparative study of the adsorption of humic acid, fulvic acid and phenol onto Bacillus subtilis and activated sludge. J. Hazard. Mater. 2007, 49, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Ojwang, L.M.; Cook, R.L. Environmental conditions that influence the ability of humic acids to induce permeability in model biomembranes. Environ. Sci. Technol. 2013, 47, 8280–8287. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Gu, Z.; Herath, H.M.S.K.; Gu, M.; He, C.; Wang, F.; Jiang, X.; Zhang, J.; Zhang, Y. Evaluation of bacterial biodegradation and accumulation of phenanthrene in the presence of humic acid. Chemosphere 2017, 184, 482–488. [Google Scholar] [CrossRef] [PubMed]
- Farhat, M.B.; Boukhris, I.; Chouayekh, H. Mineral phosphate solubilization by Streptomyces sp. CTM396 involves the excretion of gluconic acid and is stimulated by humic acids. FEMS Microbiol. Lett. 2015, 362, fnv008. [Google Scholar] [CrossRef] [Green Version]
- Shehata, A.A.; Kühnert, M.; Haufe, S.; Krüger, M. Neutralization of the antimicrobial effect of glyphosate by humic acid in vitro. Chemosphere 2014, 104, 258–261. [Google Scholar] [CrossRef]
- Tong, M.; Li, X.; Luo, Q.; Yang, C.; Lou, W.; Liu, H.; Du, C.; Nie, L.; Zhong, Y. Effects of humic acids on biotoxicity of tetracycline to microalgae Coelastrella sp. Algal Res. 2020, 50, 101962. [Google Scholar] [CrossRef]
- Alarcón, M.V.; Salguero, J.; Lloret, P.G. Auxin modulated initiation of lateral roots is linked to pericycle cell length in maize. Front. Plant Sci. 2019, 10, 11. [Google Scholar] [CrossRef] [Green Version]
- Vysotskaya, L.B.; Kudoyarova, G.R.; Arkhipova, T.N.; Kuzina, E.V.; Rafikova, G.F.; Akhtyamova, Z.A.; Ivanov, R.S.; Chetverikov, S.P.; Chetverikova, D.V.; Bakaeva, M.D.; et al. The influence of the association of barley plants with petroleum degrading bacteria on the hormone content, growth and photosynthesis of barley plants grown in the oil-contaminated soil. Acta Phys. Plant 2021, 43, 67. [Google Scholar] [CrossRef]
- De Hita, D.; Fuentes, M.; Fernández, V.; Zamarreño, A.M.; Olaetxea, M.; García-Mina, J.M. Discriminating the short-term action of root and foliar application of humic acids on plant growth: Emerging role of jasmonic acid. Front. Plant Sci. 2020, 11, 493. [Google Scholar] [CrossRef]




| Time of Incubation, h | Additives to the Medium | |||
|---|---|---|---|---|
| No Additives (Control) | HA | FA | HSs | |
| 0 | (3.18 ± 0.14) × 104 a | (3.02 ± 0.21) × 104 a | (3.26 ± 0.23) × 104 a | (2.99 ± 0.17) × 104 a |
| 4 | (3.85 ± 0.43) × 104 a | (4.53 ± 0.38) × 104 a | (3.41 ± 0.30) × 104 a | (4.28 ± 0.63) × 104 a |
| 8 | (2.27 ± 0.60) × 106 a | (5.57 ± 0.99) × 106 b | (5.37 ± 0.84) × 106 b | (1.80 ± 0.70) × 106 a |
| 24 | (1.03 ± 0.14) × 109 a | (1.77 ± 0.29) × 109 a | (1.21 ± 0.09) × 109 a | (1.55 ± 0.20) × 109 a |
| 48 | (1.79 ± 0.18) × 109 a | (2.90 ± 0.19) × 109 b | (2.25 ± 0.20) × 109 a | (2.37 ± 0.25) × 109 a |
| 96 | (2.02 ± 0.16) × 109 a | (6.35 ± 0.68) × 109 c | (7.09 ± 0.45) × 109 c | (4.19 ± 0.36) × 109 b |
| Time of Incubation, Days | Additives to the Medium | |||
|---|---|---|---|---|
| No Additives (Control) | HA | FA | HSs | |
| 0 | (4.07 ± 0.36) × 105 a | (3.85 ± 0.40) × 105 a | (3.80 ± 0.29) × 105 a | (4.11 ± 0.32) × 105 a |
| 1 | (4.15 ± 0.53) × 106 a | (1.14 ± 0.30) × 107 b | (2.87 ± 0.42) × 106 a | (7.20 ± 0.68) × 106 b |
| 2 | (4.12 ± 0.74) × 106 a | (1.22 ± 0.34) × 107 b | (3.10 ± 0.86) × 106 a | (7.62 ± 0.85) × 106 ab |
| 7 | (4.44 ± 0.50) × 106 a | (2.15 ± 0.29) × 107 b | (5.08 ± 0.79) × 106 a | (7.70 ± 0.76) × 106 ab |
| Treatment | Nutrient Medium | |
|---|---|---|
| Nutrient Agar | King Brif | |
| No additives (control) | (5.48 ± 0.49) × 106 a | - |
| Pseudomonas plecoglossicida2,4-D | (1.88 ± 0.09) × 107 d | (1.70 ± 0.14) × 107 a |
| HA | (9.94 ± 0.55) × 106 c | - |
| HA + P. plecoglossicida2,4-D | (4.33 ± 0.15) × 107 e | (3.60 ± 0.26) × 107 c |
| FA | (6.86 ± 0.44) × 106 b | - |
| FA + P. plecoglossicida 2,4-D | (1.89 ± 0.08) × 107 d | (2.03 ± 0.13) × 107 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feoktistova, A.; Bakaeva, M.; Timergalin, M.; Chetverikova, D.; Kendjieva, A.; Rameev, T.; Hkudaygulov, G.; Nazarov, A.; Kudoyarova, G.; Chetverikov, S. Effects of Humic Substances on the Growth of Pseudomonas plecoglossicida 2,4-D and Wheat Plants Inoculated with This Strain. Microorganisms 2022, 10, 1066. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10051066
Feoktistova A, Bakaeva M, Timergalin M, Chetverikova D, Kendjieva A, Rameev T, Hkudaygulov G, Nazarov A, Kudoyarova G, Chetverikov S. Effects of Humic Substances on the Growth of Pseudomonas plecoglossicida 2,4-D and Wheat Plants Inoculated with This Strain. Microorganisms. 2022; 10(5):1066. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10051066
Chicago/Turabian StyleFeoktistova, Arina, Margarita Bakaeva, Maxim Timergalin, Darya Chetverikova, Aliya Kendjieva, Timur Rameev, Gaisar Hkudaygulov, Aleksey Nazarov, Guzel Kudoyarova, and Sergey Chetverikov. 2022. "Effects of Humic Substances on the Growth of Pseudomonas plecoglossicida 2,4-D and Wheat Plants Inoculated with This Strain" Microorganisms 10, no. 5: 1066. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10051066