
Whole-Genome Sequencing Reveals Multiple Subpopulations of Dominant and Persistent Lineage I Isolates of Listeria monocytogenes in Two Meat Processing Facilities during 2011–2015

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. DNA Isolation
2.3. Molecular Confirmation
2.4. Serotyping
2.5. PFGE
2.6. In Silico Analyses
3. Results
3.1. L. monocytogenes Identification and Distribution
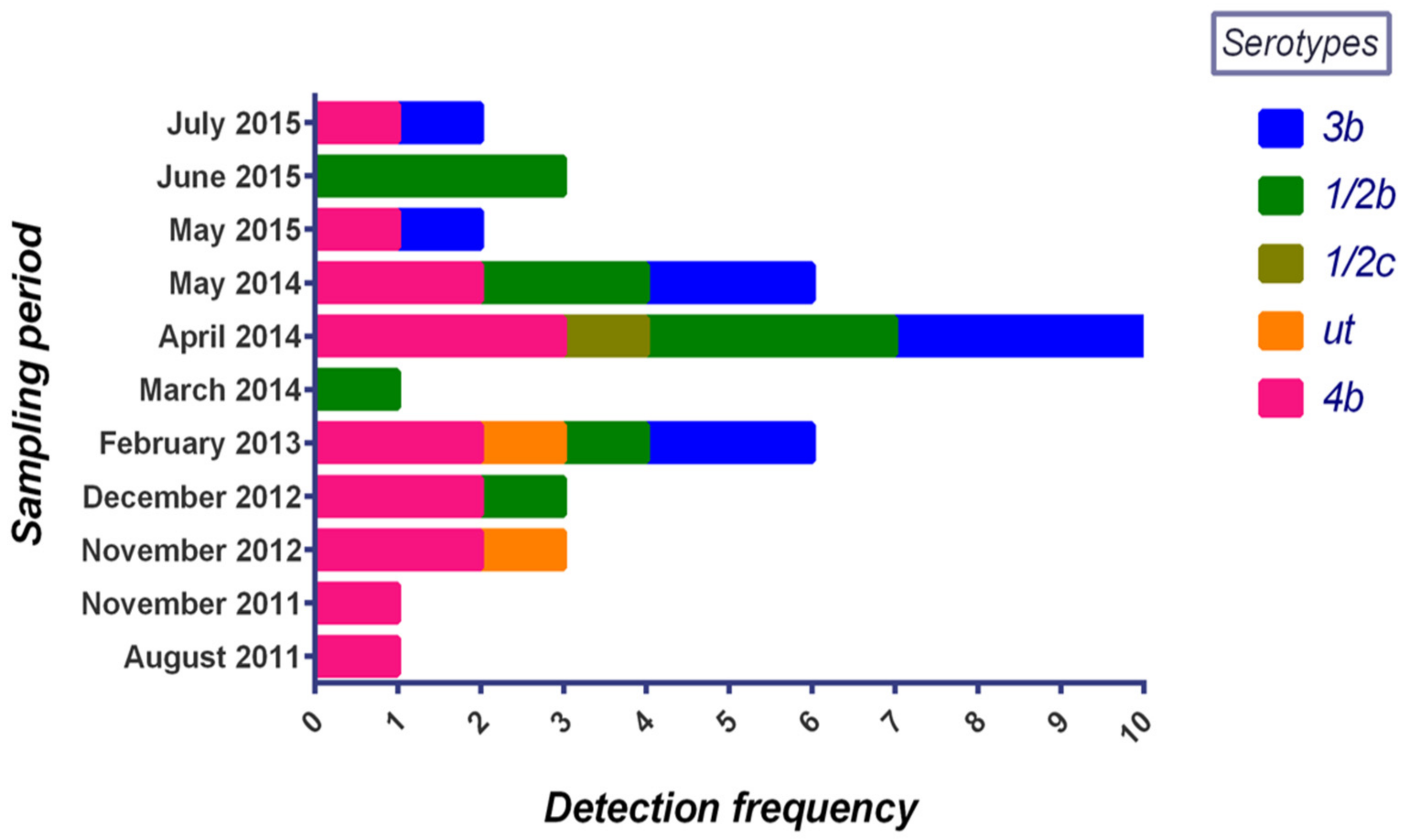
3.2. Serotyping
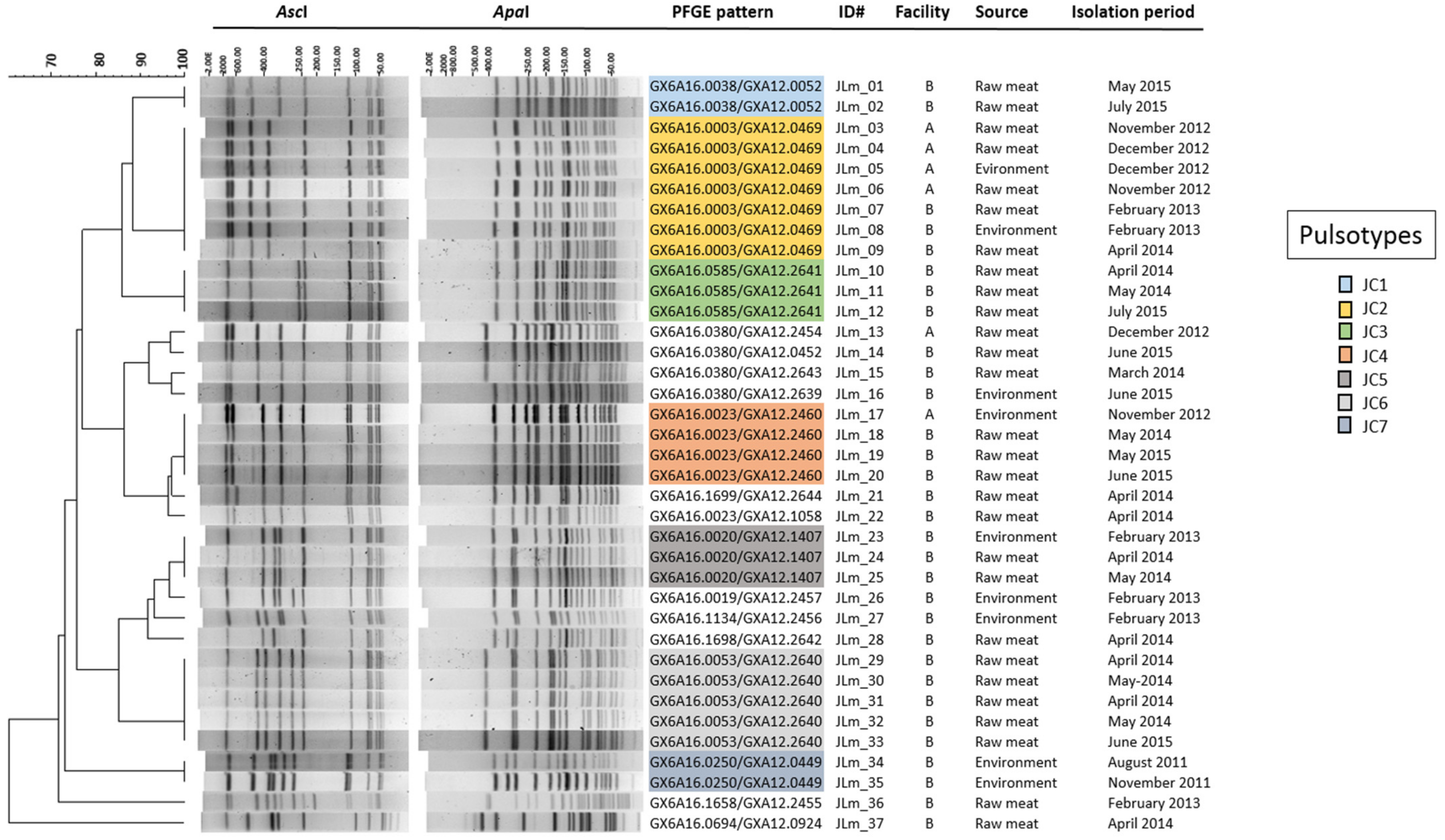
3.3. PFGE Analysis
3.4. wgMLST Analysis
3.5. Virulence and Stress-Related Genes
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Swaminathan, B.; Gerner-Smidt, P. The epidemiology of human listeriosis. Microbes Infect. 2007, 9, 1236–1243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchanan, R.L.; Gorris, L.G.M.; Hayman, M.M.; Jackson, T.C.; Whiting, R.C. A review of Listeria monocytogenes: An update on outbreaks, virulence, dose-response, ecology, and risk assessments. Food Control 2017, 75, 236. [Google Scholar] [CrossRef]
- Camejo, A.; Carvalho, F.; Reis, O.; Leitão, E.; Sousa, S.; Cabanes, D. The arsenal of virulence factors deployed by Listeria monocytogenes to promote its cell infection cycle. Virulence 2011, 2, 379–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cossart, P. Illuminating the interactions with the bacterium Listeria monocytogenes. Proc. Natl. Acad. Sci. USA 2011, 108, 19484–19491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scallan, E.; Hoekstra, R.M.; Angulo, F.J.; Tauxe, R.V.; Widdowson, M.A.; Roy, S.L.; Jones, J.L.; Griffin, P.M. Foodborne illness acquired in the United States—Major pathogens. Emerg. Infect. Dis. 2011, 17, 7. [Google Scholar] [CrossRef]
- Jacquet, C.; Doumith, M.; Gordon, J.I.; Martin, P.M.V.; Cossart, P.; Lecuit, M. A Molecular Marker for Evaluating the Pathogenic Potential of Foodborne Listeria monocytogenes. J. Infect. Dis. 2004, 189, 2094–2100. [Google Scholar] [CrossRef] [Green Version]
- Orsi, R.H.; Bakker, H.C.D.; Wiedmann, M. Listeria monocytogenes lineages: Genomics, evolution, ecology, and phenotypic characteristics. Int. J. Med Microbiol. 2011, 301, 79–96. [Google Scholar] [CrossRef]
- Hoffmann, S.; Batz, M.B.; Morris, J.G., Jr. Annual Cost of Illness and Quality-Adjusted Life Year Losses in the United States Due to 14 Foodborne Pathogens. J. Food Prot. 2012, 75, 1292–1302. [Google Scholar] [CrossRef]
- Lomonaco, S.; Nucera, D.; Filipello, V. The evolution and epidemiology of Listeria monocytogenes in Europe and the United States. Infect. Genet. Evol. 2015, 35, 172–183. [Google Scholar] [CrossRef]
- Kathariou, S. Listeria monocytogenes Virulence and Pathogenicity, a Food Safety Perspective. J. Food Prot. 2002, 65, 1811–1829. [Google Scholar] [CrossRef]
- Moura, A.; Criscuolo, A.; Pouseele, H.; Maury, M.M.; Leclercq, A.; Tarr, C.; Björkman, J.T.; Dallman, T.; Reimer, A.; Enouf, V.; et al. Whole genome-based population biology and epidemiological surveillance of Listeria monocytogenes. Nat. Microbiol. 2016, 2, 16185. [Google Scholar] [CrossRef] [PubMed]
- Knabel, S.J.; Reimer, A.; Verghese, B.; Lok, M.; Ziegler, J.; Farber, J.; Pagotto, F.; Graham, M.; Nadon, C.A.; Gilmour, M.W. Sequence Typing Confirms that a Predominant Listeria monocytogenes Clone Caused Human Listeriosis Cases and Outbreaks in Canada from 1988 to 2010. J. Clin. Microbiol. 2012, 50, 1748–1751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mammina, C.; Parisi, A.; Guaita, A.; Aleo, A.; Bonura, C.; Nastasi, A.; Pontello, M. Enhanced surveillance of invasive listeriosis in the Lombardy region, Italy, in the years 2006-2010 reveals major clones and an increase in serotype 1/2a. BMC Infect. Dis. 2013, 13, 152. [Google Scholar] [CrossRef] [Green Version]
- Yildirim, S.; Lin, W.; Hitchins, A.D.; Jaykus, L.-A.; Altermann, E.; Klaenhammer, T.R.; Kathariou, S. Epidemic Clone I-Specific Genetic Markers in Strains of Listeria monocytogenes Serotype 4b from Foods. Appl. Environ. Microbiol. 2004, 70, 4158–4164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kathariou, S.; Torrence, M.; Isaacson, R. Foodborne outbreaks of listeriosis and epidemic-associated lineages of Listeria monocytogenes. In Microbial Food Safety in Animal Agriculture: Current Topics; Wiley: Hoboken, NJ, USA, 2003; pp. 243–256. [Google Scholar]
- Kovacevic, J.; Arguedas-Villa, C.; Wozniak, A.; Tasara, T.; Allen, K.J. Examination of Food Chain-Derived Listeria monocytogenes Strains of Different Serotypes Reveals Con-siderable Diversity in inlA Genotypes, Mutability, and Adaptation to Cold Temperatures. Appl. Environ. Microbiol. 2013, 79, 1915–1922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gray, M.J.; Zadoks, R.N.; Fortes, E.D.; Dogan, B.; Cai, S.; Chen, Y.; Scott, V.N.; Gombas, D.E.; Boor, K.J.; Wiedmann, M. Listeria monocytogenes Isolates from Foods and Humans Form Distinct but Overlapping Populations. Appl. Environ. Microbiol. 2004, 70, 5833–5841. [Google Scholar] [CrossRef] [Green Version]
- Berrang, M.E.; Meinersmann, R.J.; Frank, J.F.; Ladely, S.R. Colonization of a newly constructed commercial chicken further processing plant with Listeria monocytogenes. J. Food Prot. 2010, 73, 286–291. [Google Scholar] [CrossRef]
- Tasara, T.; Stephan, R. Cold Stress Tolerance of Listeria monocytogenes: A Review of Molecular Adaptive Mechanisms and Food Safety Implications. J. Food Prot. 2006, 69, 1473–1484. [Google Scholar] [CrossRef]
- Djordjevic, D.; Wiedmann, M.; McLandsborough, L.A. Microtiter Plate Assay for Assessment of Listeria monocytogenes Biofilm Formation. Appl. Environ. Microbiol. 2002, 68, 2950–2958. [Google Scholar] [CrossRef] [Green Version]
- Ryan, S.; Begley, M.; Hill, C.; Gahan, C. A five-gene stress survival islet (SSI-1) that contributes to the growth of Listeria monocytogenes in suboptimal conditions. J. Appl. Microbiol. 2010, 109, 984–995. [Google Scholar] [CrossRef]
- Begley, M.; Sleator, R.D.; Gahan, C.G.; Hill, C. Contribution of three bile-associated loci, bsh, pva, and btlB, to gastrointestinal persistence and bile tolerance of Listeria monocytogenes. Infect. Immun. 2005, 73, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, S.; Hill, C.; Gahan, C.G. Acid Stress Responses in Listeria monocytogenes. Adv. Appl. Microbiol. 2008, 65, 67–91. [Google Scholar] [CrossRef] [PubMed]
- Buffet-Bataillon, S.; Tattevin, P.; Bonnaure-Mallet, M.; Jolivet-Gougeon, A. Emergence of resistance to antibacterial agents: The role of quaternary ammonium compounds—A critical review. Int. J. Antimicrob. Agents 2012, 39, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Elhanafi, D.; Dutta, V.; Kathariou, S. Genetic Characterization of Plasmid-Associated Benzalkonium Chloride Resistance Determinants in a Listeria monocytogenes Strain from the 1998-1999 Outbreak. Appl. Environ. Microbiol. 2010, 76, 8231–8238. [Google Scholar] [CrossRef] [Green Version]
- Dutta, V.; Elhanafi, D.; Kathariou, S. Conservation and Distribution of the Benzalkonium Chloride Resistance Cassette bcrABC in Listeria monocytogenes. Appl. Environ. Microbiol. 2013, 79, 6067–6074. [Google Scholar] [CrossRef] [Green Version]
- Mullapudi, S.; Siletzky, R.M.; Kathariou, S. Diverse Cadmium Resistance Determinants in Listeria monocytogenes Isolates from the Turkey Processing Plant Environment. Appl. Environ. Microbiol. 2010, 76, 627–630. [Google Scholar] [CrossRef] [Green Version]
- Ratani, S.S.; Siletzky, R.M.; Dutta, V.; Yildirim, S.; Osborne, J.A.; Lin, W.; Hitchins, A.D.; Ward, T.J.; Kathariou, S. Heavy Metal and Disinfectant Resistance of Listeria monocytogenes from Foods and Food Processing Plants. Appl. Environ. Microbiol. 2012, 78, 6938–6945. [Google Scholar] [CrossRef] [Green Version]
- Parsons, C.; Lee, S.; Jayeola, V.; Kathariou, S. Characterization of a novel cadmium resistance determinant in Listeria monocytogenes. Appl. Environ. Microbiol. 2016, 83, 5. [Google Scholar]
- Palaiodimou, L.; Fanning, S.; Fox, E.M. Genomic insights into persistence of Listeria species in the food processing environment. J. Appl. Microbiol. 2021, 131, 2082–2094. [Google Scholar] [CrossRef]
- Jackson, K.A.; Biggerstaff, M.; Tobin-D’Angelo, M.; Sweat, D.; Klos, R.; Nosari, J.; Garrison, O.; Boothe, E.; Saathoff-Huber, L.; Hainstock, L.; et al. Multistate Outbreak of Listeria monocytogenes Associated with Mexican-Style Cheese Made from Pasteurized Milk among Pregnant, Hispanic Women. J. Food Prot. 2011, 74, 949–953. [Google Scholar] [CrossRef]
- Mead, P.S.; Dunne, E.F.; Graves, L.; Wiedmann, M.; Patrick, M.; Hunter, S.; Salehi, E.; Mostashari, F.; Craig, A.; Mshar, P.; et al. Nationwide outbreak of listeriosis due to contaminated meat. Epidemiology Infect. 2005, 134, 744–751. [Google Scholar] [CrossRef] [PubMed]
- Laksanalamai, P.; Joseph, L.A.; Silk, B.J.; Burall, L.S.; Tarr, C.L.; Gerner-Smidt, P.; Datta, A.R. Genomic Characterization of Listeria monocytogenes Strains Involved in a Multistate Listeriosis Outbreak Associated with Cantaloupe in US. PLoS ONE 2012, 7, e42448. [Google Scholar] [CrossRef] [PubMed]
- Denny, J.; McLauchlin, J. Human Listeria monocytogenes infections in Europe—An opportunity for improved European surveillance. Euro Surveill. 2008, 13, 1854–1861. [Google Scholar] [CrossRef]
- Swaminathan, B.; Barrett, T.J.; Hunter, S.B.; Tauxe, R.V.; Force, C.P.T. PulseNet: The molecular subtyping network for foodborne bacterial disease surveillance, United States. Emerg. Infect. Dis. 2001, 7, 382. [Google Scholar] [CrossRef] [PubMed]
- Leclercq, A.; Charlier, C.; Lecuit, M. Global burden of listeriosis: The tip of the iceberg. Lancet Infect. Dis. 2014, 14, 1027–1028. [Google Scholar] [CrossRef]
- Guerra, M.M.M.; De Almeida, A.M.; Willingham, A.L. An overview of food safety and bacterial foodborne zoonoses in food production animals in the Caribbean region. Trop. Anim. Health Prod. 2016, 48, 1095–1108. [Google Scholar] [CrossRef] [Green Version]
- Lee, B. Foodborne Disease and the Need for Greater Foodborne Disease Surveillance in the Caribbean. Veter- Sci. 2017, 4, 40. [Google Scholar] [CrossRef] [Green Version]
- Syne, S.; Ramsubhag, A.; Adesiyun, A. Occurrence and genetic relatedness of Listeria spp. in two brands of locally pro-cessed ready-to-eat meats in Trinidad. Epidemiol. Infect. 2011, 139, 718–727. [Google Scholar] [CrossRef] [Green Version]
- Syne, S.-M.K. Investigation of the Microbiological Quality of Locally Processed Meats from Two Plants in Trinidad. Doctoral Dissertation, The University of the West Indies, Kingston, Jamaica, 2011. [Google Scholar]
- Syne, S.-M.; Ramsubhag, A.; Adesiyun, A.A. Microbial Quality of Popular Locally Processed Meats Sold in Retail Outlets in Trinidad, West Indies. J. Food Prot. 2015, 78, 333–339. [Google Scholar] [CrossRef]
- Gibbons, I.S.; Adesiyun, A.; Seepersadsingh, N.; Rahaman, S. Investigation for possible source (s) of contamination of ready-to-eat meat products with Listeria spp. and other pathogens in a meat processing plant in Trinidad. Food Microbiol. 2006, 23, 359–366. [Google Scholar] [CrossRef]
- Weissfeld, A.S.; Landes, N.; Livesay, H.; Trevino, E. Listeria monocytogenes Contamination of Ice Cream: A Rare Event That Occurred Twice in the Last Two Years. Clin. Microbiol. Newsl. 2017, 39, 19–22. [Google Scholar] [CrossRef]
- Graves, L.M.; Swaminathan, B. PulseNet standardized protocol for subtyping Listeria monocytogenes by macrorestriction and pulsed-field gel electrophoresis. Int. J. Food Microbiol. 2001, 65, 55–62. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Kramarenko, T.; Roasto, M.; Meremäe, K.; Kuningas, M.; Põltsama, P.; Elias, T. Listeria monocytogenes prevalence and serotype diversity in various foods. Food Control 2013, 30, 24–29. [Google Scholar] [CrossRef]
- Leong, D.; Alvarez-Ordóñez, A.; Jordan, K. Monitoring occurrence and persistence of Listeria monocytogenes in foods and food processing environments in the Republic of Ireland. Front. Microbiol. 2014, 5, 436. [Google Scholar] [CrossRef]
- Ebner, R.; Stephan, R.; Althaus, D.; Brisse, S.; Maury, M.; Tasara, T. Phenotypic and genotypic characteristics of Listeria monocytogenes strains isolated during 2011–2014 from different food matrices in Switzerland. Food Control 2015, 57, 321–326. [Google Scholar] [CrossRef] [Green Version]
- Nightingale, K.K.; Windham, K.; Martin, K.E.; Yeung, M.; Wiedmann, M. Select Listeria monocytogenes Subtypes Commonly Found in Foods Carry Distinct Nonsense Mutations in inlA, Leading to Expression of Truncated and Secreted Internalin A, and Are Associated with a Reduced Invasion Phenotype for Human Intestinal Epithelial Cells. Appl. Environ. Microbiol. 2005, 71, 8764–8772. [Google Scholar]
- Lee, S.; Chen, Y.; Gorski, L.; Ward, T.J.; Osborne, J.; Kathariou, S. Listeria monocytogenes Source Distribution Analysis Indicates Regional Heterogeneity and Ecological Niche Preference among Serotype 4b Clones. mBio 2018, 9, e00396-18. [Google Scholar] [CrossRef] [Green Version]
- Maury, M.M.; Tsai, Y.-H.; Charlier, C.; Touchon, M.; Chenal-Francisque, V.; Leclercq, A.; Criscuolo, A.; Gaultier, C.; Roussel, S.; Brisabois, S.R.A.; et al. Uncovering Listeria monocytogenes hypervirulence by harnessing its biodiversity. Nat. Genet. 2016, 48, 308–313. [Google Scholar] [CrossRef] [Green Version]
- Lachtara, B.; Osek, J.; Wieczorek, K. Molecular Typing of Listeria monocytogenes IVb Serogroup Isolated from Food and Food Production Environments in Poland. Pathogens 2021, 10, 482. [Google Scholar] [CrossRef]
- Chenal-Francisque, V.; Lopez, J.; Cantinelli, T.; Caro, V.; Tran, C.; Leclercq, A.; Lecuit, M.; Brisse, S. Worldwide Distribution of Major Clones of Listeria monocytogenes. Emerg. Infect. Dis. 2011, 17, 1110–1112. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhang, W.; Knabel, S.J. Multi-Virulence-Locus Sequence Typing Identifies Single Nucleotide Polymorphisms Which Differentiate Epidemic Clones and Outbreak Strains of Listeria monocytogenes. J. Clin. Microbiol. 2007, 45, 835–846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D. Handbook of Listeria Monocytogenes; CRC Press: Boca Raton, FL, USA, 2008. [Google Scholar]
- Lappi, V.R.; Thimothe, J.; Nightingale, K.K.; Gall, K.; Scott, V.N.; Wiedmann, M. Longitudinal Studies on Listeria in Smoked Fish Plants: Impact of Intervention Strategies on Contamination Patterns. J. Food Prot. 2004, 67, 2500–2514. [Google Scholar] [CrossRef] [PubMed]
- Quereda, J.J.; Meza-Torres, J.; Cossart, P.; Pizarro-Cerdá, J. Listeriolysin S: A bacteriocin from epidemic Listeria monocytogenes strains that targets the gut microbiota. Gut Microbes 2017, 113, 1–8. [Google Scholar]
- Quereda, J.J.; Dussurget, O.; Nahori, M.-A.; Ghozlane, A.; Volant, S.; Dillies, M.-A.; Regnault, B.; Kennedy, S.; Mondot, S.; Villoing, B.; et al. Bacteriocin from epidemic Listeria strains alters the host intestinal microbiota to favor infection. Proc. Natl. Acad. Sci. USA 2016, 113, 5706–5711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amato, E.; Panteghini, C.; Parisi, A.; Huedo, P.; Gori, M.; Tilola, M.; Losio, N.; Pontello, M.M. A microbial subtyping approach for the identification of food potentially involved in listeriosis in Northern Italy. In Shaping the Future of Food Safety, Together; EFSA: Parma, Italy, 2015; Losio, N. [Google Scholar]
- Huang, Y.-T.; Ko, W.-C.; Chan, Y.-J.; Lu, J.-J.; Tsai, H.-Y.; Liao, C.-H.; Sheng, W.-H.; Teng, L.-J.; Hsueh, P.-R. Disease Burden of Invasive Listeriosis and Molecular Characterization of Clinical Isolates in Taiwan, 2000-2013. PLoS ONE 2015, 10, e0141241. [Google Scholar] [CrossRef]
- Althaus, D.; Lehner, A.; Brisse, S.; Maury, M.; Tasara, T.; Stephan, R. Characterization of Listeria monocytogenes Strains Isolated During 2011–2013 from Human Infections in Switzerland. Foodborne Pathog. Dis. 2014, 11, 753–758. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Luo, Y.; Curry, P.; Timme, R.; Melka, D.; Doyle, M.; Parish, M.; Hammack, T.S.; Allard, M.W.; Brown, E.W.; et al. Assessing the genome level diversity of Listeria monocytogenes from contaminated ice cream and environ-mental samples linked to a listeriosis outbreak in the United States. PLoS ONE 2017, 12, e0171389. [Google Scholar]
- Lomonaco, S.; Verghese, B.; Gerner-Smidt, P.; Tarr, C.; Gladney, L.; Joseph, L.; Katz, L.; Turnsek, M.; Frace, M.; Chen, Y.; et al. Novel epidemic clones of Listeria monocytogenes, United States, 2011. Emerg. Infect. Dis. 2013, 19, 147. [Google Scholar] [CrossRef]
- Chen, Y.; Gonzalez-Escalona, N.; Hammack, T.S.; Allard, M.W.; Strain, E.A.; Brown, E.W. Core genome multilocus sequence typing for the identification of globally distributed clonal groups and differentiation of outbreak strains of Listeria monocytogenes. Appl. Environ. Microbiol. 2016, 82, 20. [Google Scholar] [CrossRef] [Green Version]
- Dreyer, M.; Aguilar-Bultet, L.; Rupp, S.; Guldimann, C.; Stephan, R.; Schock, A.; Otter, A.; Schüpbach-Regula, G.; Brisse, S.; Lecuit, M.; et al. Listeria monocytogenes sequence type 1 is predominant in ruminant rhombencephalitis. Sci. Rep. 2016, 6, 36419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, Y.; Tan, W.; Wang, G.; Kong, S.; Zhou, X.; Zhao, D.; Jia, Y.; Pan, Z.; Jiao, X. Geographical and longitudinal analysis of Listeria monocytogenes genetic diversity reveals its correlation with virulence and unique evolution. Microbiol. Res. 2015, 175, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Wu, Q.; Zhang, J.; Chen, M.; Guo, W. Analysis of Multilocus Sequence Typing and Virulence Characterization of Listeria monocytogenes Isolates from Chinese Retail Ready-to-Eat Food. Front. Microbiol. 2016, 7, 168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cantinelli, T.; Chenal-Francisque, V.; Diancourt, L.; Frezal, L.; Leclercq, A.; Wirth, T.; Lecuit, M.; Brisse, S. “Epidemic clones” of Listeria monocytogenes are widespread and ancient clonal groups. J. Clin. Microbiol. 2013, 51, 3770–3779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gelbíčová, T.; Koláčková, I.; Pantůček, R.; Karpíšková, R. A novel mutation leading to a premature stop codon in inlA of Listeria monocytogenes isolated from neonatal listeriosis. New Microbiol. 2015, 38, 293–296. [Google Scholar] [PubMed]
- Nightingale, K.K.; Ivy, R.A.; Ho, A.J.; Fortes, E.D.; Njaa, B.L.; Peters, R.M.; Wiedmann, M. inlA Premature Stop Codons Are Common among Listeria monocytogenes Isolates from Foods and Yield Virulence-Attenuated Strains That Confer Protection against Fully Virulent Strains. Appl. Environ. Microbiol. 2008, 74, 6570–6583. [Google Scholar] [CrossRef] [Green Version]
- Fletcher, S.; Maharaj, S.R.; James, K. Description of the Food Safety System in Hotels and How It Compares with HACCP Standards. J. Travel Med. 2009, 16, 35–41. [Google Scholar] [CrossRef] [Green Version]
- Mosquera, M.; Evans, E.; Walters, L.; Spreen, T. The US Food Safety Modernization Act: Implications for Caribbean Exporters. Soc. Econ. Stud. 2013, 62, 151–176. [Google Scholar]
- Lundén, J.M.; Autio, T.J.; Sjöberg, A.-M.; Korkeala, H.J. Persistent and Nonpersistent Listeria monocytogenes Contamination in Meat and Poultry Processing Plants. J. Food Prot. 2003, 66, 2062–2069. [Google Scholar] [CrossRef] [Green Version]
- Braga, V.; Vázquez, S.; Vico, V.; Pastorino, V.; Mota, M.I.; Legnani, M.; Schelotto, F.; Lancibidad, G.; Varela, G. Prevalence and serotype distribution of Listeria monocytogenes isolated from foods in Montevideo-Uruguay. Braz. J. Microbiol. 2017, 48, 689–694. [Google Scholar] [CrossRef]
- Montero, D.; Bodero, M.; Riveros, G.; Lapierre, L.; Gaggero, A.; Vidal, R.M.; Vidal, M. Molecular epidemiology and genetic diversity of Listeria monocytogenes isolates from a wide variety of ready-to-eat foods and their relationship to clinical strains from listeriosis outbreaks in Chile. Front. Microbiol. 2015, 6, 384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sant’Ana, A.S.; Igarashi, M.C.; Landgraf, M.; Destro, M.T.; Franco, B.D. Prevalence, populations and pheno- and genotypic characteristics of Listeria monocytogenes isolated from ready-to-eat vegetables marketed in São Paulo, Brazil. Int. J. Food Microbiol. 2012, 155, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Peccio, A.; Autio, T.; Korkeala, H.; Rosmini, R.; Trevisani, M. Listeria monocytogenes occurrence and characterization in meat-producing plants. Lett. Appl. Microbiol. 2003, 37, 234–238. [Google Scholar] [CrossRef] [PubMed]
- Lariviére-Gauthier, G.; Letellier, A.; Kérouanton, A.; Bekal, S.; Quessy, S.; Fournaise, S.; Fravalo, P. Analysis of Listeria monocytogenes Strain Distribution in a Pork Slaughter and Cutting Plant in the Province of Quebec. J. Food Prot. 2014, 77, 2121–2128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markkula, A.; Autio, T.; Lunden, J.; Korkeala, H. Raw and processed fish show identical Listeria monocytogenes genotypes with pulsed-field gel electro-phoresis. J. Food Prot. 2005, 68, 1228–1231. [Google Scholar] [CrossRef] [Green Version]
- Wulff, G.; Gram, L.; Ahrens, P.; Vogel, B.F. One Group of Genetically Similar Listeria monocytogenes Strains Frequently Dominates and Persists in Several Fish Slaughter- and Smokehouses. Appl. Environ. Microbiol. 2006, 72, 4313–4322. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Facility | Source | Total Samples | L. monocytogenes | Listeria spp. |
|---|---|---|---|---|
| A | Raw meat | 6 | 4 | 1 |
| Environment | 2 | 2 | 0 | |
| B | Raw meat | 53 | 24 | 23 |
| Environment | 12 | 7 | 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Burnett, E.; Kucerova, Z.; Freeman, M.; Kathariou, S.; Chen, J.; Smikle, M. Whole-Genome Sequencing Reveals Multiple Subpopulations of Dominant and Persistent Lineage I Isolates of Listeria monocytogenes in Two Meat Processing Facilities during 2011–2015. Microorganisms 2022, 10, 1070. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10051070
Burnett E, Kucerova Z, Freeman M, Kathariou S, Chen J, Smikle M. Whole-Genome Sequencing Reveals Multiple Subpopulations of Dominant and Persistent Lineage I Isolates of Listeria monocytogenes in Two Meat Processing Facilities during 2011–2015. Microorganisms. 2022; 10(5):1070. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10051070
Chicago/Turabian StyleBurnett, Elton, Zuzana Kucerova, Molly Freeman, Sophia Kathariou, Jessica Chen, and Monica Smikle. 2022. "Whole-Genome Sequencing Reveals Multiple Subpopulations of Dominant and Persistent Lineage I Isolates of Listeria monocytogenes in Two Meat Processing Facilities during 2011–2015" Microorganisms 10, no. 5: 1070. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10051070