
Assessing Microbial Corrosion Risk on Offshore Crude Oil Production Topsides under Conditions of Nitrate and Nitrite Treatment for Souring


Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples
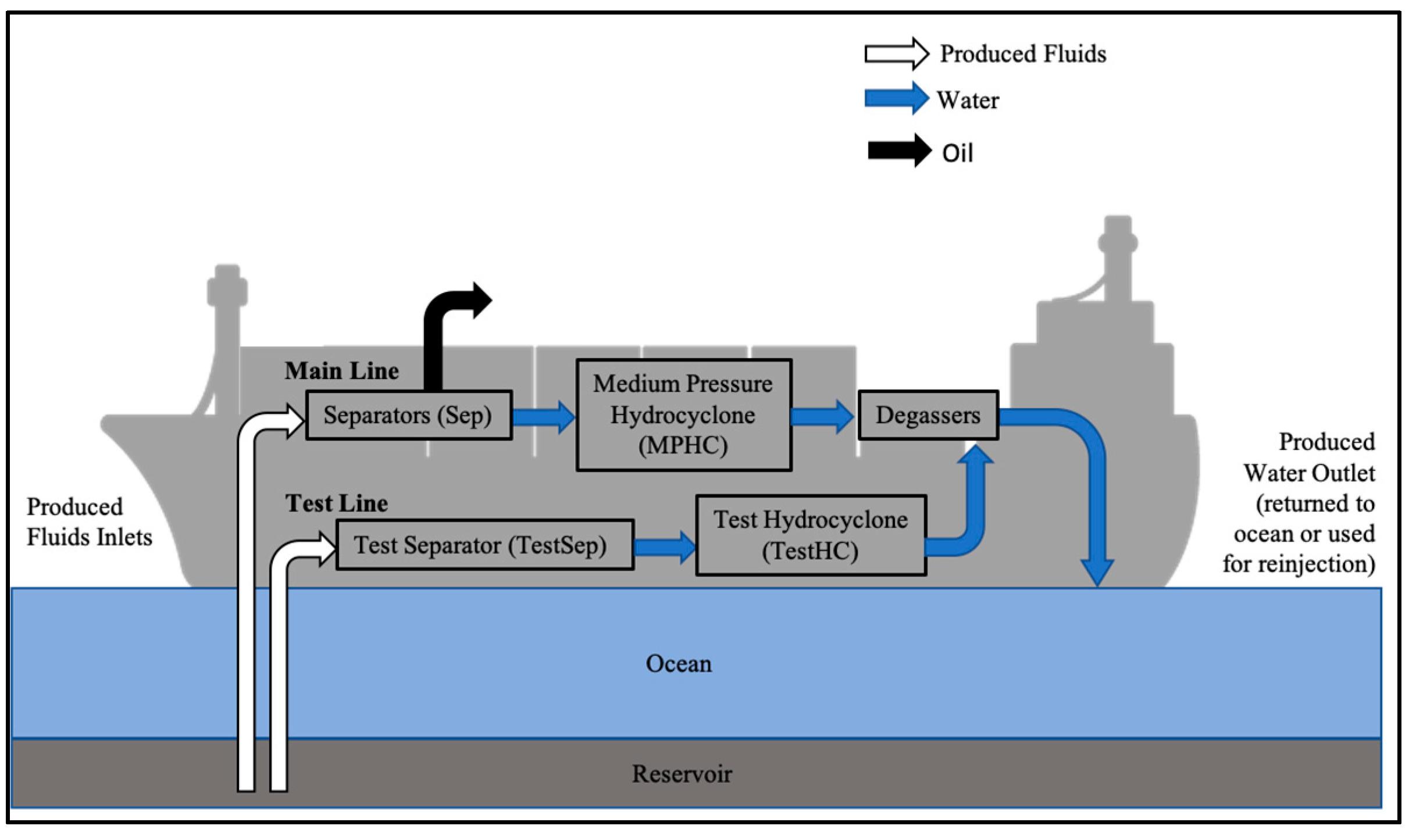
2.2. Microcosm Assembly
2.3. Corrosion Coupon Preparation and Analysis
2.4. Water Chemistry
2.5. Microbial Community Analysis
3. Results
3.1. Produced Water-Sample Analysis
3.1.1. Chemical Analysis of the Produced Water Samples
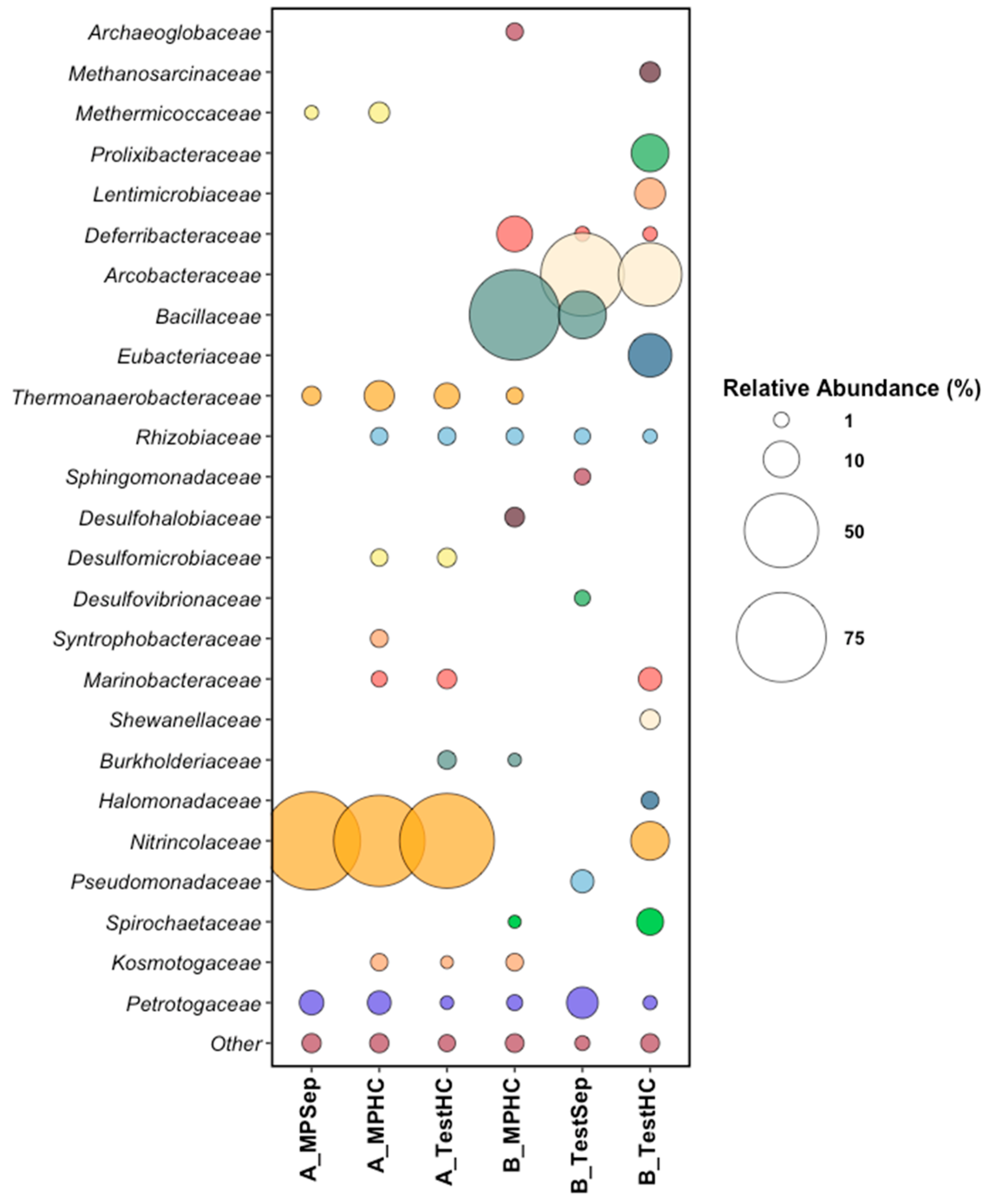
3.1.2. Microbial Analysis of the Produced Water Samples
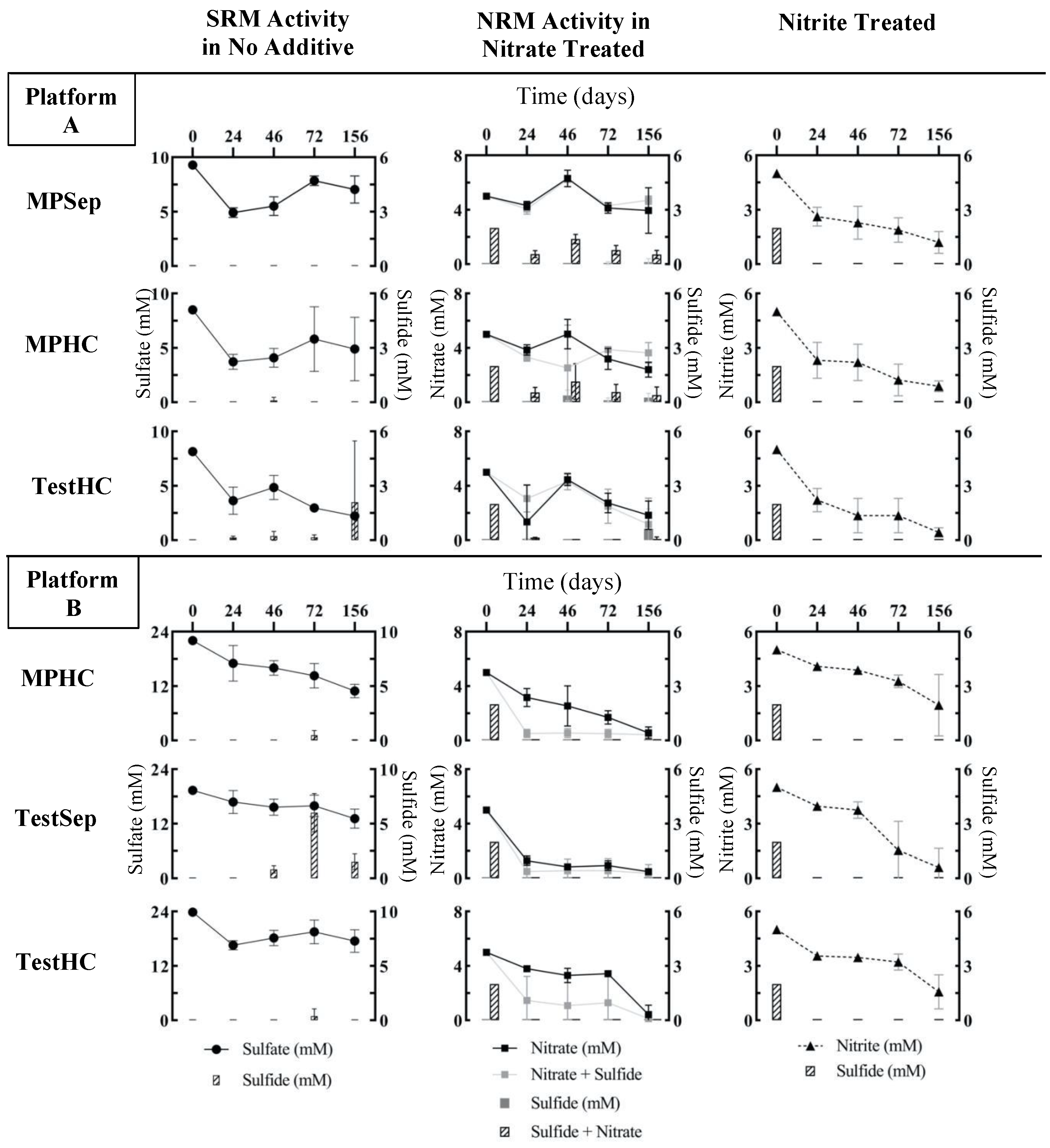
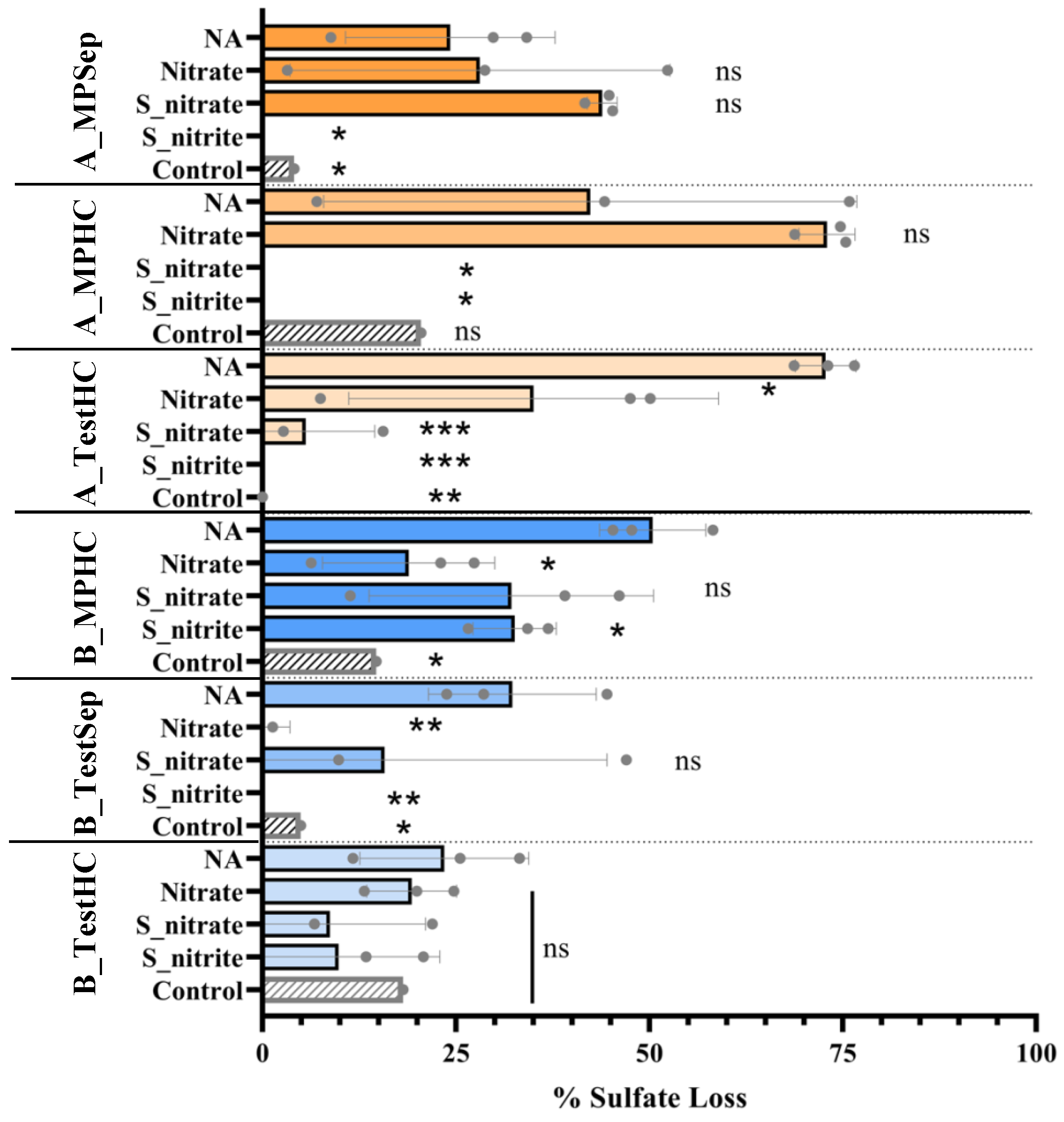
3.2. Effects of Nitrate or Nitrite Treatment on Sulfate Reduction
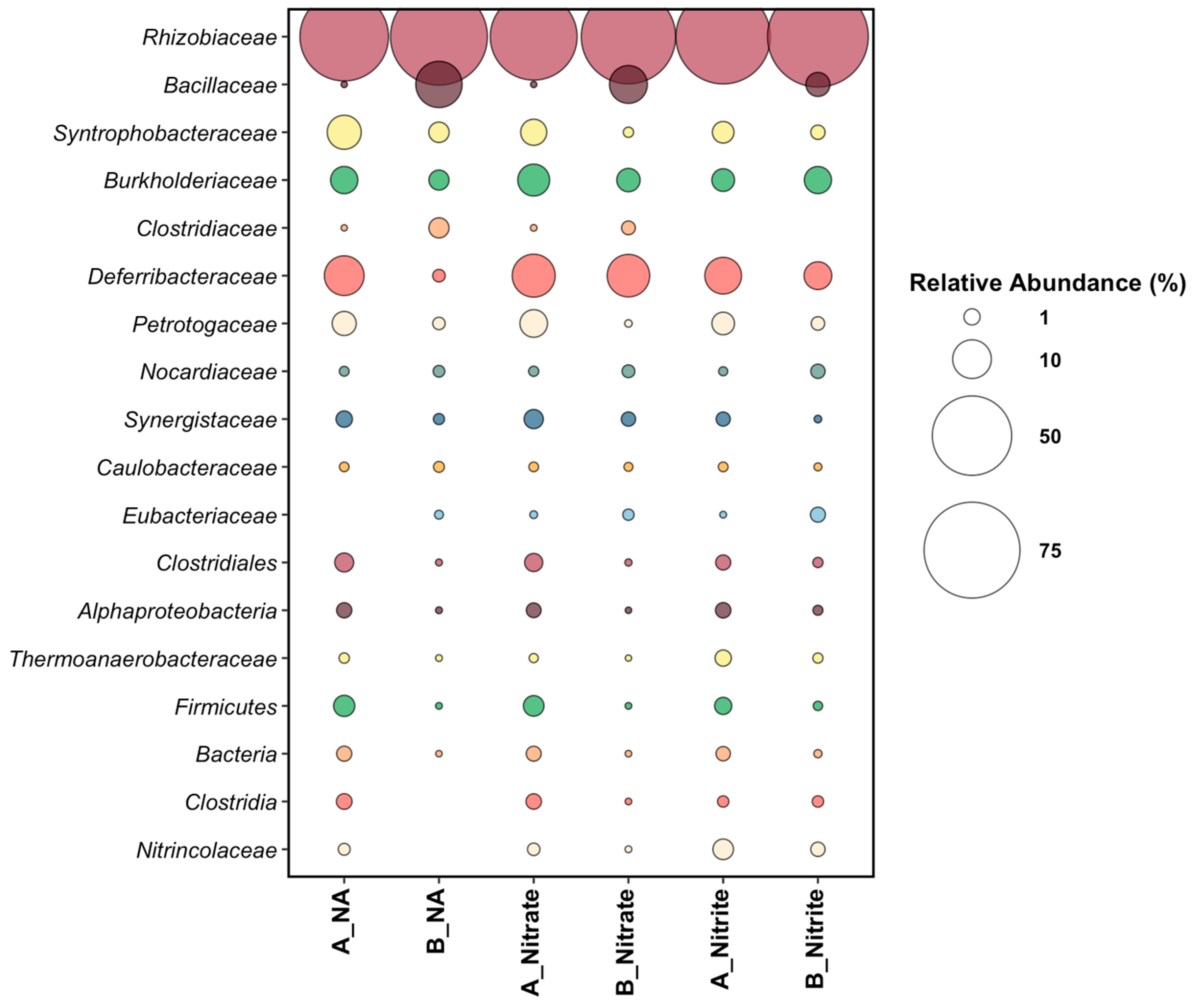
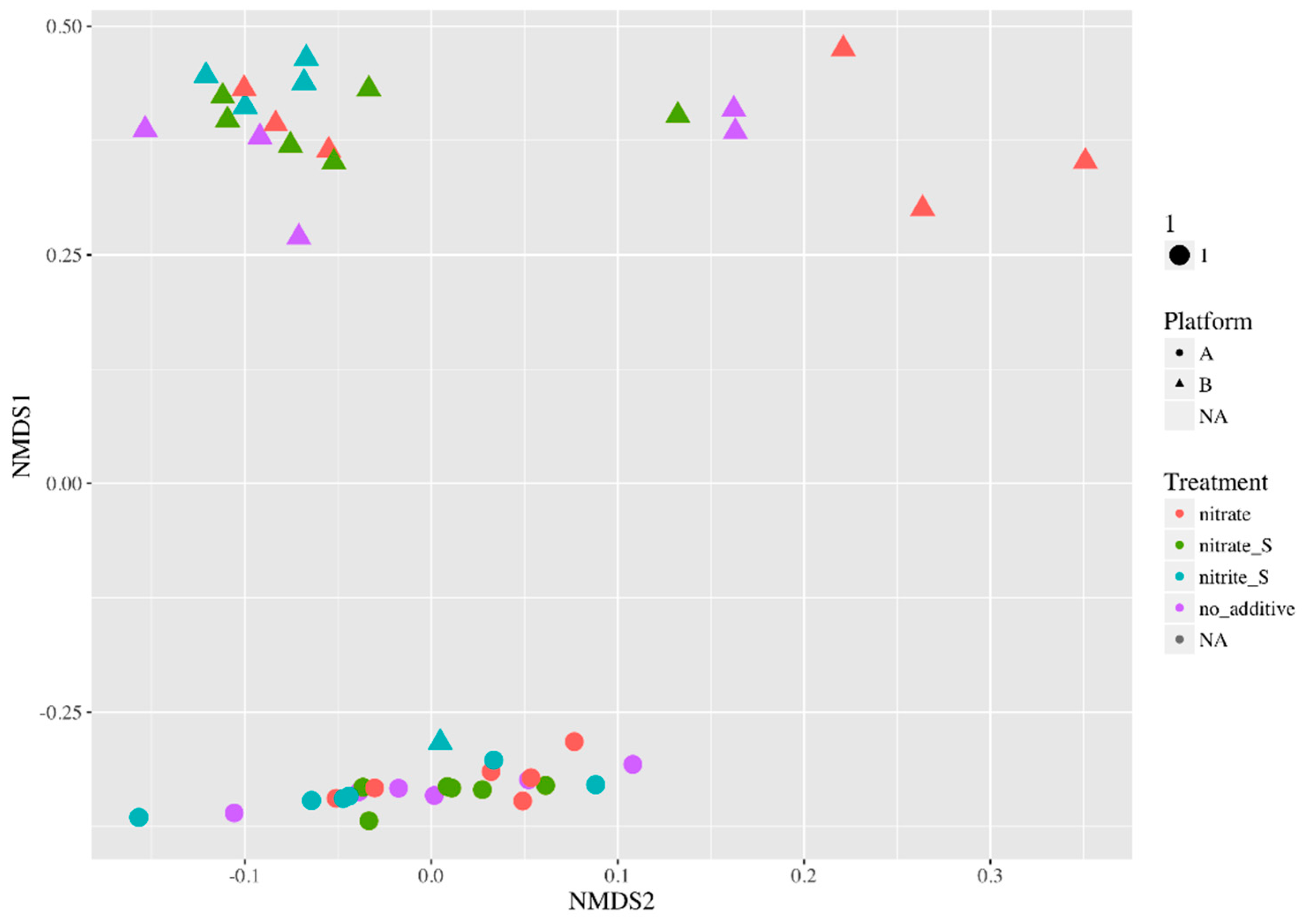
3.3. End-Point Microbial Community Analysis
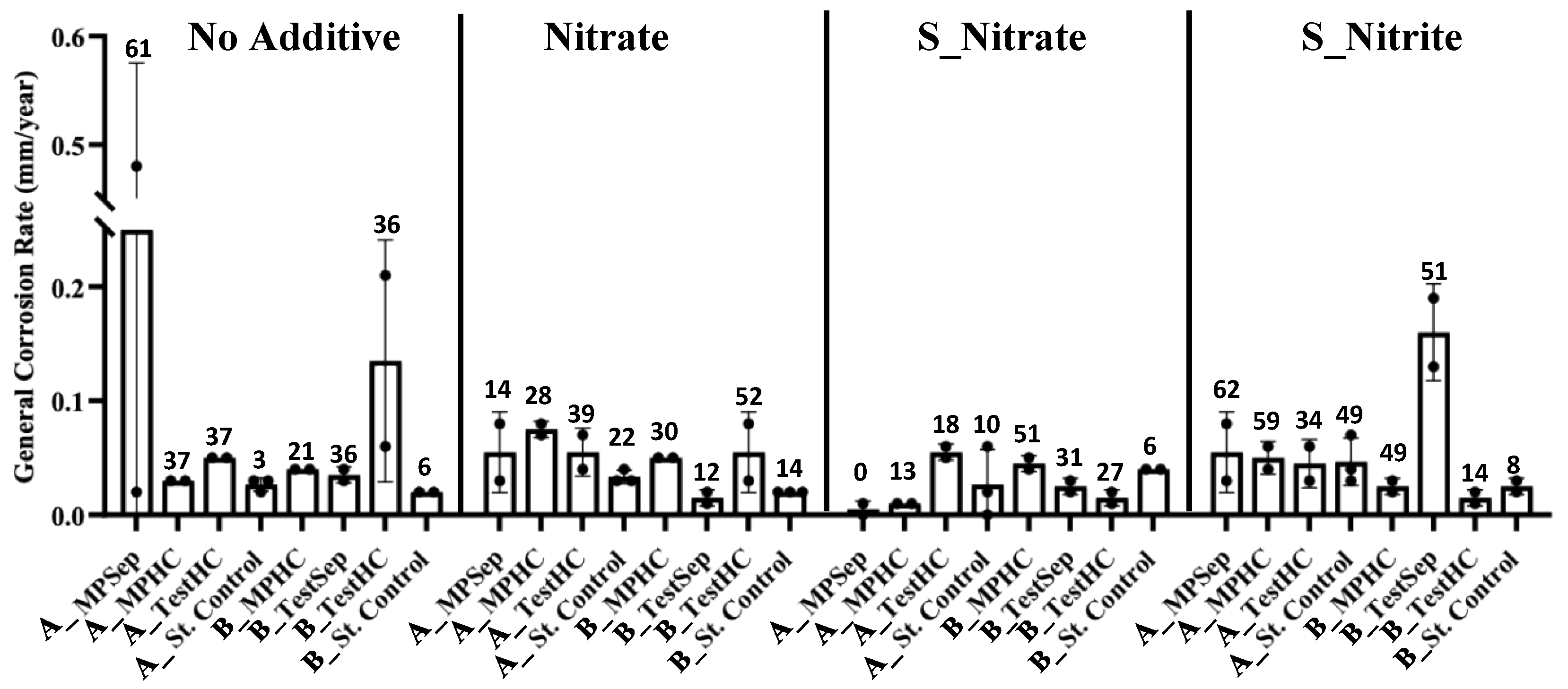
3.4. General Corrosion Rates and Pitting Counts
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- International Energy Agency. Available online: https://www.eia.gov/todayinenergy/detail.php?id=28492 (accessed on 1 March 2022).
- Vigneron, A.; Head, I.A.; Tsesmetzis, N. Damage to offshore production facilities by corrosive microbial biofilms. Appl. Microbiol. Biotech. 2018, 102, 2525–2533. [Google Scholar] [CrossRef] [PubMed]
- Rabus, R.; Venceslau, S.S.; Wohlbrand, L.; Voordouw, G.; Wall, J.; Pereira, I.A.C. A Post Genomic View of the Ecophysiology, Catabolism and Biotechnological Relevance of Sulphate-Reducing Prokaryotes. Adv. Microb. Physiol. 2015, 66, 55–316. [Google Scholar] [PubMed]
- Gaines, R.H. Bacterial activity as a corrosive influence in the soil. Ind. Eng. Chem. 1910, 2, 128–130. [Google Scholar] [CrossRef]
- Bastin, E.S.; Greer, F.E.; Merritt, C.A.; Moulton, G. The presence of sulphate reducing bacteria in oil field waters. Am. Assoc. Advanc. Sci. 1926, 63, 21–24. [Google Scholar] [CrossRef] [PubMed]
- Enning, D.; Garrelfs, J. Corrosion of iron by sulfate-reducing bacteria: New views of an old problem. Appl. Environ. Microbiol. 2014, 80, 1226–1236. [Google Scholar] [CrossRef] [Green Version]
- Gieg, L.M.; Jack, T.R.; Foght, J.M. Biological souring and mitigation in oil reservoirs. Appl. Microbiol. Biotechnol. 2011, 92, 263–282. [Google Scholar] [CrossRef]
- Johnson, R.J.; Folwell, B.D.; Wirekoh, A.; Frenzel, M.; Skovhus, T.L. Reservoir souring—Latest developments for application and mitigation. J. Biotechnol. 2017, 256, 57–67. [Google Scholar] [CrossRef]
- Nemati, M.; Jenneman, G.E.; Voordouw, G. Impact of nitrate-mediated microbial control of souring in oil reservoirs on the extent of corrosion. Biotechn. Prog. 2001, 17, 852–859. [Google Scholar] [CrossRef]
- Hubert, C.; Nemati, M.; Jenneman, G.; Voordouw, G. Corrosion risk associated with microbial souring control using nitrate or nitrite. Appl. Microbiol. Biotechnol. 2005, 68, 272–282. [Google Scholar] [CrossRef]
- van der Kraan, G.M.; Morris, B.E.L.; Widera, I.; Köhler, T.; Keene, P. A microbiological audit of an FPSO. In Proceedings of the Rio Oil & Gas Expo and Conference, Rio de Janeiro, Brazil, 15–18 September 2014; pp. 1–8. [Google Scholar]
- Dolfing, J.; Hubert, C.R.J. Using thermodynamics to predict the outcomes of nitrate-based oil reservoir souring control interventions. Front. Microbiol. 2017, 8, 2575. [Google Scholar] [CrossRef] [Green Version]
- Lahme, S.; Enning, D.; Callbeck, C.M.; Menendez Vega, D.; Curtis, T.P.; Head, I.M.; Hubert, C.R.J. Metabolites of an oil field sulfide-oxidizing, nitrate-reducing Sulfurimonas sp. cause severe corrosion. Appl. Environ. Microbiol. 2019, 85, e01891-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mand, J.; Park, H.S.; Jack, T.R.; Voordouw, G. The role of acetogens in microbially influenced corrosion of steel. Front. Microbiol. 2014, 5, 268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williamson, C.H.D.; Jain, L.A.; Mishra, B.; Olson, D.L.; Spear, J.R. Microbially influenced corrosion communities associated with fuel-grade ethanol environments. Appl. Microbiol. Biotechnol. 2015, 99, 6945–6957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valencia-Cantero, E.; Peña-Cabriales, J.J. Effects of iron-reducing bacteria on carbon steel corrosion induced by thermophilic sulfate-reducing consortia. J. Microbiol. Biotechnol. 2014, 24, 280–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrera, L.K.; Videla, H.A. Role of iron-reducing bacteria in corrosion and protection of carbon steel. Int. Biodeterior. Biodegrad. 2019, 63, 891–895. [Google Scholar] [CrossRef]
- Uchiyama, T.; Ito, K.; Mori, K.; Tsurumaru, H.; Harayama, S. Iron-corroding methanogen isolated from a crude-oil storage tank. Appl. Environ. Microbiol. 2010, 76, 1783–1788. [Google Scholar] [CrossRef] [Green Version]
- Vigneron, A.; Alsop, E.B.; Chambers, B.; Lomans, B.P.; Head, I.M.; Tsesmetzis, N. Complementary microorganisms in highly corrosive biofilms from an offshore oil production facility. Appl. Environ. Microbiol. 2016, 82, 2545–2554. [Google Scholar] [CrossRef] [Green Version]
- Duncan, K.E.; Davidova, I.A.; Nunn, H.S.; Stamps, B.W.; Stevenson, B.S.; Souquet, P.J.; Suflita, J.M. Design features of offshore oil production platforms influence their susceptibility to biocorrosion. Appl. Microbiol. Biotechnol. 2017, 16, 6517–6529. [Google Scholar] [CrossRef]
- Machuca Suarez, L.; Polomka, A. Microbiologically influenced corrosion in floating production systems. Microbiol. Aust. 2018, 39, 165–169. [Google Scholar] [CrossRef]
- Greene, E.A.; Brunelle, V.; Jenneman, G.E.; Voordouw, G. Synergistic inhibition of microbial sulfide production by combinations of the metabolic inhibitor nitrite and biocides. Appl. Environ. Microbiol. 2006, 72, 7897–7901. [Google Scholar] [CrossRef] [Green Version]
- Okpala, G.N.; Chen, C.; Fida, T.; Voordouw, G. Effect of thermophilic nitrate reduction on sulfide production in high temperature oil reservoir samples. Front. Microbiol. 2017, 8, 1573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fida, T.T.; Chen, C.; Okpala, G.; Voordouw, G. Implications of limited thermophilicity of nitrate reduction for controls of sulfide productions in oil reservoirs. Appl. Environ. Microbiol. 2016, 82, 4190–4199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voordouw, G.; Grigoryan, A.A.; Lambo, A.; Lin, S.; Park, H.S.; Jack, T.R.; Coombe, D.; Ertmoed, R.; Miner, K.; Arensdorf, J.J. Sulfide remediation by pulsed injection into a low temperature Canadian heavy oil reservoir. Environ. Sci. Technol. 2009, 43, 9512–9518. [Google Scholar] [CrossRef] [PubMed]
- Vigneron, A.; Alsop, E.B.; Lomans, B.P.; Kyrpides, N.C.; Head, I.M.; Tsesmetzis, N. Succession in the petroleum reservoir microbiome through an oil field production lifecycle. ISME J. 2017, 11, 2141–2154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- NACE International. NACE SP0775-2018: Preparation, Installation, Analysis, and Interpretation of Corrosion Coupons in Oilfield Operations; NACE: Houston, TX, USA, 2018; p. 20. [Google Scholar]
- Mitchell, A.F.; Skjevrak, I.; Waage, J. A re-evaluation of reservoir souring patterns and effect of mitigation in a mature North Sea field. In Proceedings of the SPE International Conference on Oilfield Chemistry, Montgomery, TX, USA, 3–5 April 2017; p. 10. [Google Scholar]
- Sharma, M.; Liu, H.; Chen, S.; Cheng, F.; Voordouw, G.; Gieg, L. Effect of selected biocides on microbiologically influenced corrosion caused by Desulfovibrio ferrophilis IS5. Sci. Rep. 2018, 8, 16620. [Google Scholar] [CrossRef]
- Cline, J.D. Spectrophotometric determination of hydrogen sulfide in natural waters. Limnol. Oceanogr. 1969, 14, 454–458. [Google Scholar] [CrossRef]
- American Public Health Association. Standard Methods for the Examination of Wastewater; American Water Works Association and Water Pollution Control Federation: Washington, DC, USA, 1992; pp. 439–440. [Google Scholar]
- Lovley, D.R.; Phillips, E.J.P. Rapid assay for microbially reducible ferric iron in aquatic sediments. Appl. Environ. Microbiol. 1987, 53, 1536–1540. [Google Scholar] [CrossRef] [Green Version]
- NACE International. NACE TM0194-2014: Field Monitoring of Bacterial Growth in Oil and Gas Systems; NACE: Houston, TX, USA, 2014; p. 31. [Google Scholar]
- Francis, B.; Urich, T.; Mikolasch, A.; Teeling, H.; Amann, R. North Sea spring bloom-associated Gammaproteobacteria fill diverse heterotrophic niches. Environ. Microbiome 2021, 16, 15. [Google Scholar] [CrossRef]
- King, G.M.; Kostka, J.E.; Hazen, T.C.; Sobecky, P.A. Microbial responses to the Deepwater Horizon oil spill: From coastal wetlands to the deep sea. Ann. Rev. Mar. Sci. 2015, 7, 377–401. [Google Scholar] [CrossRef] [Green Version]
- Grassia, G.S.; McLean, K.M.; Glénat, P.; Bauld, J.; Sheehy, A.J. A systematic survey for thermophilic fermentative bacteria and archaea in high temperature petroleum reservoirs. FEMS Microbiol. Ecol. 1996, 21, 47–58. [Google Scholar] [CrossRef]
- L’Haridon, S.; Jiang, L.; Alain, K.; Chalopin, M.; Rouxel, O.; Beauverger, M.; Xu, H.; Shao, Z.; Jebbar, M. Kosmotoga pacifica sp. nov., a thermophilic chemoorganoheterotrophic bacterium isolated from an East Pacific hydrothermal sediment. Extremophiles 2014, 18, 81–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fardeau, M.L.; Faudon, C.; Cayol, J.L.; Magot, M.; Patel, B.K.; Ollivier, B. Effect of thiosulphate as electron acceptor on glucose and xylose oxidation by Thermoanaerobacter finnii and a Thermoanaerobacter sp. isolated from oil field water. Res. Microbiol. 1996, 147, 159–165. [Google Scholar] [CrossRef]
- Fardeau, M.L.; Cayol, J.L.; Magot, M.; Ollivier, B. H2 oxidation in the presence of thiosulfate by a Thermoanaerobacter strain isolated from an oil-producing well. FEMS Microbiol. Lett. 1993, 113, 327–332. [Google Scholar] [CrossRef]
- De Gusseme, B.; De Schryver, P.; De Cooman, M.; Verbeken, K.; Boeckx, P.; Verstraete, W.; Boon, N. Nitrate-reducing, sulfide-oxidizing bacteria as microbial oxidants for rapid biological sulfide removal. FEMS Microbiol. Ecol. 2009, 67, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Wirsen, C.O.; Sievert, S.M.; Cavanaugh, C.M.; Molyneaux, S.J.; Ahmad, A.; Taylor, L.T.; DeLong, E.F.; Taylor, C.D. Characterization of an autotrophic sulphide-oxidizing marine Arcobacter sp. that produces filamentous sulfur. Appl. Environ. Microbiol. 2002, 68, 316–325. [Google Scholar] [CrossRef] [Green Version]
- Evans, M.V.; Panescu, J.; Hanson, A.J.; Welch, S.A.; Sheets, J.M.; Nastasi, N.; Daly, R.A.; Cole, D.R.; Darrah, T.H.; Wilkins, M.J.; et al. Members of Marinobacter and Arcobacter influence system biogeochemistry during early production of hydraulically fractured natural gas wells in the Appalachian Basin. Front. Microbiol. 2018, 9, 2646. [Google Scholar] [CrossRef]
- Duncan, K.E.; Gieg, L.M.; Parisi, V.A.; Tanner, R.S.; Tringe, S.G.; Bristow, J.; Suflita, J.M. Biocorrosive thermophilic microbial communities in Alaskan North Slope oil facilities. Environ. Sci. Technol. 2009, 43, 7977–7984. [Google Scholar] [CrossRef] [Green Version]
- Gittel, A.; Sørensen, K.B.; Skovhus, T.L.; Ingvorsen, K.; Schramm, A. Prokaryotic community structure and sulfate reducer activity in water from high-temperature oil reservoirs with and without nitrate treatment. Appl. Environ. Microbiol. 2009, 75, 7086–7096. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, A.; Hawboldt, K.; Bottaro, C.; Khan, F. Simulation of sour-oxic-nitrite chemical environment in oil and gas facilities. Can. J. Chem. Eng. 2021, 99, S83–S96. [Google Scholar] [CrossRef]
- Alves, L.C.; de Souza, J.A.M.; de Mello Varani, A.; de Macedo Lemos, E.G. The Family Rhizobiaceae; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- Bhattacharya, I.; Das, H.R. Cell surface characteristics of two halotolerant strains of Sinorhizobium meliloti. Microbiol. Res. 2003, 158, 187–194. [Google Scholar] [CrossRef]
- Eckford, R.E.; Fedorak, P.M. Chemical and microbiological changes in laboratory incubations of nitrate amendment ‘sour’ produced waters from three western Canadian oil fields. J. Ind. Microbiol. Biotechnol. 2002, 29, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Reinsel, M.A.; Sears, J.T.; Stewart, P.S.; Mclnerney, M.J. Control of microbial souring by nitrate, nitrite or glutaraldehyde injection in a sandstone column. J. Ind. Microbiol. 1996, 17, 128–136. [Google Scholar] [CrossRef]
- Jahanbani Veshareh, M.; Nick, H.M. A sulfur and nitrogen cycle informed model to simulate nitrate treatment of reservoir souring. Sci. Rep. 2019, 9, 7546. [Google Scholar] [CrossRef] [PubMed]
- Gevertz, D.; Telang, A.J.; Voordouw, G.; Jenneman, G. Isolation and characterization of strains CVO and FWKO B, two novel nitrate-reducing, sulfide-oxidizing bacteria isolated from oil field brine. Appl. Environ. Microbiol. 2000, 66, 2491–2501. [Google Scholar] [CrossRef] [Green Version]
- Callbeck, C.M.; Pelzer, C.; Lavik, G.; Ferdelman, T.G.; Graf, J.S.; Vekeman, B.; Schunck, H.; Littmann, S.; Fuchs, B.M.; Hach, P.F.; et al. Arcobacter perunensis sp. nov., a chemolithoheterotroph isolated from sulfide- and organic-rich coastal water off Peru. Appl. Environ. Microbiol. 2019, 85, e01344-19. [Google Scholar] [CrossRef]
- Slobodkina, G.B.; Kolganova, T.V.; Chernyh, N.A.; Querellou, J.; Bonch-Osmolovskaya, L.; Slobodkin, A.I. Deferribacter autotrophicus sp. nov., an iron(III)-reducing bacterium from a deep-sea hydrothermal vent. Int. J. Syst. Evol. Microbiol. 2009, 59, 1508–1512. [Google Scholar] [CrossRef] [Green Version]
- Duan, J.; Wu, S.; Zhang, X.; Huang, G.; Du, M.; Hou, B. Corrosion of carbon steel influenced by anaerobic biofilm in natural seawater. Electrochim. Acta 2008, 54, 22–28. [Google Scholar] [CrossRef]
- Lenhart, T.R.; Duncan, K.E.; Beech, I.B.; Sunner, J.A.; Smith, W.; Bonifay, V.; Biri, B.; Suflita, J.M. Identification and characterization of microbial biofilm communities associated with corroded oil pipeline surfaces. Biofouling 2014, 30, 823–835. [Google Scholar] [CrossRef]
- Xu, D.; Li, Y.; Song, F.; Gu, T. Laboratory investigation of microbiologically influenced corrosion of C1018 carbon steel by nitrate reducing bacterium Bacillus licheniformis. Corr. Sci. 2013, 77, 385–390. [Google Scholar] [CrossRef]
- Purwasena, I.A.; Astuti, D.I.; Ardini Fauziyyah, N.; Putri, D.A.S.; Sugai, Y. Inhibition of microbial influenced corrosion on carbon steel ST37 using biosurfactant produced by Bacillus sp. Mat. Res. Exp. 2019, 6, 115405. [Google Scholar] [CrossRef]
- Karn, S.K.; Fang, G.; Duan, J. Bacillus sp. acting as dual role for corrosion induction and corrosion inhibition with carbon steel (CS). Front. Microbiol. 2017, 8, 2038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, J.; Madida, B.B. Biofilms affecting progression of mild steel corrosion by Gram positive Bacillus sp. J. Basic Microbiol. 2015, 55, 1168–1178. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Platform | Log No. Cells per mL a | Salinity (M Eq. NaCl) | Temp at Sampling (°C) | pH | S2− (mM) | SO42− (mM) | Total Fe2+/3+ (mM) | NH4+ (mM) | Acetate (mM) |
|---|---|---|---|---|---|---|---|---|---|---|
| A_MPSep | A | 4 | 0.54 | 54 | 7.5 | 0 | 9.3 | 0 | 1.27 | 3.22 |
| A_MPHC | A | 5 | 0.55 | 54 | 7.9 | 0 | 8.6 | 0 | 1.43 | 3.18 |
| A_TestHC | A | 5 | 0.57 | 55 | 7.9 | 0 | 8.1 | 0 | 1.41 | 2.02 |
| B_MPHC | B | 6 | 0.51 | 60 | 7.1 | 0 | 22.0 | 0.53 | 1.20 | 0.79 |
| B_TestSep | B | 6 | 0.47 | 60 | 7.0 | 0 | 19.4 | 0.60 | 1.38 | 1.02 |
| B_TestHC | B | 6 | 0.49 | 54 | 7.1 | 0 | 23.8 | 0.53 | 1.59 | 1.13 |
| Sample | Treatment | Nitrite Production (mM) | ||
|---|---|---|---|---|
| B_MPHC | Nitrate | 0.78 | 0 | 1.03 |
| S_nitrate | 0.96 | 1.11 | 0 | |
| Control | 0 | 0 | 0 | |
| B_TestSep | Nitrate | 0.29 | 2.10 | 2.02 |
| S_nitrate | 0 | 1.78 | 2.45 | |
| Control | 0 | 0 | 0 | |
| B_TestHC | Nitrate | 0 | 0 | 0 |
| S_nitrate | 1.87 | 0.37 | 0 | |
| Control | 0 | 0 | 0 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nicoletti, D.; Sharma, M.; Gieg, L.M. Assessing Microbial Corrosion Risk on Offshore Crude Oil Production Topsides under Conditions of Nitrate and Nitrite Treatment for Souring. Microorganisms 2022, 10, 932. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10050932
Nicoletti D, Sharma M, Gieg LM. Assessing Microbial Corrosion Risk on Offshore Crude Oil Production Topsides under Conditions of Nitrate and Nitrite Treatment for Souring. Microorganisms. 2022; 10(5):932. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10050932
Chicago/Turabian StyleNicoletti, Danika, Mohita Sharma, and Lisa M. Gieg. 2022. "Assessing Microbial Corrosion Risk on Offshore Crude Oil Production Topsides under Conditions of Nitrate and Nitrite Treatment for Souring" Microorganisms 10, no. 5: 932. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10050932