
Evaluation of a Luminometric Cell Counting System in Context of Antimicrobial Photodynamic Inactivation

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Media
2.2. Culture Conditions
2.3. Assessment of Bacterial Viability
2.4. Photodynamic Treatment

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Classification | Gram-Stain | Resistances [33] |
|---|---|---|
| Escherichia coli | negative | Extended spectrum beta-lactamase (ESBL) |
| Staphylococcus aureus | positive | Methicillin-resistant (MRSA) |
| Klebsiella pneumoniae | negative | ESBL-producing, fluoroquinolone-resistant, carbapenem-resistant (CRE) |
| Acinetobacter baumanii | negative | Extensively drug-resistant (XDR) |
| Enterococcus faecium | positive | Vancomycin-resistant (VRE) |
| Pseudomonas aeruginosa | negative | Multidrug-resistant (MDR) (including carbapenems) |
| Morganella morganii | negative | - |
| Achromobacter xylosoxidans | negative | Extensively drug-resistant (XDR) |
2.5. Spectrometry
2.6. Statistical Analysis
3. Results
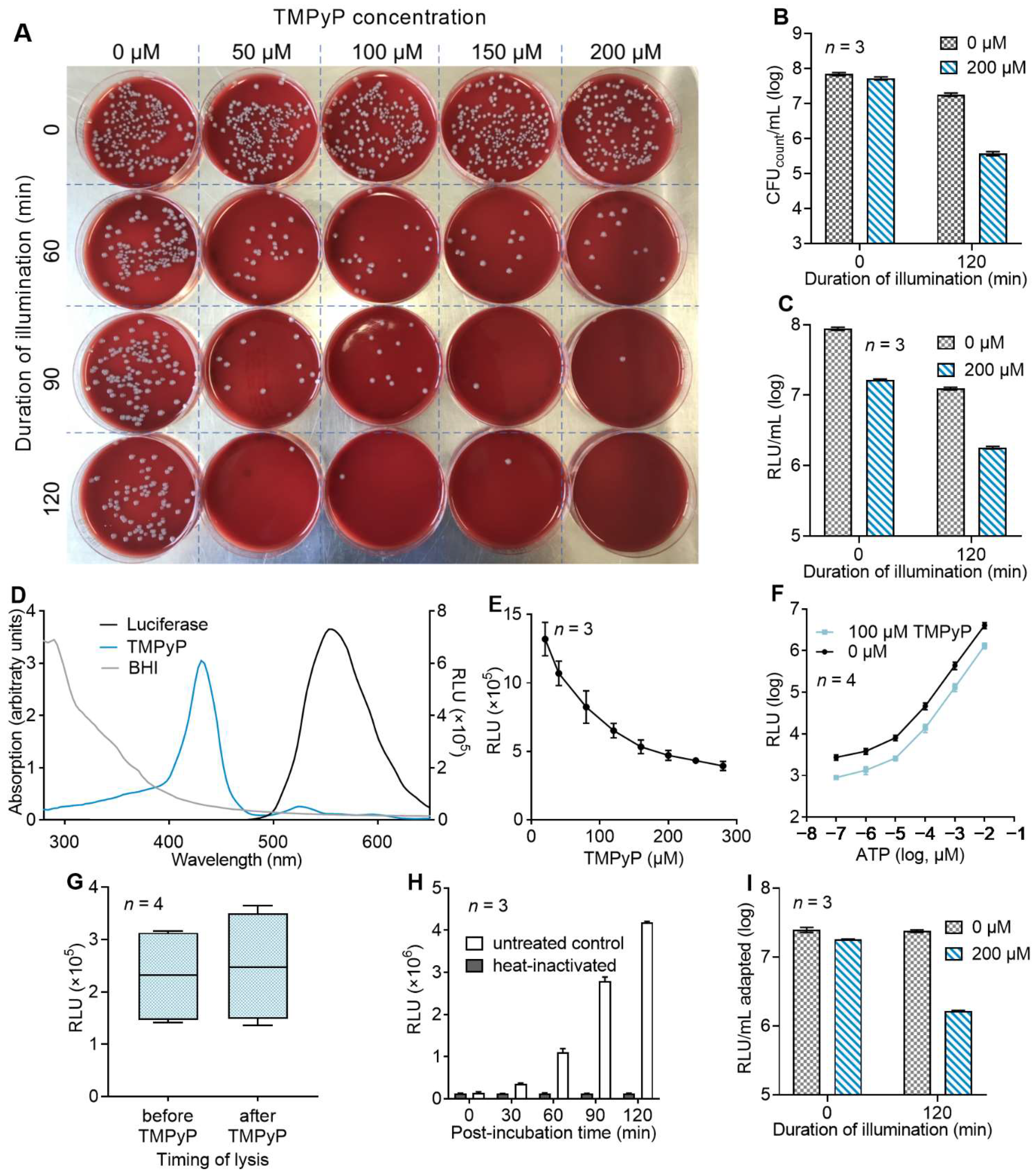
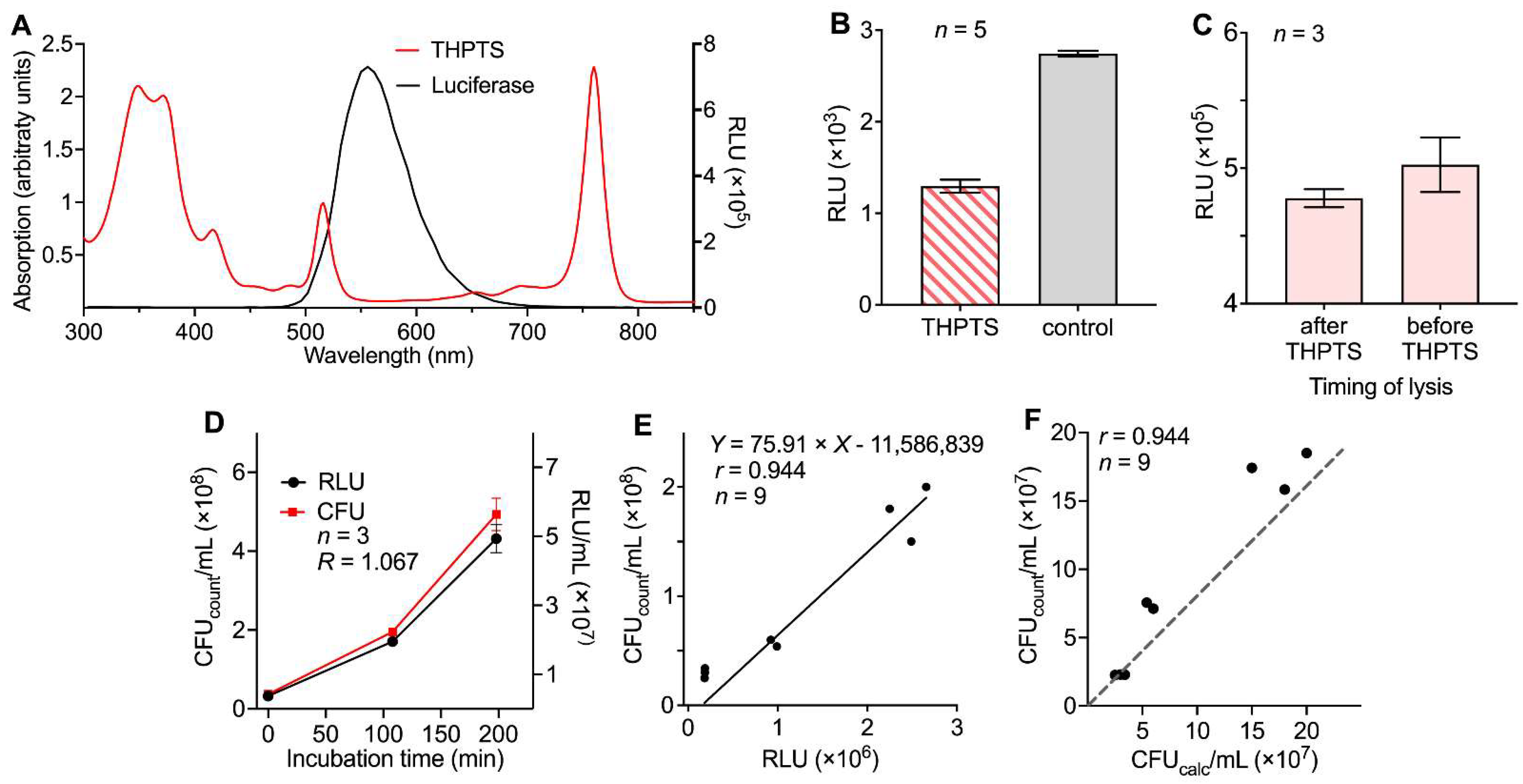
3.1. Evaluation of a Luminescence-Based Approach for the Determination of aPDI Effects
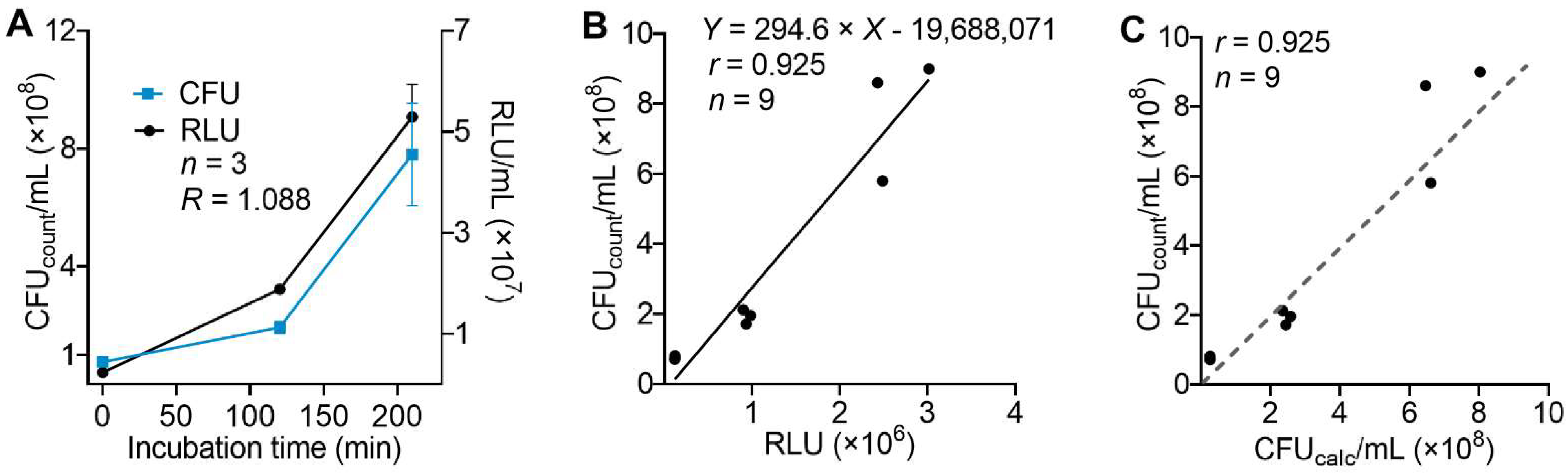
3.2. Correlation of Results for Density of Viable Bacteria Determined by Luminescence or Conventional Colony Counting
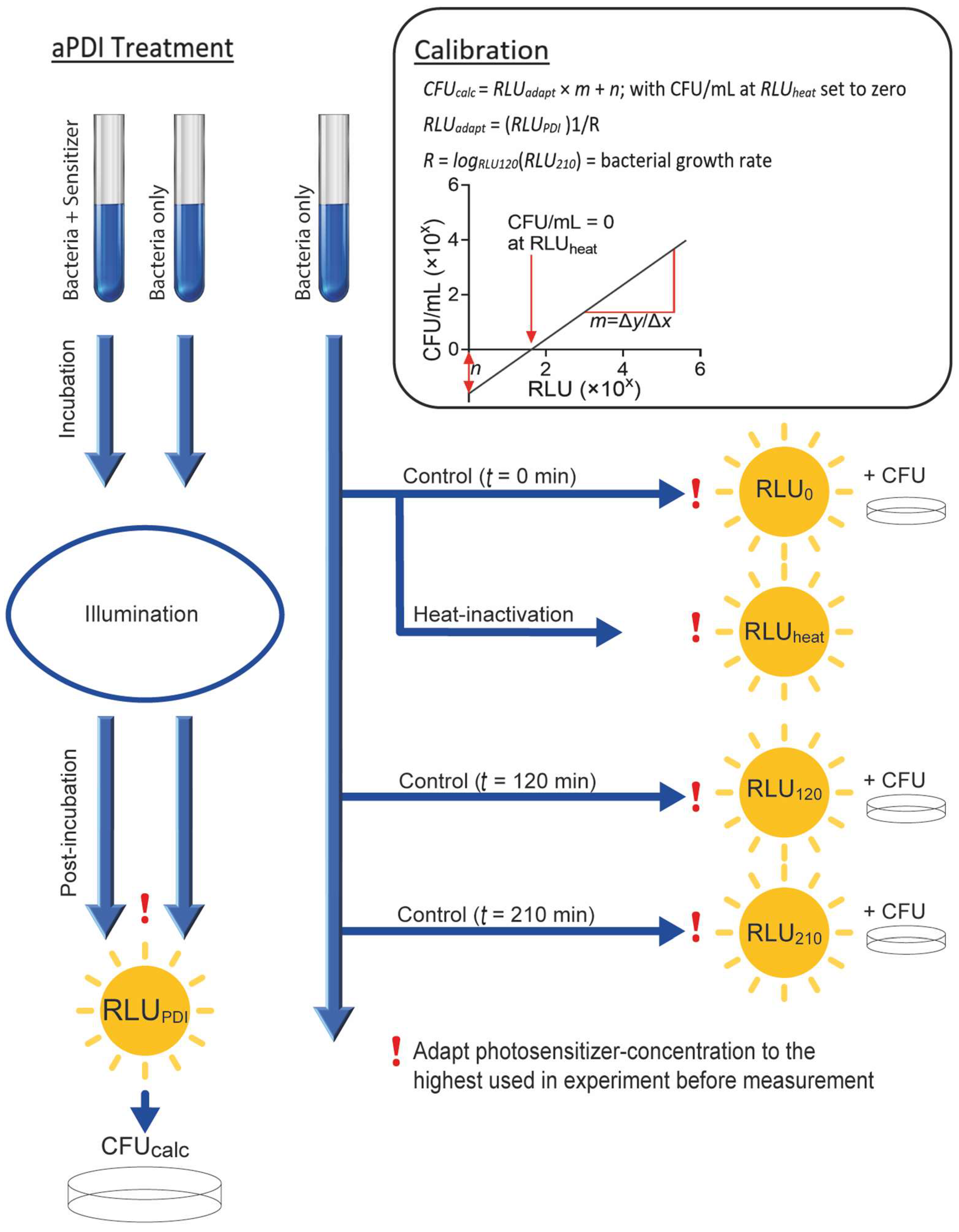
3.3. Standard Protocol to Establish Luminescence-Based Cell Density Determination for aPDI Experiments with Other Photosensitizers and Bacterial Strains
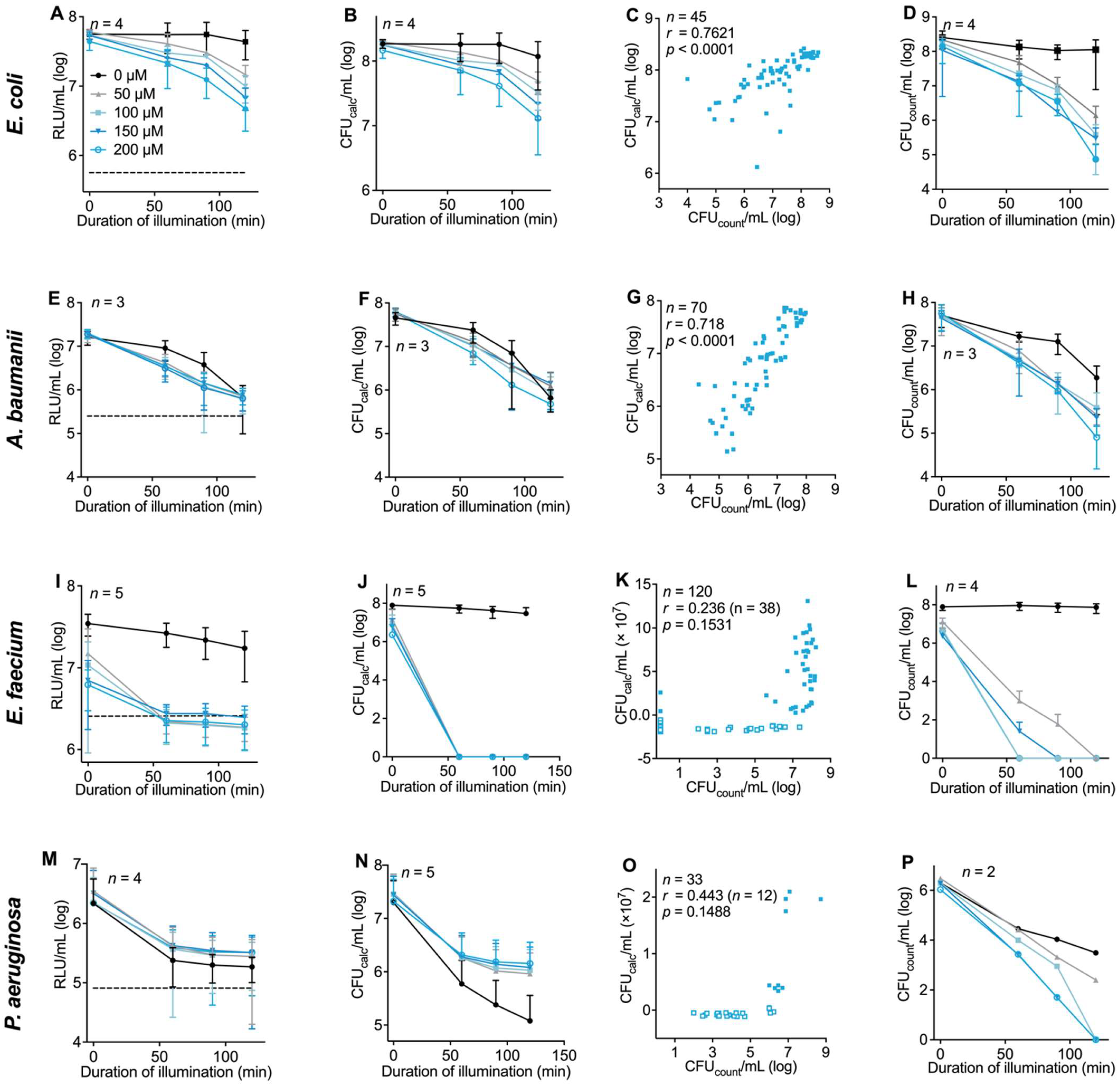
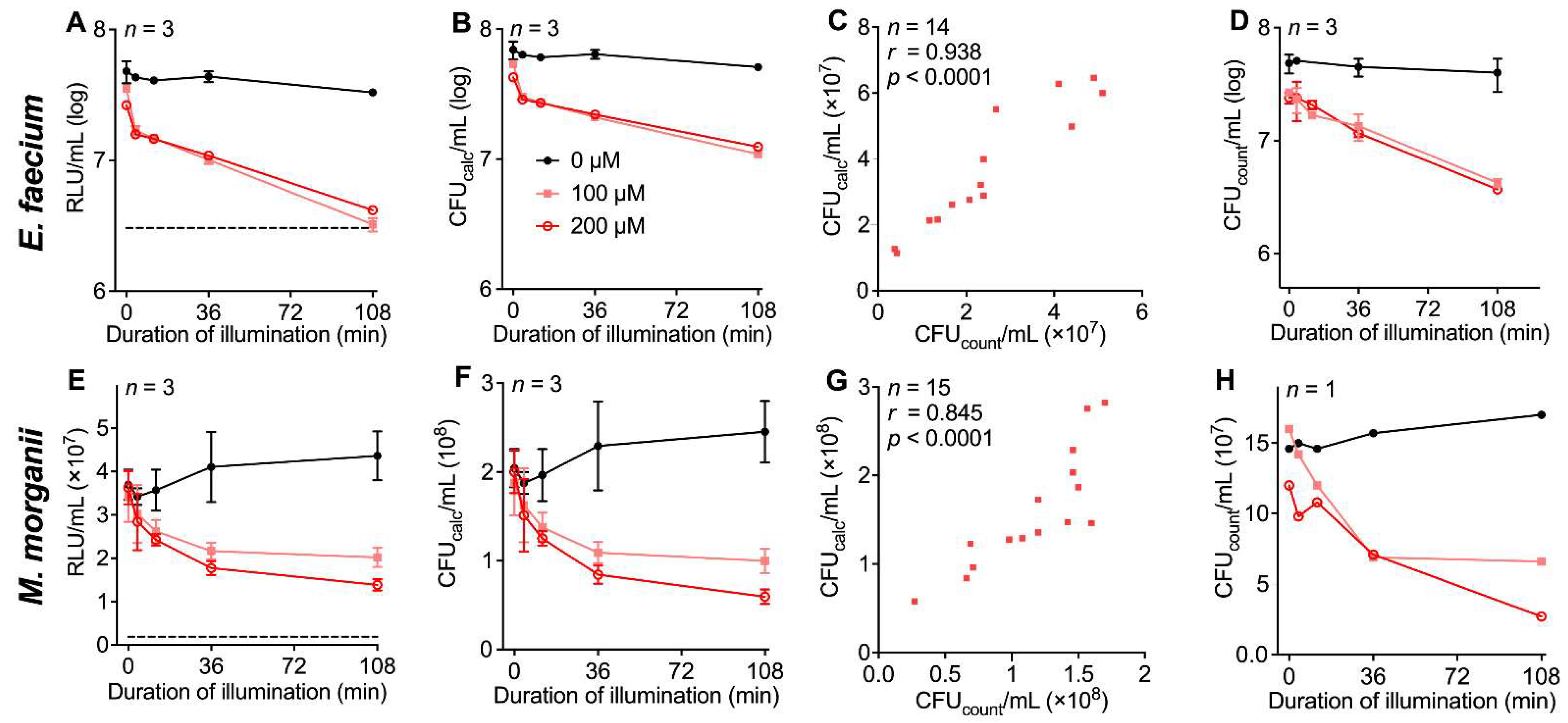
3.4. Evaluation of the Luminescence-Based Assay Protocol with Different Bacterial Strains in the Context of aPDI
3.5. Using the New Protocol for Characterization of aPDI Effects Elicited by Another Photosensitizer
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- WHO. Antimicrobial Resistance—Global Report on Surveillance 2014; World Health Organization: Geneva, Switzerland, 2014.
- Lübbert, C.; Baars, C.; Dayakar, A.; Lippmann, N.; Rodloff, A.C.; Kinzig, M.; Sörgel, F. Environmental Pollution with Antimicrobial Agents from Bulk Drug Manufacturing Industries in Hyderabad, South India, Is Associated with Dissemination of Extended-Spectrum Beta-Lactamase and Carbapenemase-Producing Pathogens. Infection 2017, 45, 479–491. [Google Scholar] [CrossRef] [PubMed]
- Wainwright, M.; Maisch, T.; Nonell, S.; Plaetzer, K.; Almeida, A.; Tegos, G.P.; Hamblin, M.R. Photoantimicrobials-Are We Afraid of the Light? Lancet Infect. Dis. 2017, 17, e49–e55. [Google Scholar] [CrossRef]
- Abrahamse, H.; Hamblin, M.R. New Photosensitizers for Photodynamic Therapy. Biochem. J. 2016, 473, 347–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amos-Tautua, B.M.; Songca, S.P.; Oluwafemi, O.S. Application of Porphyrins in Antibacterial Photodynamic Therapy. Molecules 2019, 24, 2456. [Google Scholar] [CrossRef] [Green Version]
- Ezzeddine, R.; Al-Banaw, A.; Tovmasyan, A.; Craik, J.D.; Batinic-Haberle, I.; Benov, L.T. Effect of Molecular Characteristics on Cellular Uptake, Subcellular Localization, and Phototoxicity of Zn(II) N-Alkylpyridylporphyrins. J. Biol. Chem. 2013, 288, 36579–36588. [Google Scholar] [CrossRef] [Green Version]
- Hamblin, M.R. Antimicrobial Photodynamic Inactivation: A Bright New Technique to Kill Resistant Microbes. Curr. Opin. Microbiol. 2016, 33, 67–73. [Google Scholar] [CrossRef] [Green Version]
- Ragàs, X.; He, X.; Agut, M.; Roxo-Rosa, M.; Gonsalves, A.R.; Serra, A.C.; Nonell, S. Singlet Oxygen in Antimicrobial Photodynamic Therapy: Photosensitizer-Dependent Production and Decay in E. Coli. Molecules 2013, 18, 2712–2725. [Google Scholar] [CrossRef] [Green Version]
- Ergaieg, K.; Chevanne, M.; Cillard, J.; Seux, R. Involvement of Both Type I and Type II Mechanisms in Gram-Positive and Gram-Negative Bacteria Photosensitization by a Meso-Substituted Cationic Porphyrin. Sol. Energy 2008, 82, 1107–1117. [Google Scholar] [CrossRef]
- Gao, Y.; Mai, B.; Wang, A.; Li, M.; Wang, X.; Zhang, K.; Liu, Q.; Wei, S.; Wang, P. Antimicrobial Properties of a New Type of Photosensitizer Derived from Phthalocyanine against Planktonic and Biofilm Forms of Staphylococcus Aureus. Photodiagnosis Photodyn. Ther. 2018, 21, 316–326. [Google Scholar] [CrossRef]
- Imlay, J.A. Pathways of Oxidative Damage. Annu. Rev. Microbiol. 2003, 57, 395–418. [Google Scholar] [CrossRef]
- Wang, J.; Wu, H.; Yang, Y.; Yan, R.; Zhao, Y.; Wang, Y.; Chen, A.; Shao, S.; Jiang, P.; Li, Y.-Q. Bacterial Species-Identifiable Magnetic Nanosystems for Early Sepsis Diagnosis and Extracorporeal Photodynamic Blood Disinfection. Nanoscale 2017, 10, 132–141. [Google Scholar] [CrossRef]
- Martinez De Pinillos Bayona, A.; Mroz, P.; Thunshelle, C.; Hamblin, M.R. Design Features for Optimization of Tetrapyrrole Macrocycles as Antimicrobial and Anticancer Photosensitizers. Chem. Biol. Drug Des. 2017, 89, 192–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cormick, M.P.; Quiroga, E.D.; Bertolotti, S.G.; Alvarez, M.G.; Durantini, E.N. Mechanistic Insight of the Photodynamic Effect Induced by Tri- and Tetra-Cationic Porphyrins on Candida Albicans Cells. Photochem. Photobiol. Sci. 2011, 10, 1556–1561. [Google Scholar] [CrossRef] [PubMed]
- Salmon-Divon, M.; Nitzan, Y.; Malik, Z. Mechanistic Aspects of Escherichia Coli Photodynamic Inactivation by Cationic Tetra-Meso(N-Methylpyridyl)Porphine. Photochem. Photobiol. Sci. 2004, 3, 423–429. [Google Scholar] [CrossRef]
- Tavares, A.; Dias, S.R.S.; Carvalho, C.M.B.; Faustino, M.A.F.; Tomé, J.P.C.; Neves, M.G.P.M.S.; Tomé, A.C.; Cavaleiro, J.A.S.; Cunha, Â.; Gomes, N.C.M.; et al. Mechanisms of Photodynamic Inactivation of a Gram-Negative Recombinant Bioluminescent Bacterium by Cationic Porphyrins. Photochem. Photobiol. Sci. 2011, 10, 1659–1669. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.-J.; Xu, Z.-P.; Ma, Y.-Y.; Ma, J.-D.; Hong, G. Photodynamic Antimicrobial Chemotherapy in Mice with Pseudomonas Aeruginosa-Infected Wounds. PLoS ONE 2020, 15, e0237851. [Google Scholar] [CrossRef]
- Almeida, P.P.; Pereira, Í.S.; Rodrigues, K.B.; Leal, L.S.; Marques, A.S.; Rosa, L.P.; da Silva, F.C.; da Silva, R.A.A. Photodynamic Therapy Controls of Staphylococcus Aureus Intradermal Infection in Mice. Lasers Med. Sci 2017, 32, 1337–1342. [Google Scholar] [CrossRef]
- Branco, T.M.; Valério, N.C.; Jesus, V.I.R.; Dias, C.J.; Neves, M.G.P.M.S.; Faustino, M.A.F.; Almeida, A. Single and Combined Effects of Photodynamic Therapy and Antibiotics to Inactivate Staphylococcus Aureus on Skin. Photodiagnosis Photodyn. Ther. 2018, 21, 285–293. [Google Scholar] [CrossRef]
- Wiegand, I.; Hilpert, K.; Hancock, R.E.W. Agar and Broth Dilution Methods to Determine the Minimal Inhibitory Concentration (MIC) of Antimicrobial Substances. Nat. Protoc. 2008, 3, 163–175. [Google Scholar] [CrossRef]
- McCarthy, J.A.; Thomas, H.A.; Delaney, J.E. Evaluation of the Reliability of Coliform Density Tests. Am. J. Public Health Nations Health 1958, 48, 1628–1635. [Google Scholar] [CrossRef]
- Pazos-Perez, N.; Pazos, E.; Catala, C.; Mir-Simon, B.; Gómez-de Pedro, S.; Sagales, J.; Villanueva, C.; Vila, J.; Soriano, A.; García de Abajo, F.J.; et al. Ultrasensitive Multiplex Optical Quantification of Bacteria in Large Samples of Biofluids. Sci. Rep. 2016, 6, 29014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuji, H.; Matsuda, K.; Nomoto, K. Counting the Countless: Bacterial Quantification by Targeting RRNA Molecules to Explore the Human Gut Microbiota in Health and Disease. Front. Microbiol. 2018, 9, 1417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marques, S.M.; Esteves da Silva, J.C.G. Firefly Bioluminescence: A Mechanistic Approach of Luciferase Catalyzed Reactions. IUBMB Life 2009, 61, 6–17. [Google Scholar] [CrossRef] [PubMed]
- Tannous, B.A. Gaussia Luciferase Reporter Assay for Monitoring Biological Processes in Culture and in Vivo. Nat. Protoc. 2009, 4, 582–591. [Google Scholar] [CrossRef] [PubMed]
- Dingle, T.C.; Butler-Wu, S.M. Maldi-Tof Mass Spectrometry for Microorganism Identification. Clin. Lab. Med. 2013, 33, 589–609. [Google Scholar] [CrossRef] [PubMed]
- Schastak, S.; Jean, B.; Handzel, R.; Kostenich, G.; Hermann, R.; Sack, U.; Orenstein, A.; Wang, Y.-S.; Wiedemann, P. Improved Pharmacokinetics, Biodistribution and Necrosis in Vivo Using a New near Infra-Red Photosensitizer: Tetrahydroporphyrin Tetratosylat. J. Photochem. Photobiol. B 2005, 78, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Riyad, Y.M.; Naumov, S.; Schastak, S.; Griebel, J.; Kahnt, A.; Häupl, T.; Neuhaus, J.; Abel, B.; Hermann, R. Chemical Modification of a Tetrapyrrole-Type Photosensitizer: Tuning Application and Photochemical Action beyond the Singlet Oxygen Channel. J. Phys. Chem. B 2014, 118, 11646–11658. [Google Scholar] [CrossRef]
- Habermann, N.; Wachs, M.; Schulz, S.; Werdehausen, R.; Schwarz, U.T. Development and Characterization of Planar LED Arrays for Medical Applications. Jpn. J. Appl. Phys. 2019, 58, SCCC25. [Google Scholar] [CrossRef]
- Beirão, S.; Fernandes, S.; Coelho, J.; Faustino, M.A.F.; Tomé, J.P.C.; Neves, M.G.P.M.S.; Tomé, A.C.; Almeida, A.; Cunha, A. Photodynamic Inactivation of Bacterial and Yeast Biofilms With a Cationic Porphyrin. Photochem. Photobiol. 2014, 90, 1387–1396. [Google Scholar] [CrossRef]
- Prasanth, C.S.; Karunakaran, S.C.; Paul, A.K.; Kussovski, V.; Mantareva, V.; Ramaiah, D.; Selvaraj, L.; Angelov, I.; Avramov, L.; Nandakumar, K.; et al. Antimicrobial Photodynamic Efficiency of Novel Cationic Porphyrins towards Periodontal Gram-Positive and Gram-Negative Pathogenic Bacteria. Photochem. Photobiol. 2014, 90, 628–640. [Google Scholar] [CrossRef]
- Ziganshyna, S.; Guttenberger, A.; Lippmann, N.; Schulz, S.; Bercker, S.; Kahnt, A.; Rüffer, T.; Voigt, A.; Gerlach, K.; Werdehausen, R. Tetrahydroporphyrin-Tetratosylate (THPTS)-Based Photodynamic Inactivation of Critical Multidrug-Resistant Bacteria in Vitro. Int. J. Antimicrob. Agents 2020, 55, 105976. [Google Scholar] [CrossRef] [PubMed]
- Magiorakos, A.-P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-Resistant, Extensively Drug-Resistant and Pandrug-Resistant Bacteria: An International Expert Proposal for Interim Standard Definitions for Acquired Resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [Green Version]
- Glass, S.; Kühnert, M.; Lippmann, N.; Zimmer, J.; Werdehausen, R.; Abel, B.; Eulenburg, V.; Schulze, A. Photosensitizer-Loaded Hydrogels for Photodynamic Inactivation of Multirestistant Bacteria in Wounds. RSC Adv. 2021, 11, 7600–7609. [Google Scholar] [CrossRef] [PubMed]
- Cox, C.D.; Adams, P. Siderophore Activity of Pyoverdin for Pseudomonas Aeruginosa. Infect. Immun. 1985, 48, 130–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kramer, B.; Wunderlich, J.; Muranyi, P. ATP-Synthesis Capacity of Pulsed Light-Exposed Bacteria. Syst. Appl. Microbiol. 2017, 40, 383–387. [Google Scholar] [CrossRef] [PubMed]
- Al-Mutairi, R.; Tovmasyan, A.; Batinic-Haberle, I.; Benov, L. Sublethal Photodynamic Treatment Does Not Lead to Development of Resistance. Front. Microbiol. 2018, 9, 1699. [Google Scholar] [CrossRef] [Green Version]
- Kakihana, Y.; Cheng, L.; Fang, L.-F.; Wang, S.-Y.; Jeon, S.; Saeki, D.; Rajabzadeh, S.; Matsuyama, H. Preparation of Positively Charged PVDF Membranes with Improved Antibacterial Activity by Blending Modification: Effect of Change in Membrane Surface Material Properties. Colloids Surf. A Physicochem. Eng. Asp. 2017, 533, 133–139. [Google Scholar] [CrossRef]
- Knauf, G.A.; Cunningham, A.L.; Kazi, M.I.; Riddington, I.M.; Crofts, A.A.; Cattoir, V.; Trent, M.S.; Davies, B.W. Exploring the Antimicrobial Action of Quaternary Amines against Acinetobacter Baumannii. mBio 2018, 9, e02394–e02417. [Google Scholar] [CrossRef] [Green Version]
- Hoque, J.; Adhikary, U.; Yadav, V.; Samaddar, S.; Konai, M.M.; Prakash, R.G.; Paramanandham, K.; Shome, B.R.; Sanyal, K.; Haldar, J. Chitosan Derivatives Active against Multidrug-Resistant Bacteria and Pathogenic Fungi: In Vivo Evaluation as Topical Antimicrobials. Mol. Pharm. 2016, 13, 3578–3589. [Google Scholar] [CrossRef]
- Yao, F.; Fu, G.-D.; Zhao, J.; Kang, E.-T.; Neoh, K.G. Antibacterial Effect of Surface-Functionalized Polypropylene Hollow Fiber Membrane from Surface-Initiated Atom Transfer Radical Polymerization. J. Membr. Sci. 2008, 319, 149–157. [Google Scholar] [CrossRef]
- Sułek, A.; Pucelik, B.; Kobielusz, M.; Barzowska, A.; Dąbrowski, J.M. Photodynamic Inactivation of Bacteria with Porphyrin Derivatives: Effect of Charge, Lipophilicity, ROS Generation, and Cellular Uptake on Their Biological Activity In Vitro. Int. J. Mol. Sci. 2020, 21, 8716. [Google Scholar] [CrossRef] [PubMed]
- Faraj Tabrizi, P.; Wennige, S.; Berneburg, M.; Maisch, T. Susceptibility of SodA- and SodB-Deficient Escherichia Coli Mutant towards Antimicrobial Photodynamic Inactivation via the Type I-Mechanism of Action. Photochem. Photobiol. Sci. 2018, 17, 352–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tegos, G.P.; Masago, K.; Aziz, F.; Higginbotham, A.; Stermitz, F.R.; Hamblin, M.R. Inhibitors of Bacterial Multidrug Efflux Pumps Potentiate Antimicrobial Photoinactivation. Antimicrob. Agents Chemother. 2008, 52, 3202–3209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berndt-Paetz, M.; Schulze, P.; Stenglein, P.C.; Weimann, A.; Wang, Q.; Horn, L.-C.; Riyad, Y.M.; Griebel, J.; Hermann, R.; Glasow, A.; et al. Reduction of Muscle-Invasive Tumors by Photodynamic Therapy with Tetrahydroporphyrin-Tetratosylat in an Orthotopic Rat Bladder Cancer Model. Mol. Cancer Ther. 2019, 18, 743–750. [Google Scholar] [CrossRef] [Green Version]
- Berndt-Paetz, M.; Weimann, A.; Sieger, N.; Schastak, S.; Riyad, Y.M.; Griebel, J.; Arthanareeswaran, V.K.A.; Stolzenburg, J.-U.; Neuhaus, J. Tetrahydroporphyrin-Tetratosylat (THPTS): A near-Infrared Photosensitizer for Targeted and Efficient Photodynamic Therapy (PDT) of Human Bladder Carcinoma. An in Vitro Study. Photodiagnosis Photodyn. Ther. 2017, 18, 244–251. [Google Scholar] [CrossRef]
- Walther, J.; Schastak, S.; Dukic-Stefanovic, S.; Wiedemann, P.; Neuhaus, J.; Claudepierre, T. Efficient Photodynamic Therapy on Human Retinoblastoma Cell Lines. PLoS ONE 2014, 9, e87453. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lehnig, M.; Glass, S.; Lippmann, N.; Ziganshyna, S.; Eulenburg, V.; Werdehausen, R. Evaluation of a Luminometric Cell Counting System in Context of Antimicrobial Photodynamic Inactivation. Microorganisms 2022, 10, 950. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10050950
Lehnig M, Glass S, Lippmann N, Ziganshyna S, Eulenburg V, Werdehausen R. Evaluation of a Luminometric Cell Counting System in Context of Antimicrobial Photodynamic Inactivation. Microorganisms. 2022; 10(5):950. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10050950
Chicago/Turabian StyleLehnig, Moritz, Sarah Glass, Norman Lippmann, Svitlana Ziganshyna, Volker Eulenburg, and Robert Werdehausen. 2022. "Evaluation of a Luminometric Cell Counting System in Context of Antimicrobial Photodynamic Inactivation" Microorganisms 10, no. 5: 950. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10050950