
Exploring Mitogenomes Diversity of Fusarium musae from Banana Fruits and Human Patients

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. DNA Extraction and Sequencing
2.2. Other Fungal Mitochondrial Genomes
2.3. Assembly and Annotation of Mitogenome
2.4. Alignment of Protein Genes
2.5. MGE Analysis (Minimap2)
2.6. Analysis of Nad1 Intron
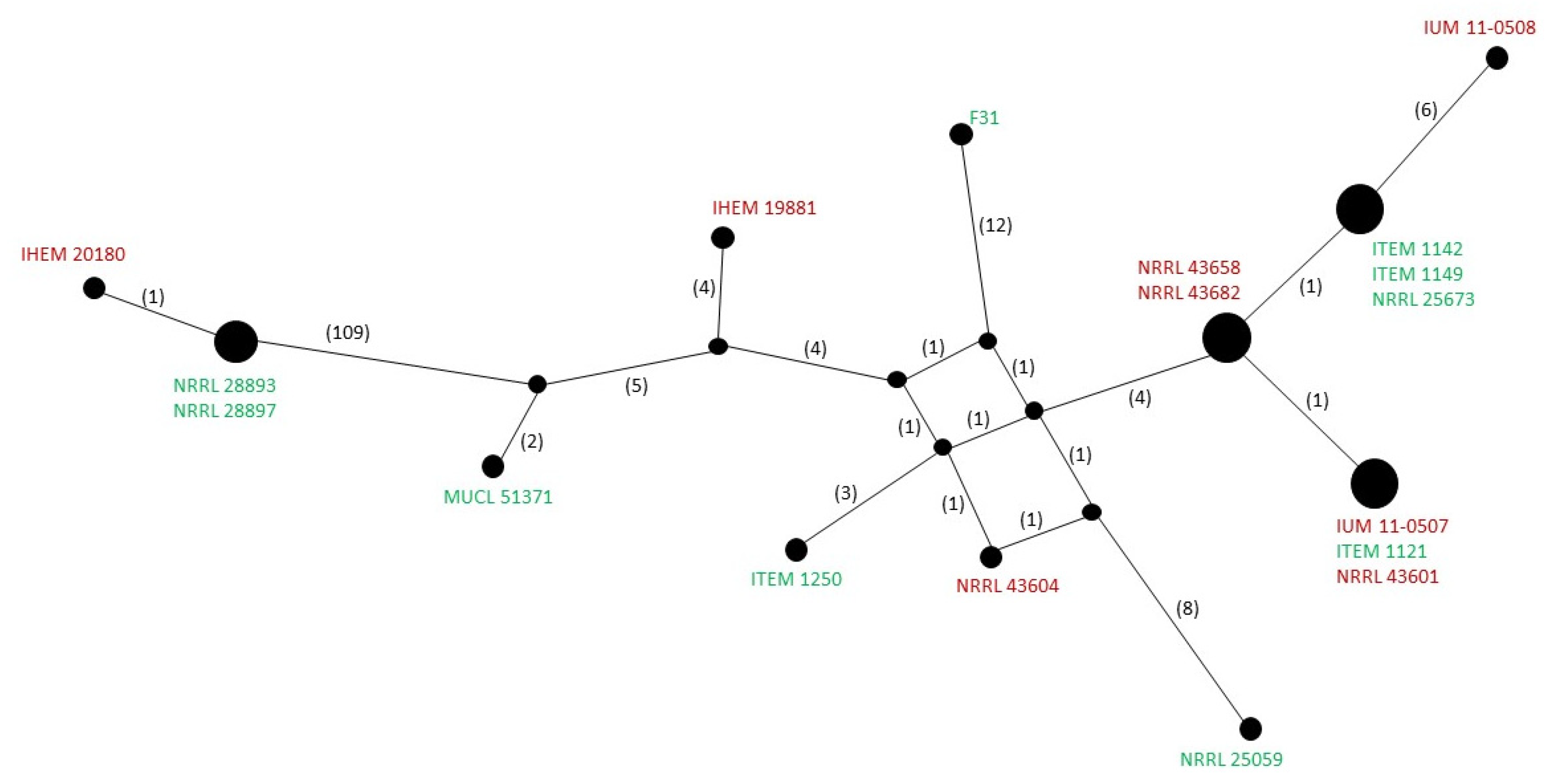
2.7. Haplotype Analysis of Mitogenomes
3. Results
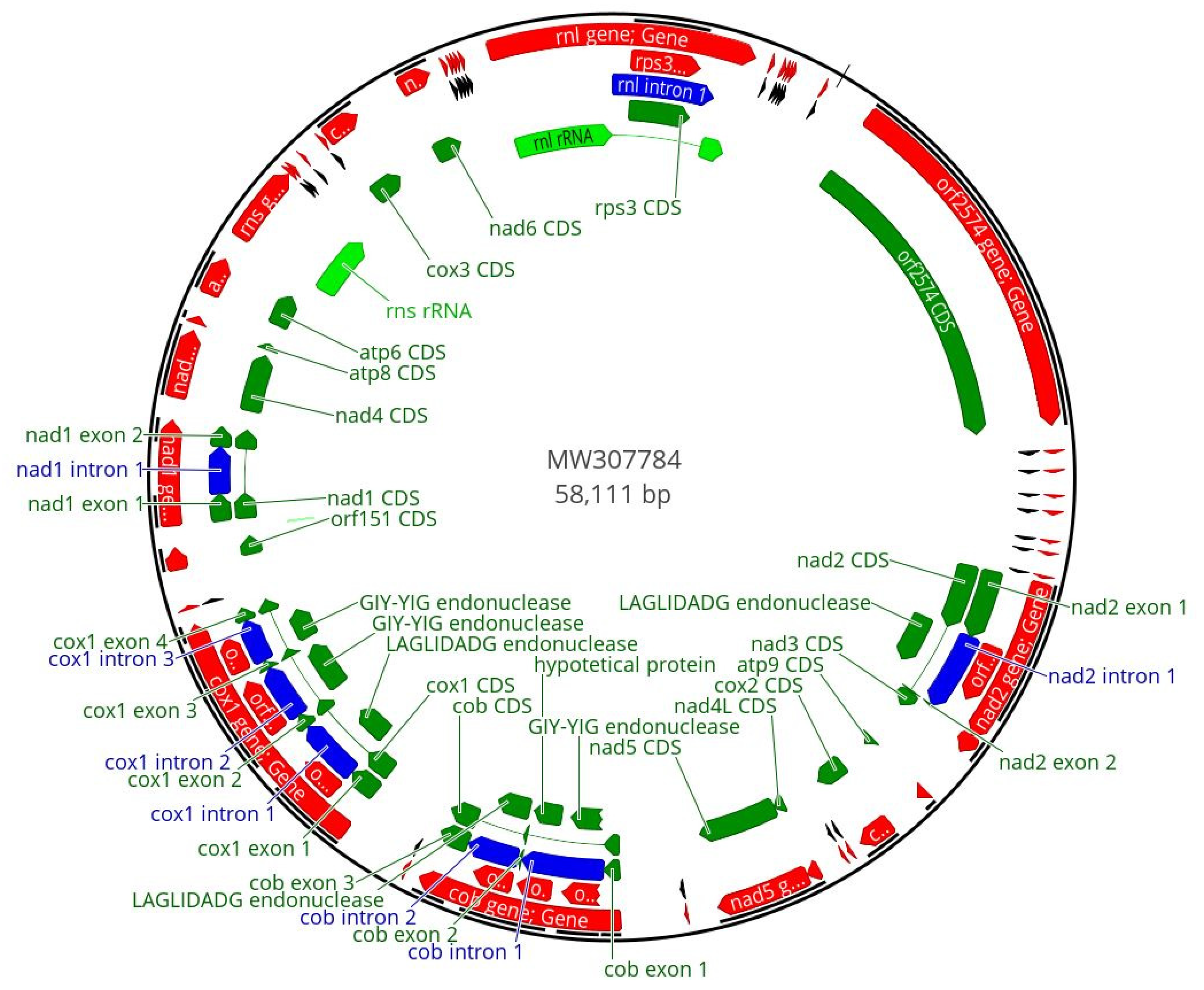
3.1. F. musae Mitogenomes
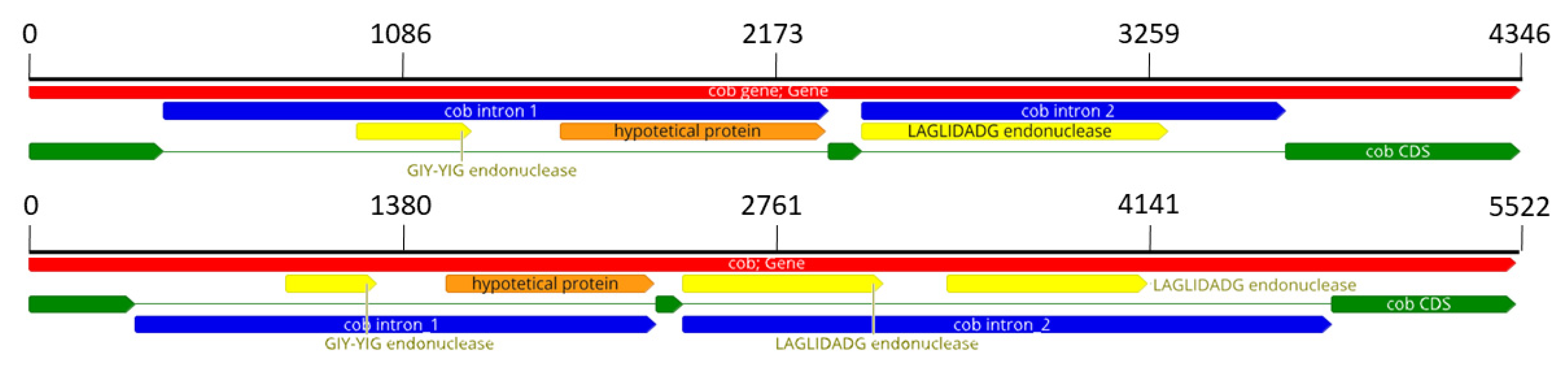
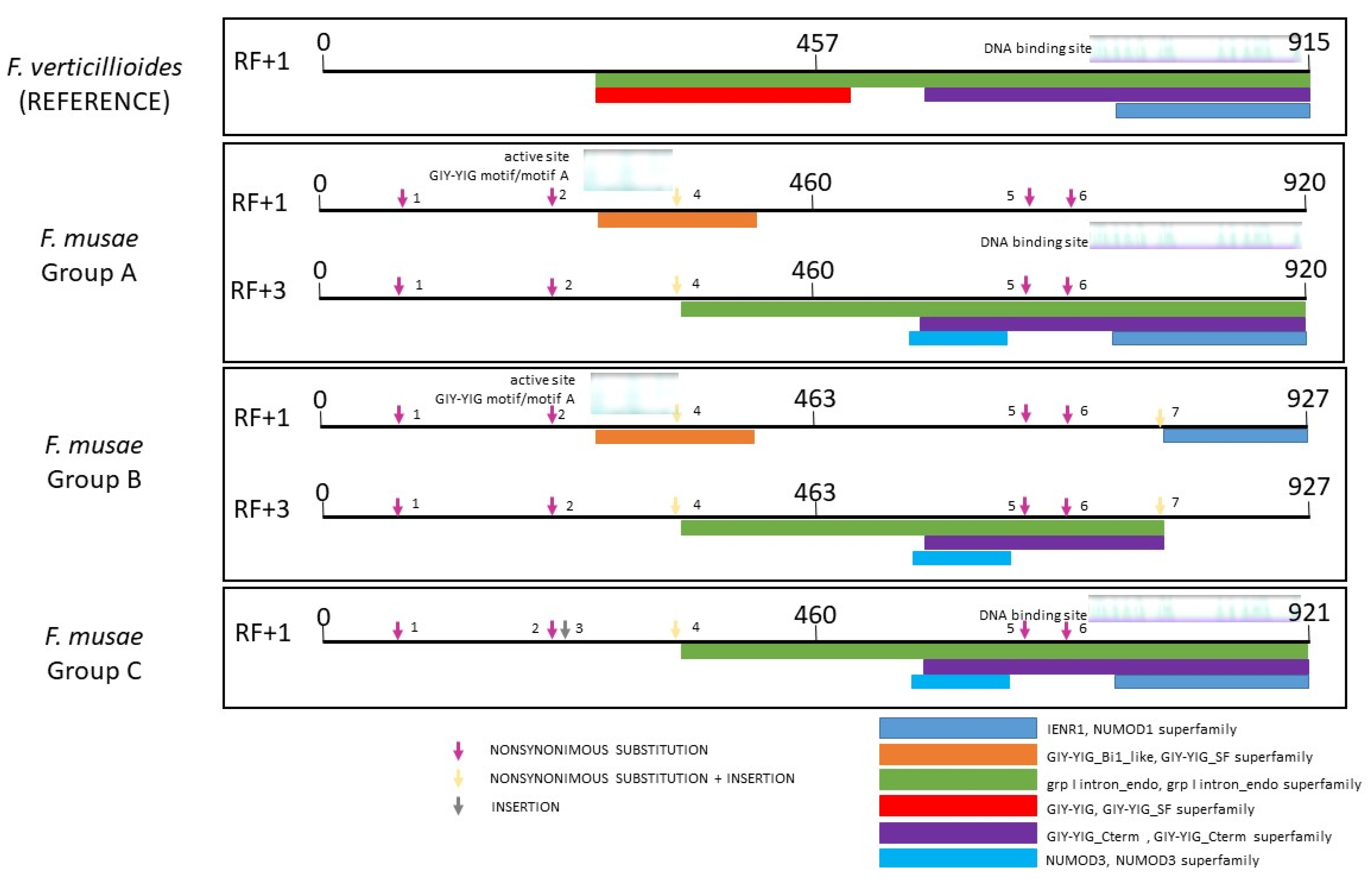
3.2. Diversity within F. musae
3.3. The Diagnostic Power of Mitochondrial Genomes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gauthier, G.M.; Keller, N.P. Crossover Fungal Pathogens: The Biology and Pathogenesis of Fungi Capable of Crossing Kingdoms to Infect Plants and Humans. Fungal Genet. Biol. 2013, 61, 146–157. [Google Scholar] [CrossRef] [PubMed]
- Hove, F.V.; Waalwijk, C.; Logrieco, A.; Munaut, F.; Moretti, A. Gibberella musae (Fusarium musae) Sp. Nov., a Recently Discovered Species from Banana Is Sister to F. verticillioides. Mycologia 2011, 103, 570–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamel, M.A.M.; Cortesi, P.; Saracchi, M. Etiological Agents of Crown Rot of Organic Bananas in Dominican Republic. Postharvest Biol. Technol. 2016, 120, 112–120. [Google Scholar] [CrossRef]
- Molnár, O.; Bartók, T.; Szécsi, Á. Occurrence of Fusarium verticillioides and Fusarium musae on Banana Fruits Marketed in Hungary. Acta Microbiol. Immunol. Hung. 2015, 62, 109–119. [Google Scholar] [CrossRef] [Green Version]
- Triest, D.; Stubbe, D.; De Cremer, K.; Piérard, D.; Detandt, M.; Hendrickx, M. Banana Infecting Fungus, Fusarium musae, Is Also an Opportunistic Human Pathogen: Are Bananas Potential Carriers and Source of Fusariosis? Mycologia 2015, 107, 46–53. [Google Scholar] [CrossRef]
- Verbeke, V.; Bourgeois, T.; Lodewyck, T.; Van Praet, J.; Lagrou, K.; Reynders, M.; Nulens, E. Successful Outcome of Disseminated Fusarium musae Fungemia with Skin Localization Treated with Liposomal Amphotericin B and Voriconazole in a Patient with Acute Myeloid Leukemia. Mycopathologia 2020, 185, 1085–1089. [Google Scholar] [CrossRef]
- Tava, V.; Prigitano, A.; Cortesi, P.; Esposto, M.C.; Pasquali, M. Fusarium musae from Diseased Bananas and Human Patients: Susceptibility to Fungicides Used in Clinical and Agricultural Settings. J. Fungi 2021, 7, 784. [Google Scholar] [CrossRef]
- Esposto, M.C.; Prigitano, A.; Tortorano, A.M. Fusarium musae as Cause of Superficial and Deep-Seated Human Infections. J. Mycol. Med. 2016, 26, 403–405. [Google Scholar] [CrossRef]
- Triest, D.; Hendrickx, M. Postharvest Disease of Banana Caused by Fusarium musae: A Public Health Concern? PLoS Pathog. 2016, 12, e1005940. [Google Scholar] [CrossRef] [Green Version]
- De Almeida, J.R.; Riaño Pachón, D.M.; Franceschini, L.M.; Dos Santos, I.B.; Ferrarezi, J.A.; de Andrade, P.A.M.; Monteiro-Vitorello, C.B.; Labate, C.A.; Quecine, M.C. Revealing the High Variability on Nonconserved Core and Mobile Elements of Austropuccinia psidii and Other Rust Mitochondrial Genomes. PLoS ONE 2021, 16, e0248054. [Google Scholar] [CrossRef]
- Wyrębek, J.; Molcan, T.; Myszczyński, K.; van Diepeningen, A.D.; Stakheev, A.A.; Żelechowski, M.; Bilska, K.; Kulik, T. Uncovering Diagnostic Value of Mitogenome for Identification of Cryptic Species Fusarium graminearum Sensu Stricto. Front. Microbiol. 2021, 12, 714651. [Google Scholar] [CrossRef] [PubMed]
- Kulik, T.; Bilska, K.; Żelechowski, M. Promising Perspectives for Detection, Identification, and Quantification of Plant Pathogenic Fungi and Oomycetes through Targeting Mitochondrial DNA. Int. J. Mol. Sci. 2020, 21, 2645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fonseca, P.L.C.; De-Paula, R.B.; Araújo, D.S.; Tomé, L.M.R.; Mendes-Pereira, T.; Rodrigues, W.F.C.; Del-Bem, L.-E.; Aguiar, E.R.G.R.; Góes-Neto, A. Global Characterization of Fungal Mitogenomes: New Insights on Genomic Diversity and Dynamism of Coding Genes and Accessory Elements. Front. Microbiol. 2021, 12, 787283. [Google Scholar] [CrossRef] [PubMed]
- Valenti, I.; Degradi, L.; Kunova, A.; Cortesi, P.; Pasquali, M.; Saracchi, M. The First Mitochondrial Genome of Ciborinia camelliae and Its Position in the Sclerotiniaceae Family. Front. Fungal Biol. 2022, 2, 802511. [Google Scholar] [CrossRef]
- Theelen, B.; Christinaki, A.C.; Dawson, T.L.; Boekhout, T.; Kouvelis, V.N. Comparative Analysis of Malassezia furfur Mitogenomes and the Development of a Mitochondria-Based Typing Approach. FEMS Yeast Res. 2021, 21, foab051. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Wu, H.; Geng, Y.; Li, Q.; Zang, R.; Guo, Y.; Xu, C.; Zhang, M. Mitogenome-Wide Comparison and Phylogeny Reveal Group I Intron Dynamics and Intraspecific Diversification within the Phytopathogen Corynespora cassiicola. Comput. Struct. Biotechnol. J. 2021, 19, 5987–5999. [Google Scholar] [CrossRef]
- Al-Reedy, R.M.; Malireddy, R.; Dillman, C.B.; Kennell, J.C. Comparative Analysis of Fusarium Mitochondrial Genomes Reveals a Highly Variable Region That Encodes an Exceptionally Large Open Reading Frame. Fungal Genet. Biol. 2012, 49, 2–14. [Google Scholar] [CrossRef]
- Hamari, Z.; Juhász, A.; Kevei, F. Role of Mobile Introns in Mitochondrial Genome Diversity of Fungi (a Mini Review). Acta Microbiol. Immunol. Hung. 2002, 49, 331–335. [Google Scholar] [CrossRef]
- Yang, M.; Zhang, H.; van der Lee, T.A.J.; Waalwijk, C.; van Diepeningen, A.D.; Feng, J.; Brankovics, B.; Chen, W. Population Genomic Analysis Reveals a Highly Conserved Mitochondrial Genome in Fusarium asiaticum. Front. Microbiol. 2020, 11, 839. [Google Scholar] [CrossRef]
- Pasquali, M.; Acquadro, A.; Balmas, V.; Migheli, Q.; Gullino, M.L.; Garibaldi, A. Development of PCR Primers for a New Fusarium oxysporum Pathogenic on Paris Daisy (Argyranthemum frutescens L.). Eur. J. Plant Pathol. 2004, 110, 7–11. [Google Scholar] [CrossRef]
- Brankovics, B.; van Diepeningen, A.D.; de Hoog, G.S.; van der Lee, T.A.J.; Waalwijk, C. Detecting Introgression Between Members of the Fusarium fujikuroi and F. oxysporum Species Complexes by Comparative Mitogenomics. Front. Microbiol. 2020, 11, 1092. [Google Scholar] [CrossRef] [PubMed]
- Degradi, L.; Tava, V.; Kunova, A.; Cortesi, P.; Saracchi, M.; Pasquali, M. Telomere to Telomere Genome Assembly of Fusarium musae F31, Causal Agent of Crown Rot Disease of Banana. Mol. Plant Microbe Interact. 2021, 34, 1455–1457. [Google Scholar] [CrossRef] [PubMed]
- Dierckxsens, N.; Mardulyn, P.; Smits, G. Unraveling Heteroplasmy Patterns with NOVOPlasty. NAR Genom. Bioinform. 2020, 2, lqz011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowe, T.M.; Chan, P.P. TRNAscan-SE On-Line: Integrating Search and Context for Analysis of Transfer RNA Genes. Nucleic Acids Res. 2016, 44, W54–W57. [Google Scholar] [CrossRef]
- Bandelt, H.J.; Forster, P.; Röhl, A. Median-Joining Networks for Inferring Intraspecific Phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef]
- Hartmann, F.E.; Snirc, A.; Cornille, A.; Godé, C.; Touzet, P.; Van Rossum, F.; Fournier, E.; Le Prieur, S.; Shykoff, J.; Giraud, T. Congruent Population Genetic Structures and Divergence Histories in Anther-Smut Fungi and Their Host Plants Silene italica and the Silene nutans Species Complex. Mol. Ecol. 2020, 29, 1154–1172. [Google Scholar] [CrossRef]
- Kulik, T.; Brankovics, B.; van Diepeningen, A.D.; Bilska, K.; Żelechowski, M.; Myszczyński, K.; Molcan, T.; Stakheev, A.; Stenglein, S.; Beyer, M.; et al. Diversity of Mobile Genetic Elements in the Mitogenomes of Closely Related Fusarium culmorum and F. graminearum Sensu Stricto Strains and Its Implication for Diagnostic Purposes. Front. Microbiol. 2020, 11, 1002. [Google Scholar] [CrossRef]
- Fourie, G.; Van der Merwe, N.A.; Wingfield, B.D.; Bogale, M.; Wingfield, M.J.; Steenkamp, E.T. Mitochondrial Introgression and Interspecies Recombination in the Fusarium fujikuroi Species Complex. IMA Fungus 2018, 9, 37–48. [Google Scholar] [CrossRef] [Green Version]
- Xiao, S.; Nguyen, D.T.; Wu, B.; Hao, W. Genetic Drift and Indel Mutation in the Evolution of Yeast Mitochondrial Genome Size. Genome Biol. Evol. 2017, 9, 3088–3099. [Google Scholar] [CrossRef] [Green Version]
- Sommerhalder, R.J.; McDonald, B.A.; Zhan, J. Concordant Evolution of Mitochondrial and Nuclear Genomes in the Wheat Pathogen Phaeosphaeria nodorum. Fungal Genet. Biol. 2007, 44, 764–772. [Google Scholar] [CrossRef]
- Misas, E.; Chow, N.A.; Gómez, O.M.; Muñoz, J.F.; McEwen, J.G.; Litvintseva, A.P.; Clay, O.K. Mitochondrial Genome Sequences of the Emerging Fungal Pathogen Candida auris. Front. Microbiol. 2020, 11, 560332. [Google Scholar] [CrossRef] [PubMed]
- Jelen, V.; de Jonge, R.; Van de Peer, Y.; Javornik, B.; Jakše, J. Complete Mitochondrial Genome of the Verticillium-Wilt Causing Plant Pathogen Verticillium nonalfalfae. PLoS ONE 2016, 11, e0148525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santamaria, M.; Vicario, S.; Pappadà, G.; Scioscia, G.; Scazzocchio, C.; Saccone, C. Towards Barcode Markers in Fungi: An Intron Map of Ascomycota Mitochondria. BMC Bioinform. 2009, 10, S15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fourie, G.; van der Merwe, N.A.; Wingfield, B.D.; Bogale, M.; Tudzynski, B.; Wingfield, M.J.; Steenkamp, E.T. Evidence for Inter-Specific Recombination among the Mitochondrial Genomes of Fusarium Species in the Gibberella fujikuroi Complex. BMC Genom. 2013, 14, 605. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| F. musae Strain/Feature | IUM 110508 | F31 | NRRL 43682 | NRRL 43658 | NRRL 43604 | NRRL 43601 | NRRL 28897 | NRRL 28893 | NRRL 25673 | MUCL 51371 | IUM 11-0507 | ITEM 1250 | ITEM 1149 | ITEM 1142 | ITEM 1121 | IHEM 20180 | IHEM 19881 | NRRL 25059 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| mtDNA | MW 307784 | MW 296866 | ON 240679 | ON 240982 | ON 240983 | ON 240980 | ON 240981 | ON 240987 | ON 240984 | ON 240985 | ON 240992 | ON 240991 | ON 240990 | ON 240989 | ON 240988 | ON 240986 | ON 012582 | MT 010916 |
| Length (bp) | 58,111 | 58,072 | 58,076 | 58,078 | 58,089 | 58,063 | 59,252 | 59,252 | 58,080 | 59,256 | 58,105 | 58,075 | 56,493 | 58,108 | 58,077 | 59,249 | 58,093 | 58,099 |
| Identity (%) | REF | 99.8 | 99.9 | 99.9 | 99.9 | 99.9 | 99.5 | 99.5 | 99.9 | 99.9 | 99.97 | 99.9 | 99.9 | 99.98 | 99.9 | 99.5 | 99.9 | 99.8 |
| Protein genes * | ||||||||||||||||||
| Number | 15 | 15 | 15 | 15 | 15 | 15 | 15 | 15 | 15 | 15 | 15 | 15 | 15 | 15 | 15 | 15 | 15 | 15 |
| Length (bp) | 14,148 | 14,148 | 14,148 | 14,148 | 14,148 | 14,148 | 14,148 | 14,148 | 14,148 | 14,148 | 14,148 | 14,148 | 14,148 | 14,148 | 14,148 | 14,148 | 14,148 | 14,148 |
| Identity (%) | REF | 99.96 | 100 | 100 | 100 | 100 | 99.99 | 99.99 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 99.99 | 100 | 100 |
| Differences | REF | 2 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 |
| Transversion | REF | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| MGEs | ||||||||||||||||||
| Number ** | 6 + 1 | 6 + 1 | 6 + 1 | 6 + 1 | 6 + 1 | 6 + 1 | 7 + 1 | 7 + 1 | 6 + 1 | 7 + 1 | 6 + 1 | 6 + 1 | 6 + 1 | 6 + 1 | 6 + 1 | 7 + 1 | 6 + 1 | 6 + 1 |
| Length (bp) | 7009 | 7010 | 7009 | 7009 | 7009 | 7009 | 7464 | 7464 | 7009 | 7448 | 7009 | 7009 | 7009 | 7009 | 7009 | 7464 | 7009 | 7017 |
| Identity (%) | REF | 100% | 100% | 100% | 100% | 100% | -- *** | -- *** | 100% | -- *** | 100% | 100% | 100% | 100% | 100% | -- *** | 100% | 99.80% |
| ORF | ||||||||||||||||||
| Number | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 |
| Length (bp) | 8952 | 8952 | 8952 | 8952 | 8952 | 8952 | 8949 | 8949 | 8952 | 8952 | 8952 | 8955 | 7377 | 8952 | 8952 | 8949 | 8952 | 8952 |
| Identitcal sites | REF | 8948 | 8945 | 8945 | 8945 | 8944 | 8931 | 8931 | 8946 | 8943 | 8944 | 8945 | 7369 | 8946 | 8944 | 8931 | 8943 | 8945 |
| rns + tRNA | ||||||||||||||||||
| Length (bp) | 1668 + 2005 | 1668 + 2005 | 1668 + 2005 | 1668 + 2005 | 1668 + 2005 | 1668 + 2005 | 1668 + 2005 | 1668 + 2005 | 1668 + 2005 | 1668 + 2005 | 1668 + 2005 | 1668 + 2005 | 1668 + 2005 | 1668 + 2005 | 1668 + 2005 | 1668 + 2005 | 1668 + 2005 | 1668 + 2005 |
| Identity (%) | REF | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Degradi, L.; Tava, V.; Prigitano, A.; Esposto, M.C.; Tortorano, A.M.; Saracchi, M.; Kunova, A.; Cortesi, P.; Pasquali, M. Exploring Mitogenomes Diversity of Fusarium musae from Banana Fruits and Human Patients. Microorganisms 2022, 10, 1115. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10061115
Degradi L, Tava V, Prigitano A, Esposto MC, Tortorano AM, Saracchi M, Kunova A, Cortesi P, Pasquali M. Exploring Mitogenomes Diversity of Fusarium musae from Banana Fruits and Human Patients. Microorganisms. 2022; 10(6):1115. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10061115
Chicago/Turabian StyleDegradi, Luca, Valeria Tava, Anna Prigitano, Maria Carmela Esposto, Anna Maria Tortorano, Marco Saracchi, Andrea Kunova, Paolo Cortesi, and Matias Pasquali. 2022. "Exploring Mitogenomes Diversity of Fusarium musae from Banana Fruits and Human Patients" Microorganisms 10, no. 6: 1115. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10061115