
Escherichia/Shigella, SCFAs, and Metabolic Pathways—The Triad That Orchestrates Intestinal Dysbiosis in Patients with Decompensated Alcoholic Cirrhosis from Western Mexico

 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Approval of Clinical Research
2.2. Study Design
2.3. Extraction of Nucleic Acids and 16S rRNA Amplicon Sequencing
2.4. Bioinformatic Analysis of 16S Amplicon Sequencing
2.5. Analysis of Fecal SCFAs
2.6. Statistical Analyses
3. Results
3.1. Cross-Sectional Study and Clinical Assessments
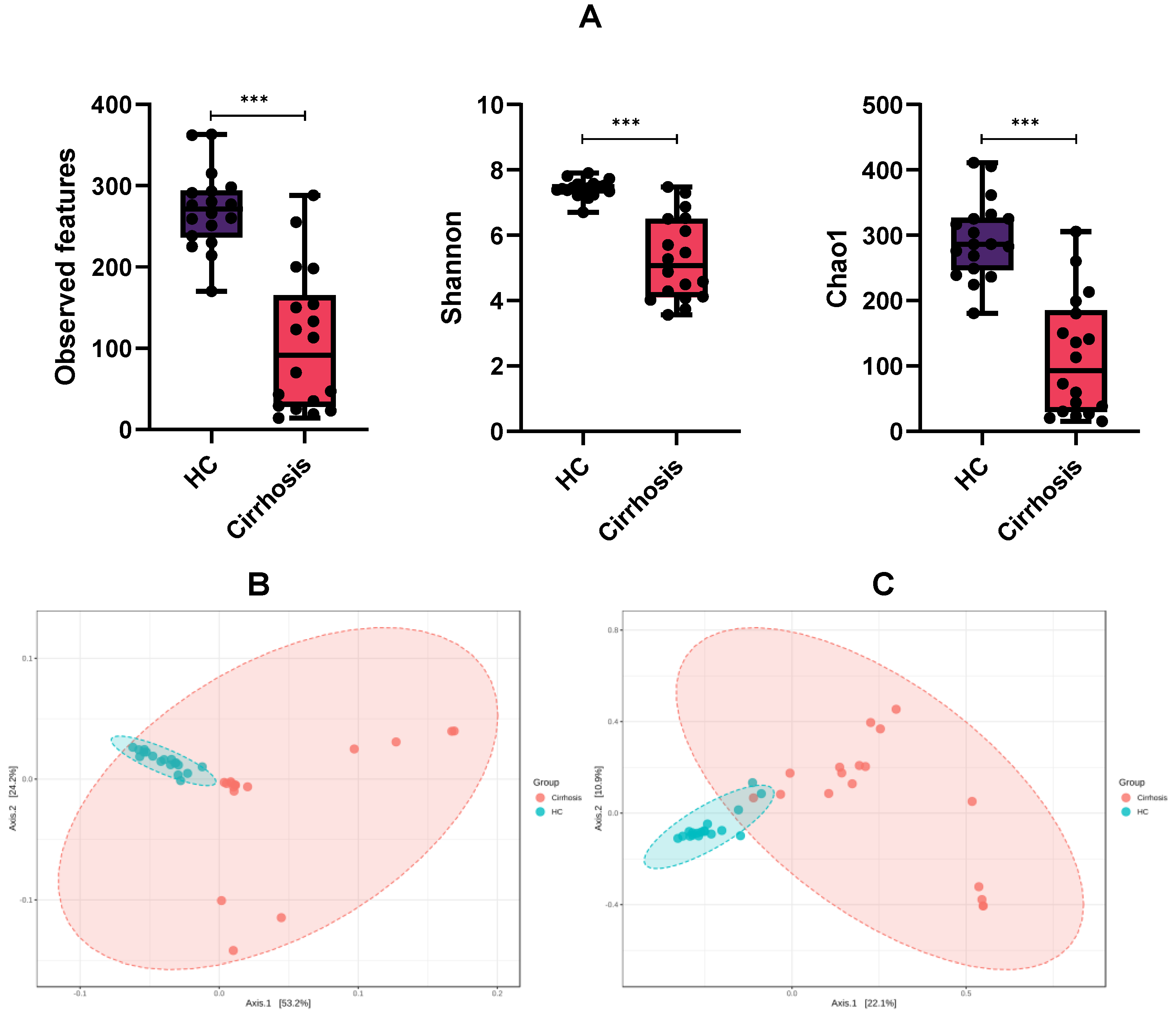
3.2. Microbiota Diversity between Groups
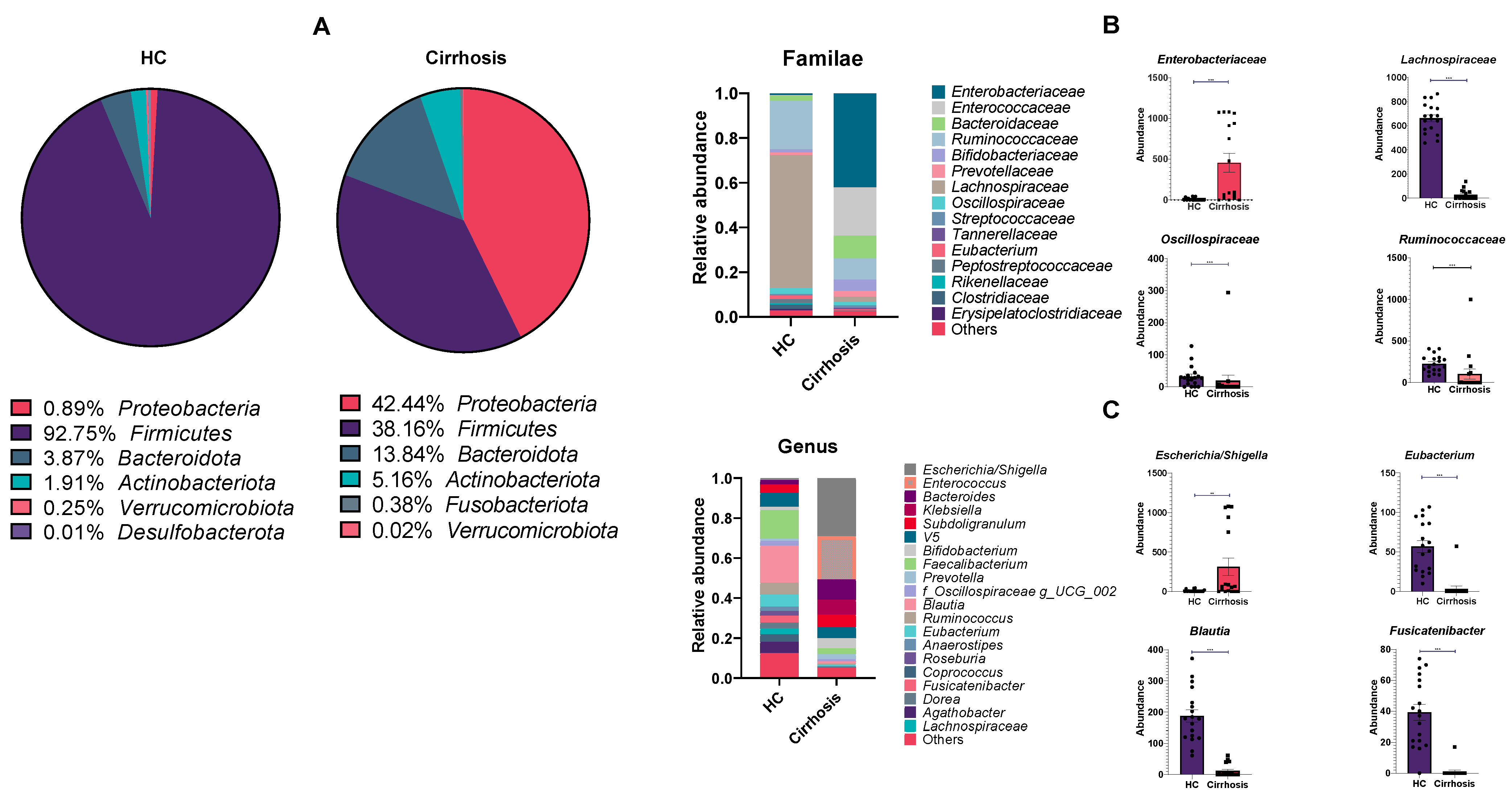
3.3. Detected ASVs across Different Taxonomic Levels
3.4. Linear Discriminant Analysis Effect Size (LEfSe)
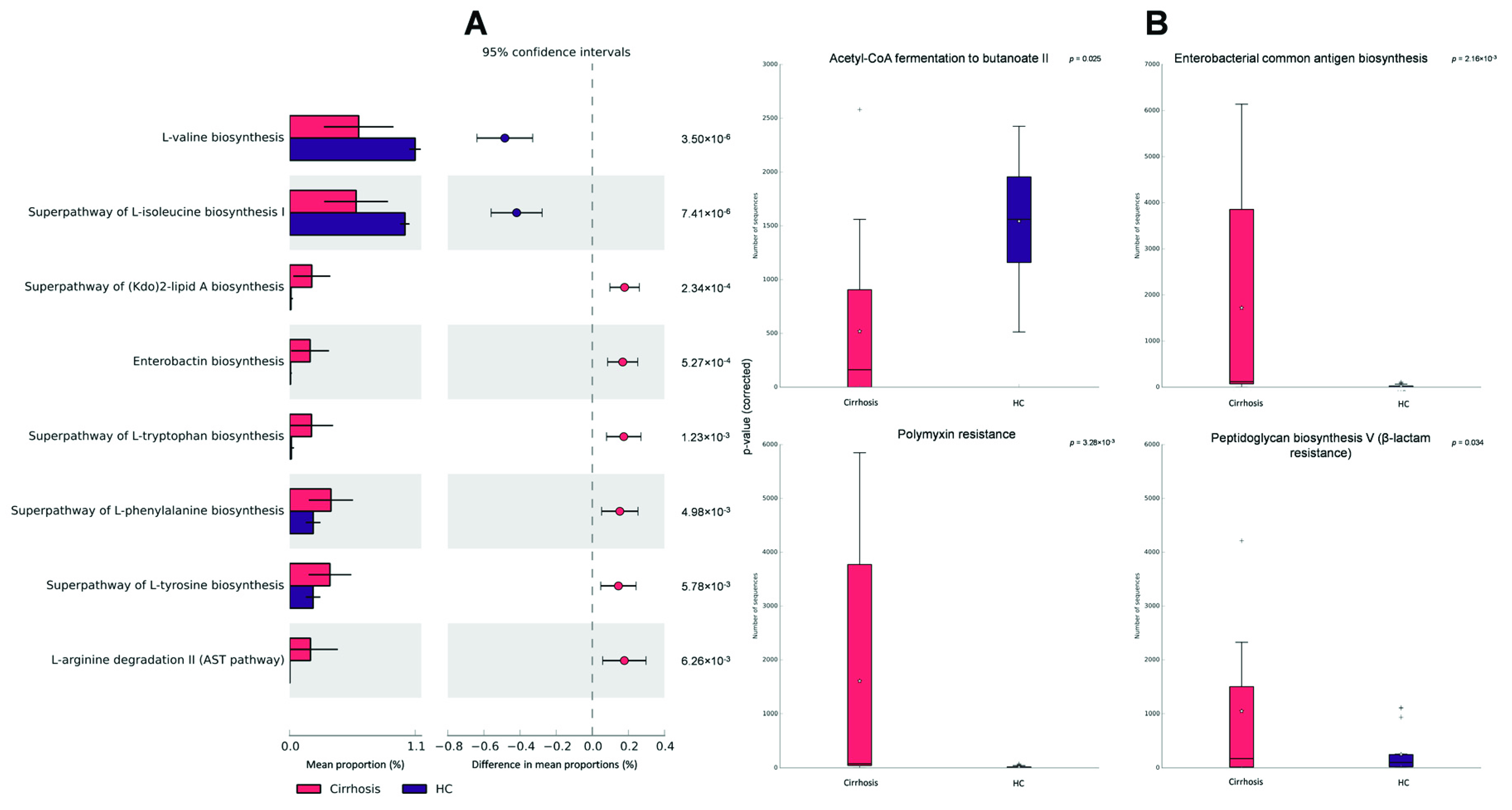
3.5. Functional Metagenomic Profiles
3.6. Compositional Methods (ANCOM)
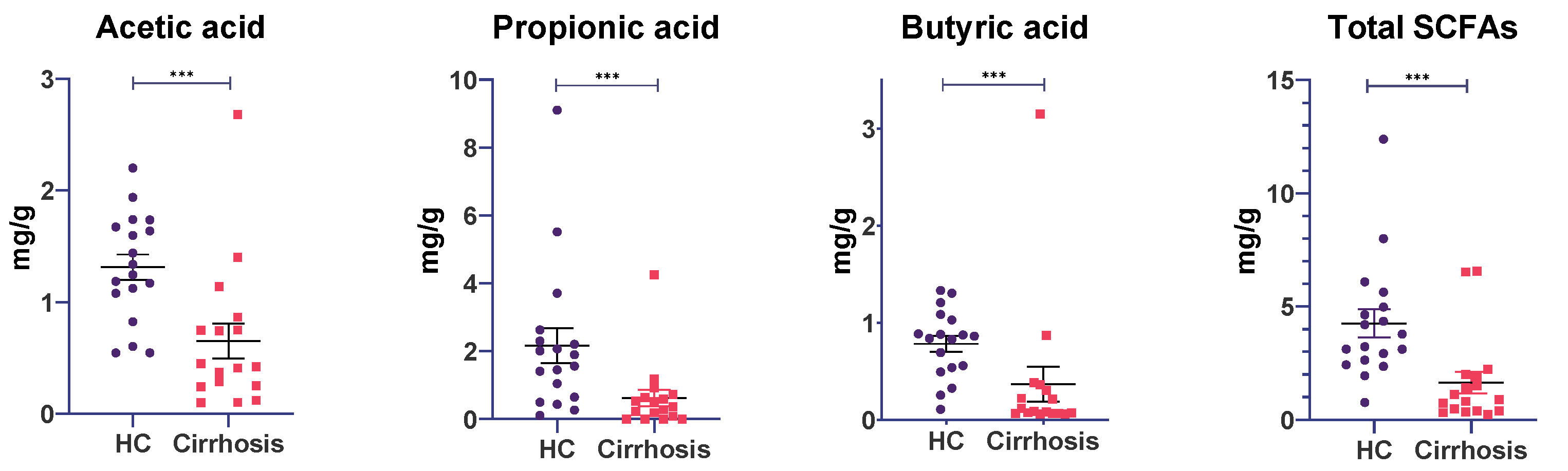
3.7. Fecal Short-Chain Fatty Acid Concentrations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- INEGI. Características de Las Defunciones Registradas En México Durante 2020; INEGI: Mexico City, Mexico, 2021.
- Bajaj, J.S.; Heuman, D.M.; Hylemon, P.B.; Sanyal, A.J.; White, M.B.; Monteith, P.; Noble, N.A.; Unser, A.B.; Daita, K.; Fisher, A.R.; et al. Altered Profile of Human Gut Microbiome Is Associated with Cirrhosis and Its Complications. J. Hepatol. 2014, 60, 940–947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gagliardi, A.; Totino, V.; Cacciotti, F.; Iebba, V.; Neroni, B.; Bonfiglio, G.; Trancassini, M.; Passariello, C.; Pantanella, F.; Schippa, S. Rebuilding the Gut Microbiota Ecosystem. Int. J. Environ. Res. Public Health 2018, 15, 1679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bajaj, J.S.; Torre, A.; Rojas, M.L.; Fagan, A.; Nandez, I.E.; Gavis, E.A.; De Leon Osorio, O.; White, M.B.; Fuchs, M.; Sikaroodi, M.; et al. Cognition and Hospitalizations Are Linked with Salivary and Faecal Microbiota in Cirrhosis Cohorts from the USA and Mexico. Liver Int. 2020, 40, 1395–1407. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Yang, F.; Lu, H.; Wang, B.; Chen, Y.; Lei, D.; Wang, Y.; Zhu, B.; Li, L. Characterization of Fecal Microbial Communities in Patients with Liver Cirrhosis. Hepatology 2011, 54, 562–572. [Google Scholar] [CrossRef] [PubMed]
- Wiest, R.; Lawson, M.; Geuking, M. Pathological Bacterial Translocation in Liver Cirrhosis. J. Hepatol. 2014, 60, 197–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, N.; Yang, F.; Li, A.; Prifti, E.; Chen, Y.; Shao, L.; Guo, J.; Le Chatelier, E.; Yao, J.; Wu, L.; et al. Alterations of the Human Gut Microbiome in Liver Cirrhosis. Nature 2014, 513, 59–64. [Google Scholar] [CrossRef]
- Ramos-Lopez, O. Genetic, Metabolic and Environmental Factors Involved in the Development of Liver Cirrhosis in Mexico. World J. Gastroenterol. 2015, 21, 11552. [Google Scholar] [CrossRef]
- Cummings, J.H.; Pomare, E.W.; Branch, W.J.; Naylor, C.P.; Macfarlane, G.T. Short Chain Fatty Acids in Human Large Intestine, Portal, Hepatic and Venous Blood. Gut 1987, 28, 1221–1227. [Google Scholar] [CrossRef] [Green Version]
- Den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.-J.; Bakker, B.M. The Role of Short-Chain Fatty Acids in the Interplay between Diet, Gut Microbiota, and Host Energy Metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [Green Version]
- Bloemen, J.G.; Venema, K.; van de Poll, M.C.; Olde Damink, S.W.; Buurman, W.A.; Dejong, C.H. Short Chain Fatty Acids Exchange across the Gut and Liver in Humans Measured at Surgery. Clin. Nutr. 2009, 28, 657–661. [Google Scholar] [CrossRef]
- Stumpff, F. A Look at the Smelly Side of Physiology: Transport of Short Chain Fatty Acids. Pflugers Arch. Eur. J. Physiol. 2018, 470, 571–598. [Google Scholar] [CrossRef] [PubMed]
- Candido, E.P.M.; Reeves, R.; Davie, J.R. Sodium Butyrate Inhibits Histone Deacetylation in Cultured Cells. Cell 1978, 14, 105–113. [Google Scholar] [CrossRef]
- Segain, J.P.; Galmiche, J.P.; Raingeard De La Blétière, D.; Bourreille, A.; Leray, V.; Gervois, N.; Rosales, C.; Ferrier, L.; Bonnet, C.; Blottière, H.M. Butyrate Inhibits Inflammatory Responses through NFκB Inhibition: Implications for Crohn’s Disease. Gut 2000, 47, 397–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, R.; De Zoeten, E.F.; Özkaynak, E.; Chen, C.; Wang, L.; Porrett, P.M.; Li, B.; Turka, L.A.; Olson, E.N.; Greene, M.I.; et al. Deacetylase Inhibition Promotes the Generation and Function of Regulatory T Cells. Nat. Med. 2007, 13, 1299–1307. [Google Scholar] [CrossRef] [PubMed]
- Dou, X.; Gao, N.; Yan, D.; Shan, A. Sodium Butyrate Alleviates Mouse Colitis by Regulating Gut Microbiota Dysbiosis. Animals 2020, 10, 1154. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.X.; Lee, J.S.; Campbell, E.L.; Colgan, S.P. Microbiota-Derived Butyrate Dynamically Regulates Intestinal Homeostasis through Regulation of Actin-Associated Protein Synaptopodin. Proc. Natl. Acad. Sci. USA 2020, 117, 11648–11657. [Google Scholar] [CrossRef]
- Peña-Rodríguez, M.; Vega-Magaña, N.; García-Benavides, L.; Zepeda-Nuño, J.S.; Gutierrez-Silerio, G.Y.; González-Hernández, L.A.; Andrade-Villanueva, J.F.; Del Toro-Arreola, S.; Pereira-Suárez, A.L.; Bueno-Topete, M.R. Butyrate Administration Strengthens the Intestinal Epithelium and Improves Intestinal Dysbiosis in a Cholestasis Fibrosis Model. J. Appl. Microbiol. 2022, 132, 571–583. [Google Scholar] [CrossRef]
- Jin, M.; Kalainy, S.; Baskota, N.; Chiang, D.; Deehan, E.C.; McDougall, C.; Tandon, P.; Martínez, I.; Cervera, C.; Walter, J.; et al. Faecal Microbiota from Patients with Cirrhosis Has a Low Capacity to Ferment Non-digestible Carbohydrates into Short-chain Fatty Acids. Liver Int. 2019, 39, 1437–1447. [Google Scholar] [CrossRef]
- Illumina Illumina 16S Metagenomic Sequencing Library. Available online: https://support.illumina.com/documents/documentation/chemistry_documentation/16s/16s-metagenomic-library-prep-guide-15044223-b.pdf (accessed on 1 November 2020).
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, Interactive, Scalable and Extensible Microbiome Data Science Using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Misawa, K.; Kuma, K.I.; Miyata, T. MAFFT: A Novel Method for Rapid Multiple Sequence Alignment Based on Fast Fourier Transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately Maximum-Likelihood Trees for Large Alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef] [PubMed]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Gregory Caporaso, J. Optimizing Taxonomic Classification of Marker-Gene Amplicon Sequences with QIIME 2′s Q2-Feature-Classifier Plugin. Microbiome 2018, 6, 90. [Google Scholar] [CrossRef] [PubMed]
- Michael, S.R.; O’Rourke, D.R.; Kaehler, B.D.; Ziemski, M.; Dillon, M.R.; Foster, J.T.; Bokulich, N.A. RESCRIPt: Reproducible Sequence Taxonomy Reference Database Management for the Masses. bioRxiv 2020, 4, 1–9. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Chao, A. Nonparametric Estimation of the Number of Classes in a Population. Scand. J. Stat. 1984, 11, 265–270. [Google Scholar]
- Lozupone, C.A.; Hamady, M.; Kelley, S.T.; Knight, R. Quantitative and Qualitative β Diversity Measures Lead to Different Insights into Factors That Structure Microbial Communities. Appl. Environ. Microbiol. 2007, 73, 1576–1585. [Google Scholar] [CrossRef] [Green Version]
- Lozupone, C.; Knight, R. UniFrac: A New Phylogenetic Method for Comparing Microbial Communities. Appl. Environ. Microbiol. 2005, 71, 8228–8235. [Google Scholar] [CrossRef] [Green Version]
- Dhariwal, A.; Chong, J.; Habib, S.; King, I.L.; Agellon, L.B.; Xia, J. MicrobiomeAnalyst: A Web-Based Tool for Comprehensive Statistical, Visual and Meta-Analysis of Microbiome Data. Nucleic Acids Res. 2017, 45, W180–W188. [Google Scholar] [CrossRef]
- Chong, J.; Liu, P.; Zhou, G.; Xia, J. Using MicrobiomeAnalyst for Comprehensive Statistical, Functional, and Meta-Analysis of Microbiome Data. Nat. Protoc. 2020, 15, 799–821. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic Biomarker Discovery and Explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afgan, E.; Baker, D.; Batut, B.; van den Beek, M.; Bouvier, D.; Čech, M.; Chilton, J.; Clements, D.; Coraor, N.; Grüning, B.A.; et al. The Galaxy Platform for Accessible, Reproducible and Collaborative Biomedical Analyses: 2018 Update. Nucleic Acids Res. 2018, 46, W537–W544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.R.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G.I. PICRUSt2 for Prediction of Metagenome Functions. Nat. Biotechnol. 2020, 38, 685–688. [Google Scholar] [CrossRef] [PubMed]
- Barbera, P.; Kozlov, A.M.; Czech, L.; Morel, B.; Darriba, D.; Flouri, T.; Stamatakis, A. EPA-Ng: Massively Parallel Evolutionary Placement of Genetic Sequences. Syst. Biol. 2019, 68, 365–369. [Google Scholar] [CrossRef]
- Czech, L.; Barbera, P.; Stamatakis, A. Genesis and Gappa: Processing, Analyzing and Visualizing Phylogenetic (Placement) Data. Bioinformatics 2020, 36, 3263–3265. [Google Scholar] [CrossRef] [Green Version]
- Louca, S.; Doebeli, M. Efficient Comparative Phylogenetics on Large Trees. Bioinformatics 2018, 34, 1053–1055. [Google Scholar] [CrossRef]
- Ye, Y.; Doak, T.G. A Parsimony Approach to Biological Pathway Reconstruction/Inference for Genomes and Metagenomes. PLoS Comput. Biol. 2009, 5, e1000465. [Google Scholar] [CrossRef] [Green Version]
- Caspi, R.; Altman, T.; Billington, R.; Dreher, K.; Foerster, H.; Fulcher, C.A.; Holland, T.A.; Keseler, I.M.; Kothari, A.; Kubo, A.; et al. The MetaCyc Database of Metabolic Pathways and Enzymes and the BioCyc Collection of Pathway/Genome Databases. Nucleic Acids Res. 2014, 42, D459–D471. [Google Scholar] [CrossRef] [Green Version]
- Mandal, S.; Van Treuren, W.; White, R.A.; Eggesbø, M.; Knight, R.; Peddada, S.D. Analysis of Composition of Microbiomes: A Novel Method for Studying Microbial Composition. Microb. Ecol. Health Dis. 2015, 26, 27663. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, W.; Vinolo, M.; Calixto, L.; Ferreira, C. Use of Gas Chromatography to Quantify Short Chain Fatty Acids in the Serum, Colonic Luminal Content and Feces of Mice. Bio-Protoc. 2018, 8, e3089. [Google Scholar] [CrossRef] [Green Version]
- Parks, D.H.; Tyson, G.W.; Hugenholtz, P.; Beiko, R.G. STAMP: Statistical Analysis of Taxonomic and Functional Profiles. Bioinformatics 2014, 30, 3123–3124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheemerla, S.; Balakrishnan, M. Global Epidemiology of Chronic Liver Disease. Clin. Liver Dis. 2021, 17, 365–370. [Google Scholar] [CrossRef] [PubMed]
- Dubinkina, V.B.; Tyakht, A.V.; Odintsova, V.Y.; Yarygin, K.S.; Kovarsky, B.A.; Pavlenko, A.V.; Ischenko, D.S.; Popenko, A.S.; Alexeev, D.G.; Taraskina, A.Y.; et al. Links of Gut Microbiota Composition with Alcohol Dependence Syndrome and Alcoholic Liver Disease. Microbiome 2017, 5, 141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bajaj, J.S.; Ridlon, J.M.; Hylemon, P.B.; Thacker, L.R.; Heuman, D.M.; Smith, S.; Sikaroodi, M.; Gillevet, P.M. Linkage of Gut Microbiome with Cognition in Hepatic Encephalopathy. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 302, 168–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bajaj, J.S.; Hylemon, P.B.; Ridlon, J.M.; Heuman, D.M.; Daita, K.; White, M.B.; Monteith, P.; Noble, N.A.; Sikaroodi, M.; Gillevet, P.M. Colonic Mucosal Microbiome Differs from Stool Microbiome in Cirrhosis and Hepatic Encephalopathy and Is Linked to Cognition and Inflammation. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 303, 675–685. [Google Scholar] [CrossRef]
- Bajaj, J.S.; Vargas, H.E.; Reddy, K.R.; Lai, J.C.; O’Leary, J.G.; Tandon, P.; Wong, F.; Mitrani, R.; White, M.B.; Kelly, M.; et al. Association Between Intestinal Microbiota Collected at Hospital Admission and Outcomes of Patients With Cirrhosis. Clin. Gastroenterol. Hepatol. 2019, 17, 756–765.e3. [Google Scholar] [CrossRef]
- Sung, C.M.; Chen, K.F.; Lin, Y.F.; Ke, H.M.; Huang, H.Y.; Gong, Y.N.; Tsai, W.S.; You, J.F.; Lu, M.J.; Cheng, H.T.; et al. Predicting Clinical Outcomes of Cirrhosis Patients With Hepatic Encephalopathy From the Fecal Microbiome. Cell. Mol. Gastroenterol. Hepatol. 2019, 8, 301–318.e2. [Google Scholar] [CrossRef] [Green Version]
- González-Navajas, J.M.; Francés, R.; Such, J. Bacterial DNA in Patients with Cirrhosis and Sterile Ascites: Its Role as a Marker of Bacterial Translocation and Prognosis Tool. Rev. Esp. Enferm. Dig. 2007, 99, 599–603. [Google Scholar] [CrossRef] [Green Version]
- Larsen, J.M. The Immune Response to Prevotella Bacteria in Chronic Inflammatory Disease. Immunology 2017, 151, 363–374. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Mao, B.; Gu, J.; Wu, J.; Cui, S.; Wang, G.; Zhao, J.; Zhang, H.; Chen, W. Blautia—A New Functional Genus with Potential Probiotic Properties? Gut Microbes 2021, 13, 1875796. [Google Scholar] [CrossRef]
- Smith, A.J.; Jackson, M.S.; Bagg, J. The Ecology of Staphylococcus Species in the Oral Cavity. J. Med. Microbiol. 2001, 50, 940–946. [Google Scholar] [CrossRef] [PubMed]
- Bajaj, J.S.; Betrapally, N.S.; Hylemon, P.B.; Heuman, D.M.; Daita, K.; White, M.B.; Unser, A.; Thacker, L.R.; Sanyal, A.J.; Kang, D.J.; et al. Salivary Microbiota Reflects Changes in Gut Microbiota in Cirrhosis with Hepatic Encephalopathy. Hepatology 2015, 62, 1260–1271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, T.; Long, W.; Zhang, C.; Liu, S.; Zhao, L.; Hamaker, B.R. Fiber-Utilizing Capacity Varies in Prevotella- versus Bacteroides-Dominated Gut Microbiota. Sci. Rep. 2017, 7, 2594. [Google Scholar] [CrossRef] [PubMed]
- Trebicka, J.; Bork, P.; Krag, A.; Arumugam, M. Utilizing the Gut Microbiome in Decompensated Cirrhosis and Acute-on-Chronic Liver Failure. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 167–180. [Google Scholar] [CrossRef] [PubMed]
- Erridge, C.; Pridmore, A.; Eley, A.; Stewart, J.; Poxton, I.R. Lipopolysaccharides of Bacteroides Fragilis, Chlamydia Trachomatis and Pseudomonas Aeruginosa Signal via Toll-like Receptor 2. J. Med. Microbiol. 2004, 53, 735–740. [Google Scholar] [CrossRef] [PubMed]
- Lebeer, S.; Vanderleyden, J.; De Keersmaecker, S.C.J. Host Interactions of Probiotic Bacterial Surface Molecules: Comparison with Commensals and Pathogens. Nat. Rev. Microbiol. 2010, 8, 171–184. [Google Scholar] [CrossRef]
- Rosero, J.A.; Killer, J.; Sechovcová, H.; Mrázek, J.; Benada, O.; Fliegerová, K.; Havlík, J.; Kopečný, J. Reclassification of Eubacterium Rectale (Hauduroy et Al. 1937) Prévot 1938 in a New Genus Agathobacter Gen. Nov. as Agathobacter Rectalis Comb. Nov., and Description of Agathobacter Ruminis Sp. Nov., Isolated from the Rumen Contents of Sheep and Cows. Int. J. Syst. Evol. Microbiol. 2016, 66, 768–773. [Google Scholar] [CrossRef]
- Engels, C.; Ruscheweyh, H.J.; Beerenwinkel, N.; Lacroix, C.; Schwab, C. The Common Gut Microbe Eubacterium Hallii Also Contributes to Intestinal Propionate Formation. Front. Microbiol. 2016, 7, 713. [Google Scholar] [CrossRef] [Green Version]
- Takeshita, K.; Mizuno, S.; Mikami, Y.; Sujino, T.; Saigusa, K.; Matsuoka, K.; Naganuma, M.; Sato, T.; Takada, T.; Tsuji, H.; et al. A Single Species of Clostridium Subcluster XIVa Decreased in Ulcerative Colitis Patients. Inflamm. Bowel Dis. 2016, 22, 2802–2810. [Google Scholar] [CrossRef] [Green Version]
- Rivera-Chávez, F.; Lopez, C.A.; Bäumler, A.J. Oxygen as a Driver of Gut Dysbiosis. Free Radic. Biol. Med. 2017, 105, 93–101. [Google Scholar] [CrossRef]
- Kelly, C.J.; Zheng, L.; Campbell, E.L.; Saeedi, B.; Scholz, C.C.; Bayless, A.J.; Wilson, K.E.; Glover, L.E.; Kominsky, D.J.; Magnuson, A.; et al. Crosstalk between Microbiota-Derived Short-Chain Fatty Acids and Intestinal Epithelial HIF Augments Tissue Barrier Function. Cell Host Microbe 2015, 17, 662–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shelton, C.D.; Byndloss, M.X. Gut Epithelial Metabolism as a Key Driver of Intestinal Dysbiosis Associated with Noncommunicable Diseases. Infect. Immun. 2020, 88, e00939-19. [Google Scholar] [CrossRef] [PubMed]
- Nie, K.; Ma, K.; Luo, W.; Shen, Z.; Yang, Z.; Xiao, M.; Tong, T.; Yang, Y.; Wang, X. Roseburia Intestinalis: A Beneficial Gut Organism From the Discoveries in Genus and Species. Front. Cell. Infect. Microbiol. 2021, 11, 757718. [Google Scholar] [CrossRef] [PubMed]
- Winter, S.E.; Winter, M.G.; Xavier, M.N.; Thiennimitr, P.; Poon, V.; Keestra, A.M.; Laughlin, R.C.; Gomez, G.; Wu, J.; Lawhon, S.D.; et al. Host-Derived Nitrate Boosts Growth of E. Coli in the Inflamed Gut. Science 2013, 339, 708–711. [Google Scholar] [CrossRef] [Green Version]
- Guarner, C.; Soriano, G.; Tomas, A.; Bulbena, O.; Novella, M.T.; Balanzo, J.; Vilardell, F.; Mourelle, M.; Moncada, S. Increased Serum Nitrite and Nitrate Levels in Patients with Cirrhosis: Relationship to Endotoxemia. Hepatology 1993, 18, 1139–1143. [Google Scholar] [CrossRef]
- Shawcross, D.L.; Davies, N.A.; Williams, R.; Jalan, R. Systemic Inflammatory Response Exacerbates the Neuropsychological Effects of Induced Hyperammonemia in Cirrhosis. J. Hepatol. 2004, 40, 247–254. [Google Scholar] [CrossRef] [Green Version]
- Iwakiri, Y.; Groszmann, R.J. Vascular Endothelial Dysfunction in Cirrhosis. J. Hepatol. 2007, 46, 927–934. [Google Scholar] [CrossRef]
- Wang, L.; Fouts, D.E.; Stärkel, P.; Hartmann, P.; Chen, P.; Llorente, C.; DePew, J.; Moncera, K.; Ho, S.B.; Brenner, D.A.; et al. Intestinal REG3 Lectins Protect against Alcoholic Steatohepatitis by Reducing Mucosa-Associated Microbiota and Preventing Bacterial Translocation. Cell Host Microbe 2016, 19, 227–239. [Google Scholar] [CrossRef] [Green Version]
- Erbel, P.J.A.; Barr, K.; Gao, N.; Gerwig, G.J.; Rick, P.D.; Gardner, K.H. Identification and Biosynthesis of Cyclic Enterobacterial Common Antigen in Escherichia Coli. J. Bacteriol. 2003, 185, 1995–2004. [Google Scholar] [CrossRef] [Green Version]
- Raymond, K.N.; Dertz, E.A.; Kim, S.S. Enterobactin: An Archetype for Microbial Iron Transport. Proc. Natl. Acad. Sci. USA 2003, 100, 3584–3588. [Google Scholar] [CrossRef] [Green Version]
- Fernández, J.; Tandon, P.; Mensa, J.; Garcia-Tsao, G. Antibiotic Prophylaxis in Cirrhosis: Good and Bad. Hepatology 2016, 63, 2019–2031. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.C.; Hung, C.H.; Chen, Y.S.; Cheng, J.S.; Lee, S.S.J.; Tseng, F.C.; Cheng, M.F.; Wang, J.L. Bloodstream Infections Caused by Extended-Spectrum Beta-Lactamase-Producing Escherichia Coli in Patients with Liver Cirrhosis. Pathogens 2021, 10, 37. [Google Scholar] [CrossRef] [PubMed]
- Luján-Ramos, M.A.; Díaz-Ramírez, G.S.; Martínez-Casas, O.Y.; Morales-Ortiz, A.F.; Donado-Gómez, J.H.; Restrepo-Gutiérrez, J.C.; Marín-Zuluaga, J.I. Characterization of Patients with Liver Cirrhosis and Bacteremia Treated at a University Hospital in Medellín-Colombia. Rev. Colomb. Gastroenterol. 2020, 35, 455–464. [Google Scholar] [CrossRef]
- Velarde-Ruiz Velasco, J.A.; Lazcano-Becerra, M.; Tapia-Calderón, D.K.; Mercado-Jáuregui, L.A.; Alonzo-García, C.J.; González-Ruiz, R.; Morel-Cerda, E.C.; Barajas-Correa, D.I.; Aldana-Ledesma, J.M.; Filio-Hermosillo, A.C.; et al. Aislamientos Microbiológicos En Pacientes Hospitalizados Con Cirrosis Hepática. Rev. Méd. MD 2019, 10, 89–94. [Google Scholar]
- Fam, N.S.; Defasque, S.; Bert, F.; Leflon-Guibout, V.; El-Ray, A.; El-Ghannam, M.; Attia, M.E.; Omar, M.; Desouki, D.G.; Valla, D.; et al. Faecal Carriage of Extended-Spectrum β-Lactamase (ESBL)-Producing Enterobacteria in Liver Disease Patients from Two Hospitals in Egypt and France: A Comparative Epidemiological Study. Epidemiol. Infect. 2015, 143, 1247–1255. [Google Scholar] [CrossRef] [Green Version]
- Shamsaddini, A.; Gillevet, P.M.; Acharya, C.; Fagan, A.; Gavis, E.; Sikaroodi, M.; McGeorge, S.; Khoruts, A.; Albhaisi, S.; Fuchs, M.; et al. Impact of Antibiotic Resistance Genes in Gut Microbiome of Patients With Cirrhosis. Gastroenterology 2021, 161, 508–521.e7. [Google Scholar] [CrossRef]
- Kinny-Köster, B.; Bartels, M.; Becker, S.; Scholz, M.; Thiery, J.; Ceglarek, U.; Kaiser, T. Plasma Amino Acid Concentrations Predict Mortality in Patients with End-Stage Liver Disease. PLoS ONE 2016, 11, e0159205. [Google Scholar] [CrossRef]
- Dejong, C.H.C.; Van De Poll, M.C.G.; Soeters, P.B.; Jalan, R.; Olde Damink, S.W.M. Aromatic Amino Acid Metabolism during Liver Failure. J. Nutr. 2007, 137, 1579S–1585S. [Google Scholar] [CrossRef]
- Holecek, M. Ammonia and Amino Acid Profiles in Liver Cirrhosis: Effects of Variables Leading to Hepatic Encephalopathy. Nutrition 2015, 31, 14–20. [Google Scholar] [CrossRef]
- Solé, C.; Guilly, S.; Da Silva, K.; Llopis, M.; Le-Chatelier, E.; Huelin, P.; Carol, M.; Moreira, R.; Fabrellas, N.; De Prada, G.; et al. Alterations in Gut Microbiome in Cirrhosis as Assessed by Quantitative Metagenomics: Relationship With Acute-on-Chronic Liver Failure and Prognosis. Gastroenterology 2021, 160, 206–218.e13. [Google Scholar] [CrossRef]
- Chen, Y.; Guo, J.; Qian, G.; Fang, D.; Shi, D.; Guo, L.; Li, L. Gut Dysbiosis in Acute-on-Chronic Liver Failure and Its Predictive Value for Mortality. J. Gastroenterol. Hepatol. 2015, 30, 1429–1437. [Google Scholar] [CrossRef] [PubMed]
- Kang, D.J.; Kakiyama, G.; Betrapally, N.S.; Herzog, J.; Nittono, H.; Hylemon, P.B.; Zhou, H.; Carroll, I.; Yang, J.; Gillevet, P.M.; et al. Rifaximin Exerts Beneficial Effects Independent of Its Ability to Alter Microbiota Composition. Clin. Transl. Gastroenterol. 2016, 7, E187. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.Y.; Bajaj, J.S.; Wang, J.; Shang, J.; Zhou, X.M.; Guo, X.L.; Zhu, X.; Meng, L.N.; Jiang, H.X.; Mi, Y.Q.; et al. Lactulose Improves Cognition, Quality of Life and Gut Microbiota in Minimal Hepatic Encephalopathy: A Multi-Center, Randomized Controlled Trial. J. Dig. Dis. 2019, 20, 547–556. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Healthy Controls (n = 18) | Cirrhosis (n = 18) | p Value |
|---|---|---|---|
| Mean age (years) | 48.72 ± 8.63 | 49.89 ± 11.49 | 0.733 a |
| BMI (kg/m2) | 25.85 ± 2.86 | 24.66 ± 3.82 | 0.355 b |
| Hemoglobin (g/dL) | 15.1 ± 0.9 | 8.66 ± 2.4 | 0.000 a |
| Platelets (109/µL) | 229.59 ± 63.5 | 140.32 ± 90.18 | <0.002 a |
| White blood cells (109/µL) | 5.68 ± 1.42 | 17.13 ± 14.23 | 0.000 b |
| Neutrophils (109/µL) | 3.29 ± 1.05 | 14.01 ± 12.92 | 0.000 b |
| Lymphocytes (109/µL) | 1.8 ± 0.43 | 1.53 ± 0.94 | <0.001 b |
| Total bilirubin (mg/dL) | 0.67 ± 0.17 | 7.72 ± 8.99 | 0.000 b |
| Direct bilirubin (mg/dL) | 0.13 ± 0.05 | 2.26 ± 3.74 | <0.002 b |
| GGT (IU/L) | 25.67 ± 13.07 | 116.22 ± 92.83 | 0.000 b |
| Albumin (g/dL) | 4.47 ± 0.32 | 2.1 ± 0.58 | 0.000 b |
| ALT (U/L) | 26.94 ± 12.91 | 31 ± 17.07 | 0.284 a |
| AST (IU/L) | 22.89 ± 9.23 | 79.06 ± 46.98 | 0.000 b |
| ALP (IU/L) | 69.67 ± 22.21 | 142.39 ± 59.78 | 0.000 b |
| Total protein (g/dL) | 7.16 ± 0.47 | 2.94 ± 1.35 | 0.000 b |
| Creatinine (mg/dL) | 0.82 ± 0.12 | 1.42 ± 0.86 | <0.008 b |
| Prothrombin time (s) | 11.49 ± 0.77 | 23.26 ± 8.55 | 0.000 b |
| INR | 1.09 ± 0.08 | 2.13 ± 0.79 | 0.000 b |
| Sodium (mmol/L) | N/A | 131.94 ± 6.46 | N/A |
| Child–Pugh score | N/A | 11.39 ± 1.33 | N/A |
| MELD-Na score | N/A | 28.11 ± 6.69 | N/A |
| Prior or actual HE | N/A | 16 (88.9) | N/A |
| West Haven grade (1/2/3/4) | N/A | 1/8/7/0 | N/A |
| Ascites | N/A | 14 (77.8) | N/A |
| Upper gastrointestinal bleeding (UGIB) | N/A | 6 (33.3) | N/A |
| Acute kidney injury | N/A | 5 (27.8) | N/A |
| Lactulose | N/A | 18 (100) | N/A |
| Mean arterial pressure (mmHg) | N/A | 78.06 ± 12.05 | N/A |
| Duration of antibiotic treatment (days) | N/A | 3.94 ± 4.15 | N/A |
| Antibiotic type c (n) | 0 | 18 | N/A |
| Only ceftriaxone | - | 4 | |
| Ceftriaxone + rifaximin | - | 5 | |
| Ceftriaxone + other | - | 3 | |
| Others | - | 6 | |
| Use of proton pump inhibitors | 0 (0) | 8 (44.4) | N/A |
| Infection at admission | N/A | 10 (55.6) | N/A |
| Neutrophil-to-lymphocyte ratio | 1.87 ± 0.58 | 10.01 ± 10.46 | 0.000 b |
| AST/ALT ratio | 0.93 ± 0.26 | 5.89 ± 1.57 | 0.000 b |
| APRI index | 0.22 ± 0.13 | 1.49 ± 1.16 | 0.000 b |
| FIB-4 index | 1.1 ± 0.66 | 6.55 ± 4.04 | 0.000 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baltazar-Díaz, T.A.; González-Hernández, L.A.; Aldana-Ledesma, J.M.; Peña-Rodríguez, M.; Vega-Magaña, A.N.; Zepeda-Morales, A.S.M.; López-Roa, R.I.; del Toro-Arreola, S.; Martínez-López, E.; Salazar-Montes, A.M.; et al. Escherichia/Shigella, SCFAs, and Metabolic Pathways—The Triad That Orchestrates Intestinal Dysbiosis in Patients with Decompensated Alcoholic Cirrhosis from Western Mexico. Microorganisms 2022, 10, 1231. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10061231
Baltazar-Díaz TA, González-Hernández LA, Aldana-Ledesma JM, Peña-Rodríguez M, Vega-Magaña AN, Zepeda-Morales ASM, López-Roa RI, del Toro-Arreola S, Martínez-López E, Salazar-Montes AM, et al. Escherichia/Shigella, SCFAs, and Metabolic Pathways—The Triad That Orchestrates Intestinal Dysbiosis in Patients with Decompensated Alcoholic Cirrhosis from Western Mexico. Microorganisms. 2022; 10(6):1231. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10061231
Chicago/Turabian StyleBaltazar-Díaz, Tonatiuh Abimael, Luz Alicia González-Hernández, Juan Manuel Aldana-Ledesma, Marcela Peña-Rodríguez, Alejandra Natali Vega-Magaña, Adelaida Sara Minia Zepeda-Morales, Rocío Ivette López-Roa, Susana del Toro-Arreola, Erika Martínez-López, Adriana María Salazar-Montes, and et al. 2022. "Escherichia/Shigella, SCFAs, and Metabolic Pathways—The Triad That Orchestrates Intestinal Dysbiosis in Patients with Decompensated Alcoholic Cirrhosis from Western Mexico" Microorganisms 10, no. 6: 1231. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10061231