
Bacterial Metabolic Potential in Response to Climate Warming Alters the Decomposition Process of Aquatic Plant Litter—In Shallow Lake Mesocosms

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
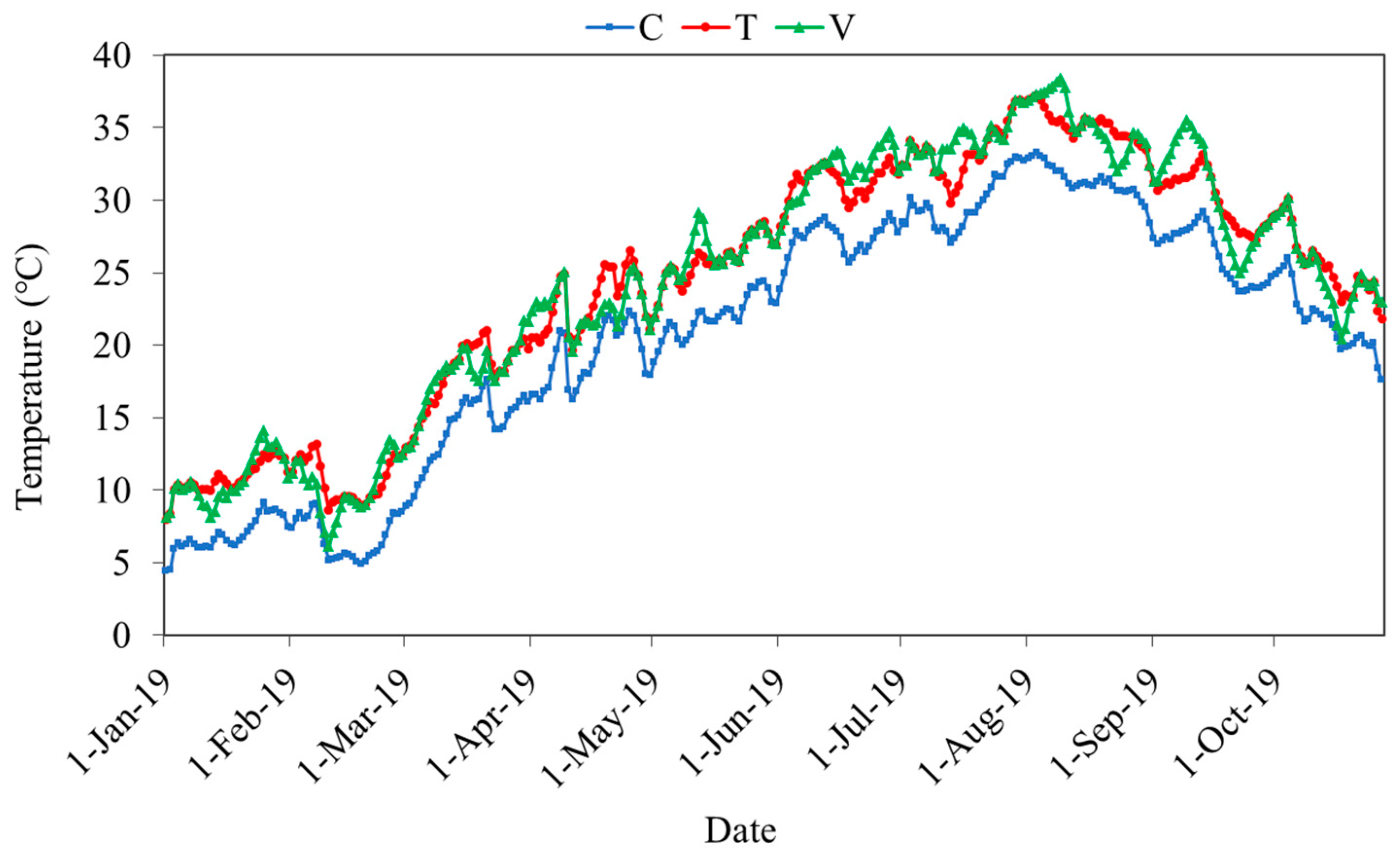
2.1. Mesocosm Experiment Set Up and Management
2.2. Sampling and Chemical Analyses
2.3. Decomposition Experiment Set Up and Sampling
2.4. DNA Extraction, PCR Amplification
2.5. FAPROTAX Analysis and Functional Prediction
2.6. Statistical Analysis
3. Results
3.1. Conditions in the Experimental Mesocosms
3.2. Bacterial Functional Abundance Analysis
3.3. Composition of Bacterial Functional Groups
3.4. Bacterial Metabolic Potential and Environmental Parameters Analysis
4. Discussion
4.1. Bacterial Metabolic Potentials in Leaf and Stem Litter Response to Climate Warming during Decomposition
4.2. Relationship between Bacterial Metabolism Potential and Environmental Factors under Climate Warming
4.3. Differences in the Responses of Bacterial Metabolic Potential of Leaf and Stem Litter to Warming and Environmental Factors
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McKenzie, D.J.; Geffroy, B.; Farrell, A.P. Effects of global warming on fishes and fisheries. J. Fish. Biol. 2021, 98, 1489–1492. [Google Scholar] [CrossRef] [PubMed]
- IPCC. Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Pachauri, R.K., Meyer, L.A., Eds.; IPCC: Geneva, Switzerland, 2014; p. 151. [Google Scholar]
- Janssen, A.B.G.; Hilt, S.; Kosten, S.; Klein, J.J.M.D.; Waal, D.B.V.D. Shifting states, shifting services: Linking regime shifts to changes in ecosystem services of shallow lakes. Freshw. Biol. 2021, 66, 1–12. [Google Scholar] [CrossRef]
- Zhang, Y.; Jeppesen, E.; Liu, X.; Qin, B.; Shi, K.; Zhou, Y.; Thomaz, S.M.; Deng, J. Global loss of aquatic vegetation in lakes. Earth-Sci. Rev. 2017, 173, 259–265. [Google Scholar] [CrossRef]
- Wu, J.; Cheng, S.; Liang, W.; He, F.; Wu, Z. Effects of sediment anoxia and light on turion germination and early growth of Potamogeton crispus. Hydrobiologia 2009, 628, 111–119. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Wang, T.; Molinos, J.G.; Li, C.; Hu, B.; Pan, M.; Zhang, M. Effects of warming, climate extremes and phosphorus enrichment on the growth, sexual reproduction and propagule carbon and nitrogen stoichiometry of Potamogeton crispus L. Environ. Int. 2020, 137, 105502. [Google Scholar] [CrossRef]
- Migliorini, G.H.; Romero, G.Q. Warming and leaf litter functional diversity, not litter quality, drive decomposition in a freshwater ecosystem. Sci. Rep. 2020, 10, 20333. [Google Scholar] [CrossRef]
- Glassman, S.I.; Weihe, C.; Li, J.; Albright, M.B.N.; Looby, C.I.; Martiny, A.C.; Treseder, K.K.; Allison, S.D.; Martiny, J.B.H. Decomposition responses to climate depend on microbial community composition. Proc. Natl. Aacad. Sci. USA 2018, 115, 11994. [Google Scholar] [CrossRef] [Green Version]
- Seer, F.K.; Putze, G.; Pennings, S.C.; Zimmer, M. Drivers of litter mass loss and faunal composition of detritus patches change over time. Ecol. Evol. 2021, 11, 9642–9651. [Google Scholar] [CrossRef]
- Dong, X.; Gao, P.; Zhou, R.; Li, C.; Dun, X.; Niu, X. Changing characteristics and influencing factors of the soil microbial community during litter decomposition in a mixed Quercus acutissima Carruth and Robinia pseudoacacia L forest in Northern China. Catena 2021, 196, 104811. [Google Scholar] [CrossRef]
- Kirschbaum, M.U.F. The temperature dependence of organic matter decomposition: Seasonal temperature variations turn a sharp short-term temperature response into a more moderate annually averaged response. Glob. Chang. Biol. 2010, 16, 2117–2129. [Google Scholar] [CrossRef]
- Jabiol, J.; Gossiaux, A.; Lecerf, A.; Rota, T.; Guérold, F.; Danger, M.; Poupin, P.; Gilbert, F.; Chauvet, E. Variable temperature effects between heterotrophic stream processes and organisms. Freshw. Biol. 2020, 65, 1543–1554. [Google Scholar] [CrossRef]
- Dang, C.K.; Schindler, M.; Chauvet, E.; Gessner, M.O. Temperature oscillation coupled with fungal community shifts can modulate warming effects on litter decomposition. Ecology 2009, 90, 122–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez, A.; Larrañaga, A.; Pérez, J.; Descals, E.; Pozo, J. Temperature affects leaf litter decomposition in low-order forest streams: Field and microcosm approaches. FEMS Microbiol. Ecol. 2014, 87, 257–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, V.; Canhoto, C. Effect of experimental and seasonal warming on litter decomposition in a temperate stream. Aquat. Sci. 2014, 76, 155–163. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, V.; Canhoto, C. Future increase in temperature may stimulate litter decomposition in temperate mountain streams: Evidence from a stream manipulation experiment. Freshw. Biol. 2015, 60, 881–892. [Google Scholar] [CrossRef]
- Mao, R.; Zhang, X.; Song, C.; Wang, X.; Finnegan, P. Plant functional group controls litter decomposition rate and its temperature sensitivity: An incubation experiment on litters from a boreal peatland in northeast China. Sci. Total. Environ. 2018, 626, 678–683. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, V.; Chauvet, E. Synergistic effects of water temperature and dissolved nutrients on litter decomposition and associated fungi. Glob. Chang. Biol. 2011, 17, 551–564. [Google Scholar] [CrossRef] [Green Version]
- Erhagen, B. Temperature sensitivity of soil carbon decomposition. Acta Univ. Agric. Suec. 2013, 440, 165–173. [Google Scholar] [CrossRef]
- Font, R.A.; Khamis, K.; Milner, A.M.; Smith, G.; Ledger, M.E. Low flow and heatwaves alter ecosystem functioning in a stream mesocosm experiment. Sci. Total. Environ. 2021, 777, 146067. [Google Scholar] [CrossRef]
- Pan, M.; Wang, T.; Hu, B.W.; Shi, P.L.; Xu, J.; Zhang, M. Mesocosm Experiments Reveal Global Warming Accelerates Macrophytes Litter Decomposition and Alters Decomposition-Related Bacteria Community Structure. Water 2021, 13, 1940. [Google Scholar] [CrossRef]
- Fierer, N.; Craine, J.M.; McLauchlan, K.; Schimel, J.P. Litter quality and the temperature sensitivity of decomposition. Ecology 2005, 86, 320–326. [Google Scholar] [CrossRef] [Green Version]
- Wilmot, O.J.; Hood, J.M.; Huryn, A.D.; Benstead, J.P. Decomposing decomposition: Isolating direct effects of temperature from other drivers of detrital processing. Ecology 2021, 102, e03467. [Google Scholar] [CrossRef] [PubMed]
- Bruder, A.; Salis, R.K.; McHugh, N.J.; Matthaei, C.D. Multiple-stressor effects on leaf litter decomposition and fungal decomposers in agricultural streams contrast between litter species. Funct. Ecol. 2016, 30, 1257–1266. [Google Scholar] [CrossRef] [Green Version]
- González-Benítez, N.; García-Corral, L.; Morán, X.; Middelburg, J.J.; Pizay, M.; Gattuso, J.P. Drivers of Microbial Carbon Fluxes Variability in Two Oligotrophic Mediterranean Coastal Systems. Sci. Rep. 2019, 9, 17669. [Google Scholar] [CrossRef]
- Goffredi, S.K.; Kantor, A.H.; Woodside, W.T. Aquatic Microbial Habitats Within a Neotropical Rainforest: Bromeliads and pH-Associated Trends in Bacterial Diversity and Composition. Microb. Ecol. 2011, 61, 529–542. [Google Scholar] [CrossRef]
- Berggren, M.; Laudon, H.; Jonsson, A.; Jansson, M. Nutrient Constraints on Metabolism Affect the Temperature Regulation of Aquatic Bacterial Growth Efficiency. Microb. Ecol. 2010, 60, 894–902. [Google Scholar] [CrossRef]
- López-Urrutia, A.; Morán, X. Resource limitation of bacterial production distorts the temperature dependence of oceanic carbon cycling. Ecology 2007, 88, 817–822. [Google Scholar] [CrossRef]
- Wu, S.; He, S.; Huang, J.; Gu, J.; Zhou, W.; Gao, L. Decomposition of Emergent Aquatic Plant (Cattail) Litter Under Different Conditions and the Influence on Water Quality. Water Air Soil Pollut. 2017, 228, 70. [Google Scholar] [CrossRef]
- Cornelissen, J.H.C.; Van Bodegom, P.M.; Aerts, R.; Callaghan, T.V.; Van Logtestijn, R.S.P.; Alatalo, J.; Chapin, F.S.; Gerdol, R.; Gudmundsson, J.; Gwynn-Jones, D. Global negative vegetation feedback to climate warming responses of leaf litter decomposition rates in cold biomes. Ecol. Lett. 2007, 10, 619–627. [Google Scholar] [CrossRef]
- Griffiths, N.A.; Tiegs, S.D. Organic-matter decomposition along a temperature gradient in a forested headwater stream. Freshw. Sci. 2016, 35, 518–533. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Wang, T.; Zhang, M.; Xu, J. Maternal Environment Effect of Warming and Eutrophication on the Emergence of Curled Pondweed, Potamogeton crispus L. Water 2018, 10, 1285. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Xu, J.; Molinos, J.G.; Li, C.; Hu, B.; Pan, M.; Zhang, M. A dynamic temperature difference control recording system in shallow lake mesocosm. MethodsX 2020, 7, 100930. [Google Scholar] [CrossRef] [PubMed]
- IPCC. Climate Change 2013: The Physical Science Basis; Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Tignor, M., Allen, S., Boschung, J., Nauels, A., Xia, Y., Bex, V., Pm, M., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013. [Google Scholar]
- Guo, X.; Huang, J.; Luo, Y.; Zhao, Z.; Xu, Y. Projection of heat waves over China for eight different global warming targets using 12 CMIP5 models. Theor. Appl. Climatol. 2017, 128, 507–522. [Google Scholar] [CrossRef]
- Jin, X.C.; Tu, Q.Y. Investigation Specifications for Lake Eutrophication, 2nd ed.; China Environmental Science Press: Beijing, China, 1990. [Google Scholar]
- Jespersen, A.; Christoffersen, K.J. Measurements of chlorophyll-a from phytoplankton using ethanol as extraction solvent. Arch. Hydrobiol. 1987, 109, 445–454. [Google Scholar]
- Murphy, J.; Riley, J.P. A modified single solution method for the determination of phosphate in natural waters. Anal. Chim. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Velthuis, M.; Kosten, S.; Aben, R.; Kazanjian, G.; Hilt, S. Warming enhances sedimentation and decomposition of organic carbon in shallow macrophyte-dominated systems with zero net effect on carbon burial. Glob. Chang. Biol. 2018, 24, 5231–5242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graças, M.; Bärlocher, F.; Gessner, M.O. Methods to Study Litter Decomposition: A Practical Guide; Springer: Dordrecht, The Netherlands, 2006; Volume 51, pp. 363–364. [Google Scholar] [CrossRef]
- Chen, M.; Zhu, X.; Zhao, C.; Yu, P.; Jia, H. Rapid microbial community evolution in initial Carex litter decomposition stages in Bayinbuluk alpine wetland during the freeze–thaw period. Ecol. Indic. 2021, 121, 107180. [Google Scholar] [CrossRef]
- Mori, H.; Maruyama, F.; Kato, H.; Toyoda, A.; Dozono, A.; Ohtsubo, Y.; Nagata, Y.; Fujiyama, A.; Tsuda, M.; Kurokawa, K. Design and Experimental Application of a Novel Non-Degenerate Universal Primer Set that Amplifies Prokaryotic 16S rRNA Genes with a Low Possibility to Amplify Eukaryotic rRNA Genes. DNA Res. 2014, 21, 217–227. [Google Scholar] [CrossRef]
- Shi, P.; Wang, H.; Feng, M.; Cheng, H.; Yang, Q.; Yan, Y.; Xu, J.; Zhang, M. The coupling response between different bacterial metabolic functions in water and sediment improve the ability to mitigate climate change. Water 2022, 14, 1203. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Jia, G. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Magoč, T.; Salzberg, S.L. FLASH: Fast Length Adjustment of Short Reads to Improve Genome Assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schloss, P.D.; Handelsman, J. Introducing DOTUR, a computer program for defining operational taxonomic units and estimating species richness. Appl. Environ. Microbiol. 2005, 71, 1501–1506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Louca, S.; Parfrey, L.W.; Doebeli, M. Decoupling function and taxonomy in the global ocean microbiome. Science 2016, 353, 1272–1277. [Google Scholar] [CrossRef]
- Kumar, A.; Ng, D.H.P.; Wu, Y.; Cao, B. Microbial community composition and putative biogeochemical functions in the sediment and water of tropical granite quarry lakes. Microb. Ecol. 2019, 77, 1–11. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Lenth, R.; Singmann, H.; Love, J.; Buerkner, P.; Herve, M. Package “Emmeans”; R Package Version 4.0-3; 2018. Available online: http://cran.r-project.org/package=emmeans (accessed on 28 June 2022).
- Ren, L.; Liu, Y.; Lauridsen, T.L.; Sndergaard, M.; Han, B.; Wang, J.; Jeppesen, E.; Zhou, J.; Wu, Q.L. Warming exacerbates the impact of nutrient enrichment on microbial functional potentials important to the nutrient cycling in shallow lake mesocosms. Limnol. Oceanogr. 2021, 66, 2481–2495. [Google Scholar] [CrossRef]
- Picazo, A.; Villaescusa, J.A.; Rochera, C.; Miralles-Lorenzo, J.; Quesada, A.; Camacho, A. Functional Metabolic Diversity of Bacterioplankton in Maritime Antarctic Lakes. Microorganisms 2021, 9, 2077. [Google Scholar] [CrossRef]
- Amblard, C.; Rachiq, S.; Bourdier, G. Photolithotrophy, photoheterotrophy and chemoheterotrophy during spring phytoplankton development (Lake Pavin). Microb. Ecol. 1992, 24, 109–123. [Google Scholar] [CrossRef]
- Galachyants, A.D.; Krasnopeev, A.Y.; Podlesnaya, G.V.; Potapov, S.A.; Sukhanova, E.V.; Tikhonova, I.V.; Zimens, E.A.; Kabilov, M.R.; Zhuchenko, N.A.; Gorshkova, A.S.; et al. Diversity of Aerobic Anoxygenic Phototrophs and Rhodopsin-Containing Bacteria in the Surface Microlayer, Water Column and Epilithic Biofilms of Lake Baikal. Microorganisms 2021, 9, 842. [Google Scholar] [CrossRef]
- Sharma, A.K.; Sommerfeld, K.; Bullerjahn, G.S.; Matteson, A.R.; Wilhelm, S.W.; Jezbera, J.; Brandt, U.; Doolittle, W.F.; Hahn, M.W. Actinorhodopsin genes discovered in diverse freshwater habitats and among cultivated freshwater Actinobacteria. ISME J. 2009, 3, 726–737. [Google Scholar] [CrossRef] [PubMed]
- Wagner-Döbler, I.; Biebl, H. Environmental biology of the marine Roseobacter lineage. Annu. Rev. Microbiol. 2006, 60, 255–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le, Y.L.; Wang, H.L. High-level soluble expression of a thermostable xylanase from thermophilic fungus Thermomyces lanuginosus in Escherichia coli via fusion with OsmY protein. Protein Expr. 2014, 99, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Lidstrom, M.E. Aerobic Methylotrophic Prokaryotes; Springer: New York, NY, USA, 2006. [Google Scholar]
- Battistuzzi, F.U.; Feijao, A.; Hedges, S.B. A genomic timescale of prokaryote evolution: Insights into the origin of methanogenesis, phototrophy, and the colonization of land. BMC Evol. Biol. 2004, 4, 44. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, L.; Bertilsson, S.; Peura, S. Non-cyanobacterial diazotrophs dominate nitrogen-fixing communities in permafrost thaw ponds. Limnol. Oceanogr. 2019, 65, S180–S193. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Yang, S.; Van, N.; Zhou, J.; Wei, F.; Qi, Q.; Liu, Y.; Wullschleger, S.D.; Liang, L.; Graham, D.E. Microbial Community and Functional Gene Changes in Arctic Tundra Soils in a Microcosm Warming Experiment. Front. Microbiol. 2017, 8, 1741. [Google Scholar] [CrossRef] [Green Version]
- Vekeman, B.; Kerckhof, F.-M.; Cremers, G.; De Vos, P.; Vandamme, P.; Boon, N.; Op den Camp, H.H.; Heylen, K. New Methyloceanibacter diversity from North Sea sediments includes methanotroph containing solely the soluble methane monooxygenase. Environ. Microbiol. 2016, 8, 4523–4536. [Google Scholar] [CrossRef] [Green Version]
- Chistoserdova, L. Modularity of methylotrophy, revisited. Environ. Microbiol. 2011, 13, 2603–2622. [Google Scholar] [CrossRef]
- Ho, A.; Luke, C.; Frenzel, P. Recovery of methanotrophs from disturbances: Population dynamics, evenness and functioning. ISME J. 2011, 5, 750–758. [Google Scholar] [CrossRef] [Green Version]
- Hu, B.L.; Shen, L.D.; Lian, X.; Zhu, Q.; Liu, S.; Huang, Q.; He, Z.F.; Geng, S.; Cheng, D.Q.; Lou, L.P.; et al. Evidence for nitrite-dependent anaerobic methane oxidation as a previously overlooked microbial methane sink in wetlands. Proc. Natl. Aacad. Sci. USA 2014, 111, 4495–4500. [Google Scholar] [CrossRef] [Green Version]
- Vojtěch, T.; Jana, V.; Petr, B. Bacterial succession on decomposing leaf litter exhibits a specific occurrence pattern of cellulolytic taxa and potential decomposers of fungal mycelia. FEMS Microbiol. Ecol. 2016, 92, 177. [Google Scholar] [CrossRef]
- Treseder, K.; Kivlin, S.; Hawkes, C. Evolutionary trade-offs among decomposers determine responses to nitrogen enrichment. Ecol. Lett. 2011, 14, 933–938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, P.; Markham, J. Soil temperature limits nitrogen fixation, photosynthesis, and growth in a boreal actinorhizal shrub. Plant. Soil. 2021, 468, 411–421. [Google Scholar] [CrossRef]
- Houlton, B.Z.; Wang, Y.P.; Vitousek, P.M.; Field, C.B. A unifying framework for dinitrogen fixation in the terrestrial biosphere. Nature 2008, 454, 327–330. [Google Scholar] [CrossRef] [PubMed]
- Gillooly, J.; Brown, J.; West, G.; Savage, V.; Charnov, E. Effects of Size and Temperature on Metabolic Rate. Science 2001, 293, 2248–2251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stief, P.; Lundgaard, A.S.B.; Treusch, A.H.; Thamdrup, B.; Grossart, H.-P.; Glud, R.N. Freshwater copepod carcasses as pelagic microsites of dissimilatory nitrate reduction to ammonium. FEMS Microbiol. Ecol. 2018, 94, 144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, K.; Yuan, M.M.; Zhou, J.S.; Qin, Y.; Deng, Y.; Cheng, L.; Wu, L.; He, Z.; Van Nostrand, J.D.; Bracho, R.; et al. Tundra soil carbon is vulnerable to rapid microbial decomposition under climate warming. Nat. Clim. Chang. 2016, 6, 595–600. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.Z.; Kai, X.; Xie, J.P.; Ye, D.; Luo, Y.Q. Microbial mediation of carbon-cycle feedbacks to climate warming. Nat. Clim. Chang. 2011, 2, 106–110. [Google Scholar] [CrossRef]
- Tscherko, D.; Kandeler, E.; Jones, T.H. Effect of temperature on below-ground N-dynamics in a weedy model ecosystem at ambient and elevated atmospheric CO2 levels. Soil Biol. Biochem. 2001, 33, 491–501. [Google Scholar] [CrossRef]
- Sara, H.; Allana, W.; John, S.; Stephanie, H.; Karin, E.; David, B.; Laurent, P.; Minna-Maarit, K. Soil Functional Operating Range Linked to Microbial Biodiversity and Community Composition Using Denitrifiers as Model Guild. PLoS ONE 2012, 7, e51962. [Google Scholar] [CrossRef] [Green Version]
- King, D.; Nedwell, D.B. Changes in the Nitrate-reducing Community of an Anaerobic Saltmarsh Sediment in Response to Seasonal Selection by Temperature. Microbiology 1984, 130, 2935–2941. [Google Scholar] [CrossRef] [Green Version]
- Tan, E.; Zou, W.; Zheng, Z.; Yan, X.; Kao, S.J. Warming stimulates sediment denitrification at the expense of anaerobic ammonium oxidation. Nat. Clim. Chang. 2020, 10, 349–355. [Google Scholar] [CrossRef]
- Canion, A.; Overholt, W.A.; Kostka, J.E.; Huettel, M.; Lavik, G.; Kuypers, M. Temperature response of denitrification and anaerobic ammonium oxidation rates and microbial community structure in Arctic fjord sediments. Environ. Microbiol. 2014, 16, 3331–3344. [Google Scholar] [CrossRef] [PubMed]
- Bahram, M.; Hildebrand, F.; Forslund, S.K.; Anderson, J.L.; Soudzilovskaia, N.A.; Bodegom, P.M.; Bengtsson-Palme, J.; Anslan, S.; Coelho, L.P.; Harend, H. Structure and function of the global topsoil microbiome. Nature 2018, 560, 233–237. [Google Scholar] [CrossRef] [PubMed]
- Beales, N. Adaptation of Microorganisms to Cold Temperatures, Weak Acid Preservatives, Low pH, and Osmotic Stress: A Review. Compr. Rev. Food. Sci. Food Saf. 2010, 3, 1–20. [Google Scholar] [CrossRef]
- Curtin, D.; Peterson, M.E.; Anderson, C.R. pH-dependence of organic matter solubility: Base type effects on dissolved organic C, N, P, and S in soils with contrasting mineralogy. Geoderma 2016, 271, 161–172. [Google Scholar] [CrossRef]
- Rousk, J.; Bååth, E.; Brookes, P.C.; Lauber, C.L.; Lozupone, C.; Caporaso, J.G.; Knight, R.; Fierer, N. Soil bacterial and fungal communities across a pH gradient in an arable soil. ISME J. 2010, 4, 1340–1351. [Google Scholar] [CrossRef]
- Černá, B.; Rejmánková, E.; Snyder, J.M.; Šantrůčková, H. Heterotrophic nitrogen fixation in oligotrophic tropical marshes: Changes after phosphorus addition. Hydrobiologia 2009, 627, 55–65. [Google Scholar] [CrossRef] [Green Version]
- Markham, J.H.; Zekveld, C. Nitrogen fixation makes biomass allocation to roots independent of soil nitrogen supply. Can. J. Bot. 2007, 85, 787–793. [Google Scholar] [CrossRef]
- Gentili, F.; Wall, L.G.; Huss-Danell, K. Effects of Phosphorus and Nitrogen on Nodulation are Seen Already at the Stage of Early Cortical Cell Divisions in Alnus incana. Ann. Bot. 2006, 98, 309–315. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, S.; Andersen, K.H.; Visser, A.W.; Inomura, K.; Riemann, L. Quantifying nitrogen fixation by heterotrophic bacteria in sinking marine particles. Nat. Commun. 2021, 12, 4085. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, I.; Nadler, V.; Hochman, A. Mechanism of nitrogenase switch-off by oxygen. J. Bacteriol. 1987, 169, 874–879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, T.C.; Aguilar, J.V.; Souza, L.A.; Justino, G.C.; Aguiar, L.F.; Camargos, L.S. pH effects on nodulation and biological nitrogen fixation in Calopogonium mucunoides. Braz. J. Bot. 2016, 39, 1015–1020. [Google Scholar] [CrossRef] [Green Version]
- Hou, L.; Wang, R.; Yin, G.; Liu, M.; Zheng, Y. Nitrogen Fixation in the Intertidal Sediments of the Yangtze Estuary: Occurrence and Environmental Implications. J. Geophys. Res-Biogeo. 2018, 123, 936–944. [Google Scholar] [CrossRef]
- Burris, R.H. Comparative study of the response ofAzotobacter vinelandii andAcetobacter diazotrophicus to changes in pH. Protoplasma 1994, 183, 62–66. [Google Scholar] [CrossRef]
- Taylor, B.R.; Parkinson, D.; Parsons, W.F. Nitrogen and lignin content as predictors of litter decay rates: A microcosm test. Ecology 1989, 70, 97–104. [Google Scholar] [CrossRef]
- Prieto, I.; Almagro, M.; Bastida, F.; Querejeta, J.I. Altered leaf litter quality exacerbates the negative impact of climate change on decomposition. J. Ecol. 2018, 107, 2364–2382. [Google Scholar] [CrossRef]
- Yang, Z.; Song, W.; Zhao, Y.; Zhou, J.; Wang, Z.; Luo, Y.; Li, Y.; Lin, G. Differential responses of litter decomposition to regional excessive nitrogen input and global warming between two mangrove species. Estuar. Coast. Shelf Sci. 2018, 214, 141–148. [Google Scholar] [CrossRef]
- Zhang, P.; BaKker, E.S.; Zhang, M.; Xu, J.; Zhang, P.; Ba Kker, E.S.; Zhang, M.; Xu, J. Effects of warming on Potamogeton crispus growth and tissue stoichiometry in the growing season. Aquat. Bot. 2016, 128, 13–17. [Google Scholar] [CrossRef] [Green Version]
- Elser, J.J.; Fagan, W.F.; Kerkhoff, A.J.; Enquist, N. Biological stoichiometry of plant production: Metabolism, scaling and ecological response to global change. New. Phytol. 2010, 186, 593–608. [Google Scholar] [CrossRef] [Green Version]
- Wang, T. The Growth snd Reproduction of Potamogeton crispus L. Response to Global Different Warming Patterns and Eutrophication. Master’s Thesis, Huazhong Agricultural University, Wuhan, China, 2019. [Google Scholar]
- Pope, C.A.; Halvorson, H.M.; Findlay, R.H.; Francoeur, S.N.; Kuehn, K.A. Light and temperature mediate algal stimulation of heterotrophic activity on decomposing leaf litter. Freshw. Biol. 2020, 65, 1210–1222. [Google Scholar] [CrossRef]
- Florent, R.; Clarisse, M.; Christophe, P.; Florence, D.; Frédérique, B.; Joan, A. Stimulation or inhibition: Leaf microbial decomposition in streams subjected to complex chemical contamination. Sci. Total. Environ. 2018, 648, 1371–1383. [Google Scholar] [CrossRef]
- Lidman, J.; Jonsson, M.; Burrows, R.M.; Bundschuh, M.; Sponseller, R.A. Composition of riparian litter input regulates organic matter decomposition: Implications for headwater stream functioning in a managed forest landscape. Ecol. Evol. 2017, 7, 1068–1077. [Google Scholar] [CrossRef] [PubMed]
- Ardón, M.; Pringle, C.M.; Eggert, S.L. Does leaf chemistry differentially affect breakdown in tropical vs temperate streams? Importance of standardized analytical techniques to measure leaf chemistry. Freshw. Sci. 2009, 28, 440–453. [Google Scholar] [CrossRef]
- Zhang, M.; Cheng, X.; Geng, Q.; Shi, Z.; Luo, Y.; Xu, X. Leaf litter traits predominantly control litter decomposition in streams worldwide. Global. Ecol. Biogeogr. 2019, 28, 1469–1486. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pH | Cond (μs·cm−1) | DO (mg·L−1) | TN (mg·L−1) | TP (μg·L−1) | PO43−-P (μg·L−1) | NH4+-N (mg·L−1) | NO3−-N (mg·L−1) | Chl-a (μg·L−1) | ||
|---|---|---|---|---|---|---|---|---|---|---|
| Spring | C | 9.43 ± 0.18 a | 170.57 ± 7.81 a | 12.44 ± 0.44 a | 0.38 ± 0.04 a | 0.03 ± 0.01 a | 5.81 ± 1.33 a | 0.10 ± 0.01 a | 0.08 ± 0.01 a | 11.11 ± 1.47 a |
| T | 9.43 ± 0.18 a | 230.48 ± 7.53 b | 10.51 ± 0.40 a | 0.67 ± 0.06 b | 0.07 ± 0.02 a | 17.91 ± 7.91 a | 0.11 ± 0.02 a | 0.12 ± 0.02 a | 14.01 ± 2.33 a | |
| V | 9.49 ± 0.12 a | 212.31 ± 7.16 b | 10.39 ± 0.56 a | 0.54 ± 0.07 ab | 0.06 ± 0.01 a | 26.32 ± 7.77 a | 0.16 ± 0.02 a | 0.11 ± 0.02 a | 15.67 ± 2.85 a | |
| Summer | C | 7.59 ± 0.17 a | 180.05 ± 9.44 a | 3.74 ± 0.74 a | 0.65 ± 0.08 a | 0.08 ± 0.02 a | 58.38 ± 34.24 a | 0.09 ± 0.02 a | 0.11 ± 0.01 a | 4.77 ± 1.09 a |
| T | 8.31 ± 0.18 b | 247.06 ± 11.10 b | 5.92 ± 0.52 b | 1.03 ± 0.11 b | 0.13 ± 0.03 a | 59.45 ± 9.30 ab | 0.19 ± 0.04 a | 0.18 ± 0.03 b | 11.12 ± 3.67 a | |
| V | 8.03 ± 0.19 ab | 243.77 ± 6.39 b | 4.19 ± 0.50 ab | 0.81 ± 0.05 ab | 0.12 ± 0.02 a | 98.50 ± 24.36 b | 0.17 ± 0.02 a | 0.15 ± 0.01 ab | 3.30 ± 0.51 a | |
| Autumn | C | 7.85 ± 0.19 a | 265.91 ± 18.40 a | 4.27 ± 0.98 a | 0.58 ± 0.07 a | 0.04 ± 0.00 a | 20.21 ± 3.68 a | 0.21 ± 0.04 a | 0.18 ± 0.01 a | 1.93 ± 0.32 a |
| T | 8.38 ± 0.16 a | 315.61 ± 11.81 b | 6.85 ± 1.12 a | 1.25 ± 0.11 b | 0.06 ± 0.01 a | 58.05 ± 25.31 a | 0.42 ± 0.17 a | 0.23 ± 0.03 a | 2.57 ± 0.32 a | |
| V | 8.25 ± 0.15 a | 335.50 ± 9.83 b | 5.55 ± 0.91 a | 1.02 ± 0.09 b | 0.05 ± 0.01 a | 22.62 ± 4.17 a | 0.28 ± 0.04 a | 0.23 ± 0.01 a | 2.17 ± 0.41 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, P.; Wang, H.; Feng, M.; Cheng, H.; Yang, Q.; Yan, Y.; Xu, J.; Zhang, M. Bacterial Metabolic Potential in Response to Climate Warming Alters the Decomposition Process of Aquatic Plant Litter—In Shallow Lake Mesocosms. Microorganisms 2022, 10, 1327. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10071327
Shi P, Wang H, Feng M, Cheng H, Yang Q, Yan Y, Xu J, Zhang M. Bacterial Metabolic Potential in Response to Climate Warming Alters the Decomposition Process of Aquatic Plant Litter—In Shallow Lake Mesocosms. Microorganisms. 2022; 10(7):1327. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10071327
Chicago/Turabian StyleShi, Penglan, Huan Wang, Mingjun Feng, Haowu Cheng, Qian Yang, Yifeng Yan, Jun Xu, and Min Zhang. 2022. "Bacterial Metabolic Potential in Response to Climate Warming Alters the Decomposition Process of Aquatic Plant Litter—In Shallow Lake Mesocosms" Microorganisms 10, no. 7: 1327. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10071327