
Prevalence of pvmrp1 Polymorphisms and Its Contribution to Antimalarial Response

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites and Participants
2.2. Gene Sequence and Protein Structure Prediction
2.3. Single Nucleotide Polymorphisms (SNPs) Identification in pvmrp1 Gene
2.4. Determination of pvmrp1 Copy Number (CN)
2.5. Data Analysis
3. Results
3.1. Characterization of PvMRP1
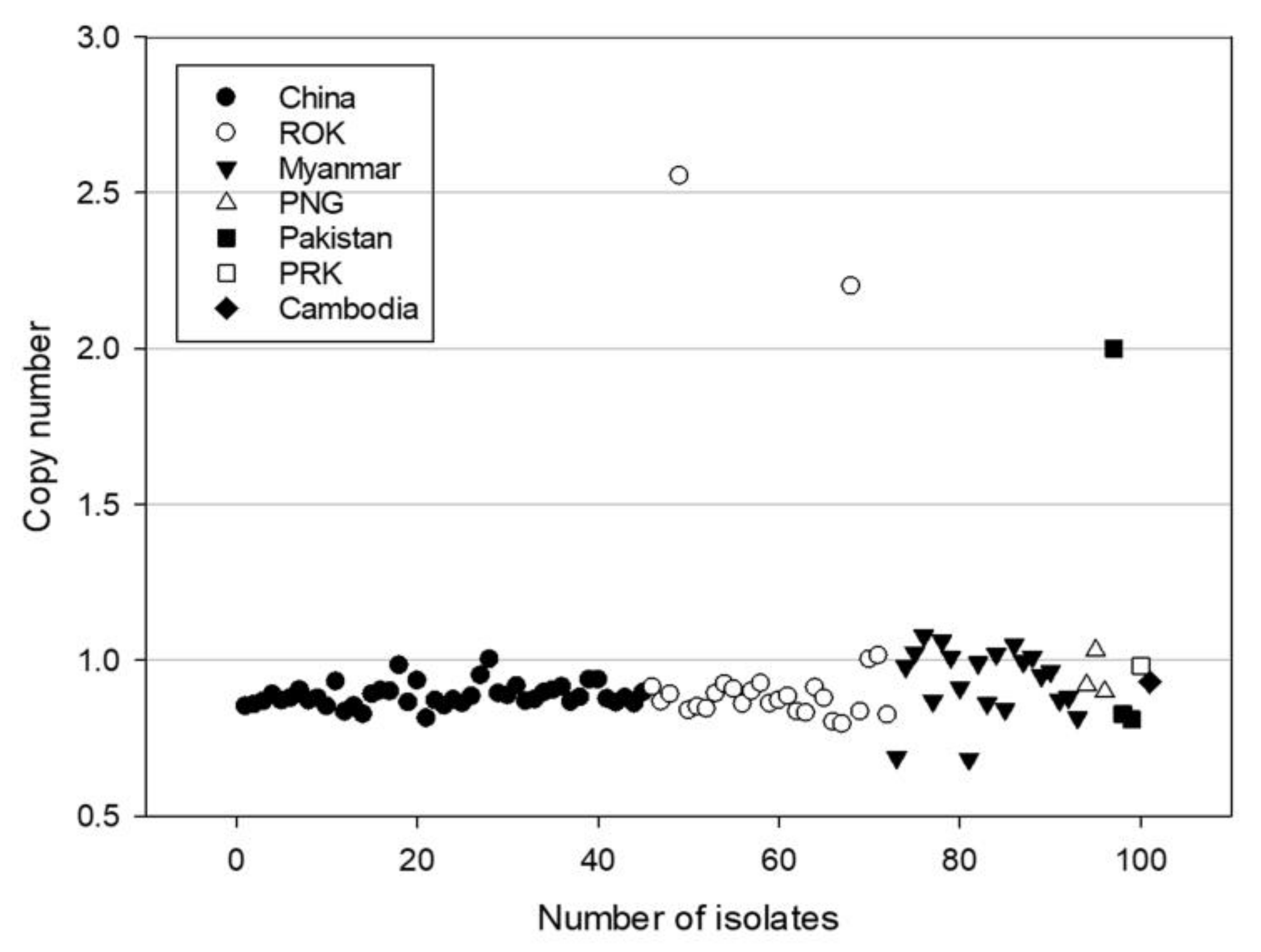
3.2. Identification of Gene Mutations and CN in pvmrp1 among Collected Blood Samples in Asia
3.3. Correlation between Polymorphisms and In Vitro Drug Susceptibilities
3.4. The Polymorphism of pvmrp1 from Different Regions
3.5. Natural Selection of Polymorphic Region of pvmrp1 from Different P. vivax Isolates
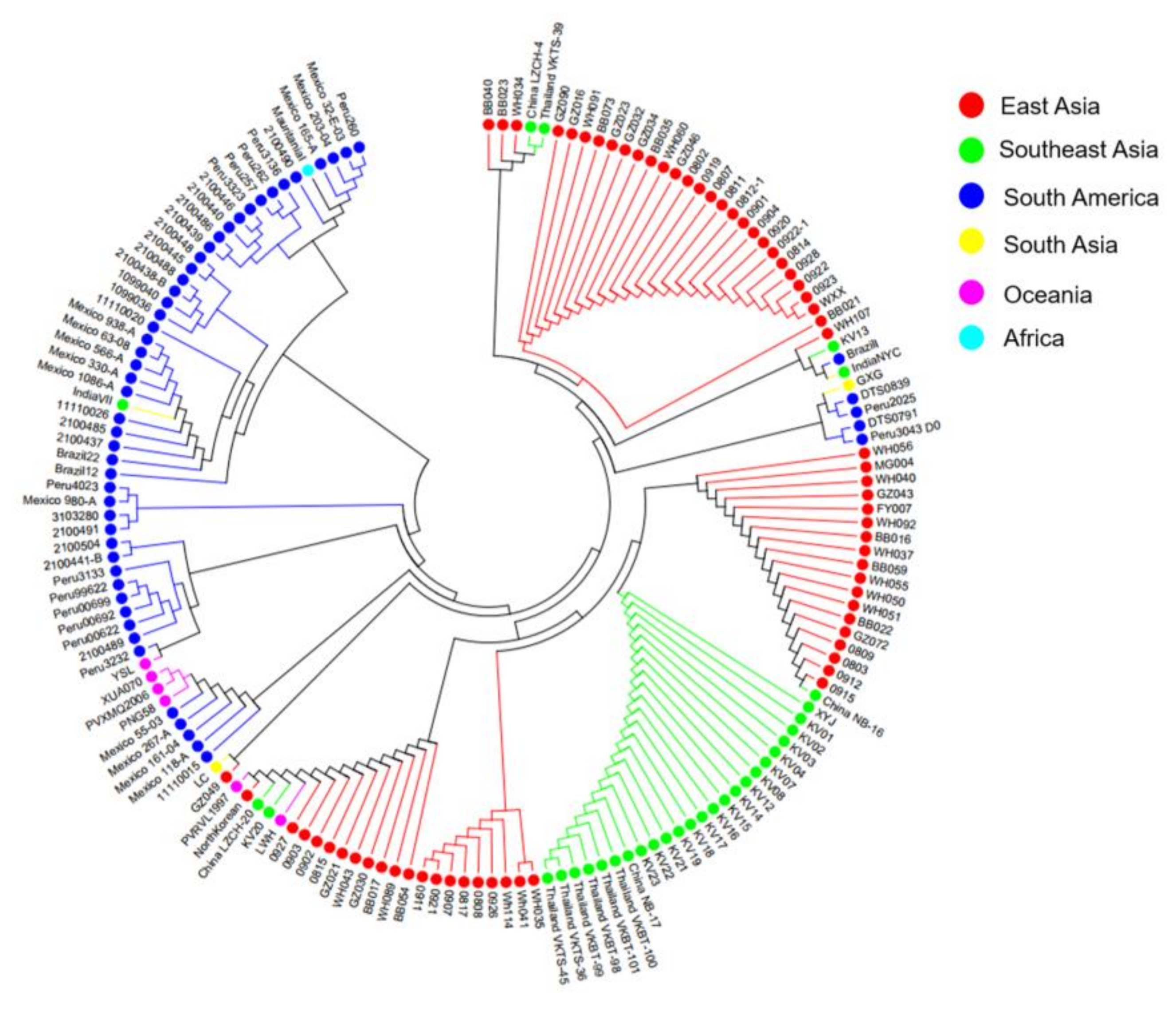
3.6. Genetic Differentiation, Haplotype Network and LD Analysis of Polymorphic Region of pvmrp1
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- WHO. World Malaria Report 2021. Available online: https://www.who.int/publications/i/item/9789240040496 (accessed on 6 December 2021).
- Battle, K.E.; Lucas, T.C.D.; Nguyen, M.; Howes, R.E.; Nandi, A.K.; Twohig, K.A.; Pfeffer, D.A.; Cameron, E.; Rao, P.C.; Casey, D.; et al. Mapping the global endemicity and clinical burden of Plasmodium vivax, 2000–2017: A spatial and temporal modelling study. Lancet 2019, 394, 332–343. [Google Scholar] [CrossRef] [Green Version]
- Rieckmann, K.H.; Davis, D.R.; Hutton, D.C. Plasmodium vivax resistance to chloroquine? Lancet 1989, 2, 1183–1184. [Google Scholar] [CrossRef]
- Cotter, C.; Sturrock, H.J.; Hsiang, M.S.; Liu, J.; Phillips, A.A.; Hwang, J.; Gueye, C.S.; Fullman, N.; Gosling, R.D.; Feachem, R.G. The changing epidemiology of malaria elimination: New strategies for new challenges. Lancet 2013, 382, 900–911. [Google Scholar] [CrossRef]
- Xu, S.; Zeng, W.; Ngassa Mbenda, H.G.; Liu, H.; Chen, X.; Xiang, Z.; Li, C.; Zhang, Y.; Baird, J.K.; Yang, Z.; et al. Efficacy of directly-observed chloroquine-primaquine treatment for uncomplicated acute Plasmodium vivax malaria in northeast Myanmar: A prospective open-label efficacy trial. Travel Med. Infect. Dis. 2020, 36, 101499. [Google Scholar] [CrossRef] [PubMed]
- Htun, M.W.; Mon, N.C.N.; Aye, K.M.; Hlaing, C.M.; Kyaw, M.P.; Handayuni, I.; Trimarsanto, H.; Bustos, D.; Ringwald, P.; Price, R.N.; et al. Chloroquine efficacy for Plasmodium vivax in Myanmar in populations with high genetic diversity and moderate parasite gene flow. Malar. J. 2017, 16, 281. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Wang, Y.; Parker, D.M.; Gupta, B.; Yang, Z.; Liu, H.; Fan, Q.; Cao, Y.; Xiao, Y.; Lee, M.C.; et al. Therapeutic responses of Plasmodium vivax malaria to chloroquine and primaquine treatment in northeastern Myanmar. Antimicrob. Agents Chemother. 2015, 59, 1230–1235. [Google Scholar] [CrossRef] [Green Version]
- Ngassa Mbenda, H.G.; Wang, M.; Guo, J.; Siddiqui, F.A.; Hu, Y.; Yang, Z.; Kittichai, V.; Sattabongkot, J.; Cao, Y.; Jiang, L.; et al. Evolution of the Plasmodium vivax multidrug resistance 1 gene in the Greater Mekong Subregion during malaria elimination. Parasites Vectors 2020, 13, 67. [Google Scholar] [CrossRef]
- Diez Benavente, E.; Ward, Z.; Chan, W.; Mohareb, F.R.; Sutherland, C.J.; Roper, C.; Campino, S.; Clark, T.G. Genomic variation in Plasmodium vivax malaria reveals regions under selective pressure. PLoS ONE 2017, 12, e0177134. [Google Scholar] [CrossRef] [Green Version]
- Benavente, E.D.; Manko, E.; Phelan, J.; Campos, M.; Nolder, D.; Fernandez, D.; Velez-Tobon, G.; Castaño, A.T.; Dombrowski, J.G.; Marinho, C.R.F.; et al. Distinctive genetic structure and selection patterns in Plasmodium vivax from South Asia and East Africa. Nat. Commun. 2021, 12, 3160. [Google Scholar] [CrossRef]
- Valderramos, S.G.; Fidock, D.A. Transporters involved in resistance to antimalarial drugs. Trends Pharmacol. Sci. 2006, 27, 594–601. [Google Scholar] [CrossRef] [Green Version]
- Srikant, S.; Gaudet, R.; Murray, A.W. Selecting for Altered Substrate Specificity Reveals the Evolutionary Flexibility of ATP-Binding Cassette Transporters. Curr. Biol. 2020, 30, 1689–1702.e1686. [Google Scholar] [CrossRef] [Green Version]
- Gil, J.P.; Fançony, C. Plasmodium falciparum Multidrug Resistance Proteins (pfMRPs). Front. Pharmacol. 2021, 12, 759422. [Google Scholar] [CrossRef]
- Dahlström, S.; Veiga, M.I.; Mårtensson, A.; Björkman, A.; Gil, J.P. Polymorphism in PfMRP1 (Plasmodium falciparum multidrug resistance protein 1) amino acid 1466 associated with resistance to sulfadoxine-pyrimethamine treatment. Antimicrob. Agents Chemother. 2009, 53, 2553–2556. [Google Scholar] [CrossRef] [Green Version]
- Gendrot, M.; Wague Gueye, M.; Tsombeng Foguim, F.; Madamet, M.; Wade, K.A.; Bou Kounta, M.; Fall, M.; Diawara, S.; Benoit, N.; Lo, G.; et al. Modulation of in vitro antimalarial responses by polymorphisms in Plasmodium falciparum ABC transporters (pfmdr1 and pfmdr5). Acta Trop. 2019, 196, 126–134. [Google Scholar] [CrossRef]
- Kavishe, R.A.; Paulo, P.; Kaaya, R.D.; Kalinga, A.; van Zwetselaar, M.; Chilongola, J.; Roper, C.; Alifrangis, M. Surveillance of artemether-lumefantrine associated Plasmodium falciparum multidrug resistance protein-1 gene polymorphisms in Tanzania. Malar. J. 2014, 13, 264. [Google Scholar] [CrossRef] [Green Version]
- Rijpma, S.R.; van der Velden, M.; Bilos, A.; Jansen, R.S.; Mahakena, S.; Russel, F.G.; Sauerwein, R.W.; van de Wetering, K.; Koenderink, J.B. MRP1 mediates folate transport and antifolate sensitivity in Plasmodium falciparum. FEBS Lett. 2016, 590, 482–492. [Google Scholar] [CrossRef] [Green Version]
- Noulin, F.; Borlon, C.; Van Den Abbeele, J.; D’Alessandro, U.; Erhart, A. 1912-2012: A century of research on Plasmodium vivax in vitro culture. Trends Parasitol. 2013, 29, 286–294. [Google Scholar] [CrossRef]
- Suwanarusk, R.; Russell, B.; Chavchich, M.; Chalfein, F.; Kenangalem, E.; Kosaisavee, V.; Prasetyorini, B.; Piera, K.A.; Barends, M.; Brockman, A.; et al. Chloroquine resistant Plasmodium vivax: In vitro characterisation and association with molecular polymorphisms. PLoS ONE 2007, 2, e1089. [Google Scholar] [CrossRef]
- Lu, F.; Gao, Q.; Chotivanich, K.; Xia, H.; Cao, J.; Udomsangpetch, R.; Cui, L.; Sattabongkot, J. In vitro anti-malarial drug susceptibility of temperate Plasmodium vivax from central China. Am. J. Trop. Med. Hyg. 2011, 85, 197–201. [Google Scholar] [CrossRef] [Green Version]
- Lu, F.; Lim, C.S.; Nam, D.H.; Kim, K.; Lin, K.; Kim, T.S.; Lee, H.W.; Chen, J.H.; Wang, Y.; Sattabongkot, J.; et al. Genetic polymorphism in pvmdr1 and pvcrt-o genes in relation to in vitro drug susceptibility of Plasmodium vivax isolates from malaria-endemic countries. Acta Trop. 2011, 117, 69–75. [Google Scholar] [CrossRef]
- Yang, M.; Derbyshire, M.K.; Yamashita, R.A.; Marchler-Bauer, A. NCBI’s Conserved Domain Database and Tools for Protein Domain Analysis. Curr. Protoc. Bioinform. 2020, 69, e90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Letunic, I.; Khedkar, S.; Bork, P. SMART: Recent updates, new developments and status in 2020. Nucleic Acids Res. 2021, 49, D458–D460. [Google Scholar] [CrossRef] [PubMed]
- Krogh, A.; Larsson, B.; von Heijne, G.; Sonnhammer, E.L. Predicting transmembrane protein topology with a hidden Markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Tajima, F. Simple methods for testing the molecular evolutionary clock hypothesis. Genetics 1993, 135, 599–607. [Google Scholar] [CrossRef]
- Hall, B.G. Building phylogenetic trees from molecular data with MEGA. Mol. Biol. Evol. 2013, 30, 1229–1235. [Google Scholar] [CrossRef] [Green Version]
- Bandelt, H.J.; Forster, P.; Röhl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Lu, F.; Wang, B.; Cao, J.; Sattabongkot, J.; Zhou, H.; Zhu, G.; Kim, K.; Gao, Q.; Han, E.T. Prevalence of drug resistance-associated gene mutations in Plasmodium vivax in Central China. Korean J. Parasitol. 2012, 50, 379–384. [Google Scholar] [CrossRef]
- Carlton, J.M.; Fidock, D.A.; Djimdé, A.; Plowe, C.V.; Wellems, T.E. Conservation of a novel vacuolar transporter in Plasmodium species and its central role in chloroquine resistance of P. falciparum. Curr. Opin. Microbiol. 2001, 4, 415–420. [Google Scholar] [CrossRef]
- Sharrock, W.W.; Suwanarusk, R.; Lek-Uthai, U.; Edstein, M.D.; Kosaisavee, V.; Travers, T.; Jaidee, A.; Sriprawat, K.; Price, R.N.; Nosten, F.; et al. Plasmodium vivax trophozoites insensitive to chloroquine. Malar. J. 2008, 7, 94. [Google Scholar] [CrossRef] [Green Version]
- Zeng, W.; Zhao, H.; Zhao, W.; Yang, Q.; Li, X.; Li, X.; Duan, M.; Wang, X.; Li, C.; Xiang, Z.; et al. Molecular Surveillance and Ex Vivo Drug Susceptibilities of Plasmodium vivax Isolates From the China-Myanmar Border. Front. Cell. Infect. Microbiol. 2021, 11, 738075. [Google Scholar] [CrossRef]
- Anderson, T.J.; Patel, J.; Ferdig, M.T. Gene copy number and malaria biology. Trends Parasitol. 2009, 25, 336–343. [Google Scholar] [CrossRef] [Green Version]
- Hiroi, N.; Takahashi, T.; Hishimoto, A.; Izumi, T.; Boku, S.; Hiramoto, T. Copy number variation at 22q11.2: From rare variants to common mechanisms of developmental neuropsychiatric disorders. Mol. Psychiatry 2013, 18, 1153–1165. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Purpose | Primer Name | Role in PCR | Sequence (5′-3′) | Position | PCR Product (bp) | Annealing Temperature |
|---|---|---|---|---|---|---|
| Sequencing | ||||||
| Fragment 1 | Frag1-F1 | First, second and Seq PCR | AAA AAA TAA GCC AAA TTA ACC TTA CAC | −48–−74 | First round 2500 bp | 56 °C |
| Frag1-R1 | First PCR | GGC AAA ATG CAC TTA TTT TGT CT | 2430–2452 | |||
| Frag1-R2 | Second PCR | TTT ATG TTC TTC AAA ATG TAC TTC TCA | 1926–1952 | second round 2000 bp | 56 °C | |
| Frag1-SF1 | Seq PCR | GGA TTT TTT TTA ACA CTG TTA CAG CTT | 726–752 | |||
| Frag1-SR2 | Seq PCR | GAT TTA ACG AAT CAT TCT GTG TAT AGA AA | 652–680 | |||
| Frag1-SR3 | Seq PCR | CAT AGT TGA AGT GTT GTT ATT TTT TTT GT | 1302–1330 | |||
| Fragment 2 | Frag2-F1 | First PCR | TGA TGT AGA GAA AGT GTG TTT CCA G | 1752–1776 | First round 2401 bp | 61 °C |
| Frag2-R1 | First PCR | CTC CTC TAG CCT CTG CAC ACA | 4132–4152 | |||
| Frag2-F2 | Second and Seq PCR | TTA GAA AAT GCT TTT TTT GGC AC | 1882–1904 | second round 1971 bp | 56 °C | |
| Frag2-R2 | Second PCR | TAT ACC TAA ATG GTA CCA ATT CTT TTC | 3826–3852 | |||
| Frag2-SF1 | Seq PCR | AAT AAG AGC TTC AAA GAC TAT TGC AGT | 2602–2628 | |||
| Frag2-SF2 | Seq PCR | TAT ATC TAT CTT TAC AGA TGA AAT AAA ATT | 3252–3281 | |||
| Fragment 3 | Frag3-F1 | First PCR | CAG TGA AGG TGC ACA CAG ATG | 3452–3472 | First round 1792 bp | 59 °C |
| Frag3-R1 | First, second and Seq PCR | TAC CAC ACG TCG AAC GTG G | +44–+62 | |||
| Frag3-F2 | Second and Seq PCR | TTA TGC AAC ATA TAT AGC AAC ACC AT | 3772–3797 | second round 1472 bp | 56 °C | |
| Frag3-SF1 | Seq PCR | TTA TTT TGT CCA TCT TAG GGC TC | 4502–4524 | |||
| CN detection | ||||||
| Reference gene | Pvtubulin–FP | Real-Time PCR | CAA GAA CTC CTC CTA CTT CGT CG | 1644–1666 | 122 bp | 58 °C * |
| Pvtubulin–RP | Real-Time PCR | GTT GCG TGG AAA GCC ATC TC | 1746–1765 | |||
| Pvtubulin–P | Probe | HEX-TGCCCAACAGGGAGGAAGCGATT-BHQ1 | 1699–1721 | |||
| Target gene | PvMRP-FP | Real-Time PCR | AAA GTG TGT TTC CAG ACA AGA GTT | 1762–1785 | 111 bp | 58 °C |
| PvMRP-RP | Real-Time PCR | CAA ATT GCT TCG CTC CTC TG | 1853–1872 | |||
| PvMRP-P | Probe | FAM-TGCTCCAATGGCGGCAGTAGTAGTAG-BHQ1 | 1789–1814 | |||
| NO. | AA Change | Effect | Number (%) | Frequency (%) | |||
|---|---|---|---|---|---|---|---|
| East Asia (n = 71) | Southeast Asia (n = 19) | South Asia (n = 2) | Oceania (n = 2) | ||||
| 1 | N67S | non-synonymous mutation | 1 (1.06) | 1.4 | |||
| 2 | K36Q | non-synonymous mutation | 1 (1.06) | 1.4 | |||
| 3 | T234M | non-synonymous mutation | 18 (19.15) | 1.4 | 89.5 | ||
| 4 | T259R | non-synonymous mutation | 92 (97.87) | 100 | 100 | 100 | |
| 5 | F271Y | non-synonymous mutation | 1 (1.06) | 50 | |||
| 6 | R281K | non-synonymous mutation | 11 (11.70) | 15.5 | |||
| 7 | T282M | non-synonymous mutation | 1 (1.06) | 50 | |||
| 8 | S354N | non-synonymous mutation | 9 (9.57) | 12.7 | |||
| 9 | F406L | non-synonymous mutation | 1 (1.06) | 1.4 | |||
| 10 | K448I | non-synonymous mutation | 1 (1.06) | 1.4 | |||
| 11 | E533 | gene deletion | 3 (3.19) | 4.2 | |||
| 12 | F560I | non-synonymous mutation | 1 (1.06) | 50 | |||
| 13 | N568 | synonymous mutation | 2 (2.13) | 50 | 50 | ||
| 14 | E787D | non-synonymous mutation | 30 (31.9) | 42.3 | |||
| 15 | C817 | synonymous mutation | 1 (1.06) | ||||
| 16 | A853 | synonymous mutation | 20 (21.28) | 28.2 | |||
| 17 | V879 | synonymous mutation | 17 (18.09) | 89.5 | |||
| 18 | Q906E | non-synonymous mutation | 59 (62.77) | 56.3 | 94.7 | 50 | |
| 19 | G949D | non-synonymous mutation | 27 (28.72) | 38.0 | |||
| 20 | E967G | non-synonymous mutation | 1 (1.06) | 5.3 | |||
| 21 | L1207I | non-synonymous mutation | 17 (18.09) | 89.5 | |||
| 22 | K1219N | non-synonymous mutation | 2 (2.13) | 2.8 | |||
| 23 | I1232 | synonymous mutation | 38 (40.43) | 28.2 | 89.5 | 50 | |
| 24 | Y1393D | non-synonymous mutation | 92 (97.87) | 98.6 | 100 | 50 | 100 |
| 25 | V1360 | non-synonymous mutation | 9 (9.57) | 12.7 | |||
| 26 | G1419A | non-synonymous mutation | 1 (1.06) | 50 | |||
| 27 | V1478I | non-synonymous mutation | 90 (95.74) | 95.8 | 100 | 50 | 100 |
| 28 | H1586Y | non-synonymous mutation | 29 (30.85) | 38.0 | 5.3 | 50 | |
| Mutations | CQ Sensitive (n) a | CQ Resistant (n) a | p-Value * | |
|---|---|---|---|---|
| E787D | D | 16 | 1 | 0.2491 |
| E | 22 | 0 | ||
| G949D | D | 15 | 1 | 0.2245 |
| G | 23 | 0 | ||
| Q906E | E | 20 | 1 | 0.3483 |
| Q | 18 | 0 | ||
| H1586Y | Y | 14 | 1 | 0.2 |
| H | 24 | 0 | ||
| S354N | N | 6 | 1 | 0.0303 * |
| S | 32 | 0 | ||
| Region | n | S | η | H | Hd | π | k | dn/ds | Tajima’s D | p-Value | Fu and Li’s D* | p-Value |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| South America | 53 | 24 | 25 | 28 | 0.9544 | 0.0009 | 3.4933 | 0.7748 | −1.1743 | p > 0.1 | −2.0414 | 0.05 < p < 0.1 |
| East Asia | 72 | 11 | 11 | 15 | 0.8581 | 0.0008 | 3.2021 | 0.5630 | 1.1341 | p > 0.1 | 0.8084 | p > 0.1 |
| Southeast Asia | 30 | 15 | 15 | 8 | 0.4180 | 0.0006 | 2.0172 | 0.6154 | −1.5484 | p > 0.1 | −2.0390 | 0.05 < p< 0.1 |
| South Asia | 4 | 10 | 10 | 4 | 1.0000 | 0.0012 | 5.0000 | 0.4009 | 0.2982 | p > 0.1 | 0.2982 | p > 0.1 |
| Oceania | 6 | 10 | 10 | 6 | 1.0000 | 0.0009 | 3.3333 | 0.6762 | −0.9837 | p > 0.1 | −1.1374 | p > 0.1 |
| Africa | 1 | 6 | 6 | 1 | 1.0000 | 0.0015 | 6.0000 | - | - | - | - | - |
| Total | 166 | 34 | 35 | 46 | 0.9290 | 0.0011 | 3.6571 | 0.5536 | −1.1863 | p > 0.1 | −3.9871 | p < 0.02 |
| Population | East Asia | South America | Southeast Asia | South Asia | Oceania |
|---|---|---|---|---|---|
| East Asia (n = 72) | 0.0000 | ||||
| South America (n = 53) | 0.3788 | 0.0000 | |||
| Southeast Asia (n = 30) | 0.4832 | 0.5498 | 0.0000 | ||
| South Asia (n = 4) | 0.1957 | 0.0809 | 0.3682 | 0.0000 | |
| Oceania (n = 6) | 0.2350 | 0.1589 | 0.5351 | 0.1661 | 0.0000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yin, Y.; Chen, G.; Nyunt, M.H.; Zhang, M.; Liu, Y.; Zhu, G.; He, X.; Tian, F.; Cao, J.; Han, E.-t.; et al. Prevalence of pvmrp1 Polymorphisms and Its Contribution to Antimalarial Response. Microorganisms 2022, 10, 1482. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10081482
Yin Y, Chen G, Nyunt MH, Zhang M, Liu Y, Zhu G, He X, Tian F, Cao J, Han E-t, et al. Prevalence of pvmrp1 Polymorphisms and Its Contribution to Antimalarial Response. Microorganisms. 2022; 10(8):1482. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10081482
Chicago/Turabian StyleYin, Yi, Gangcheng Chen, Myat Htut Nyunt, Meihua Zhang, Yaobao Liu, Guoding Zhu, Xinlong He, Fang Tian, Jun Cao, Eun-taek Han, and et al. 2022. "Prevalence of pvmrp1 Polymorphisms and Its Contribution to Antimalarial Response" Microorganisms 10, no. 8: 1482. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10081482