
Current Advances in Biodegradation of Polyolefins

1
Frontier Science Center for Synthetic Biology and Key Laboratory of Systems Bioengineering (Ministry of Education), School of Chemical Engineering and Technology, Tianjin University, Tianjin 300072, China
2
Collaborative Innovation Center of Chemical Science and Engineering (Tianjin), Tianjin University, Tianjin 300072, China
*
Author to whom correspondence should be addressed.
Microorganisms 2022, 10(8), 1537; https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10081537
Submission received: 29 June 2022
/
Revised: 22 July 2022
/
Accepted: 27 July 2022
/
Published: 29 July 2022
(This article belongs to the Special Issue Biodegradation and Environmental Microbiomes)
Abstract
:Polyolefins, including polyethylene (PE), polypropylene (PP) and polystyrene (PS), are widely used plastics in our daily life. The excessive use of plastics and improper handling methods cause considerable pollution in the environment, as well as waste of energy. The biodegradation of polyolefins seems to be an environmentally friendly and low-energy consumption method for plastics degradation. Many strains that could degrade polyolefins have been isolated from the environment. Some enzymes have also been identified with the function of polyolefin degradation. With the development of synthetic biology and metabolic engineering strategies, engineered strains could be used to degrade plastics. This review summarizes the current advances in polyolefin degradation, including isolated and engineered strains, enzymes and related pathways. Furthermore, a novel strategy for polyolefin degradation by artificial microbial consortia is proposed, which would be helpful for the efficient degradation of polyolefin.
1. Introduction
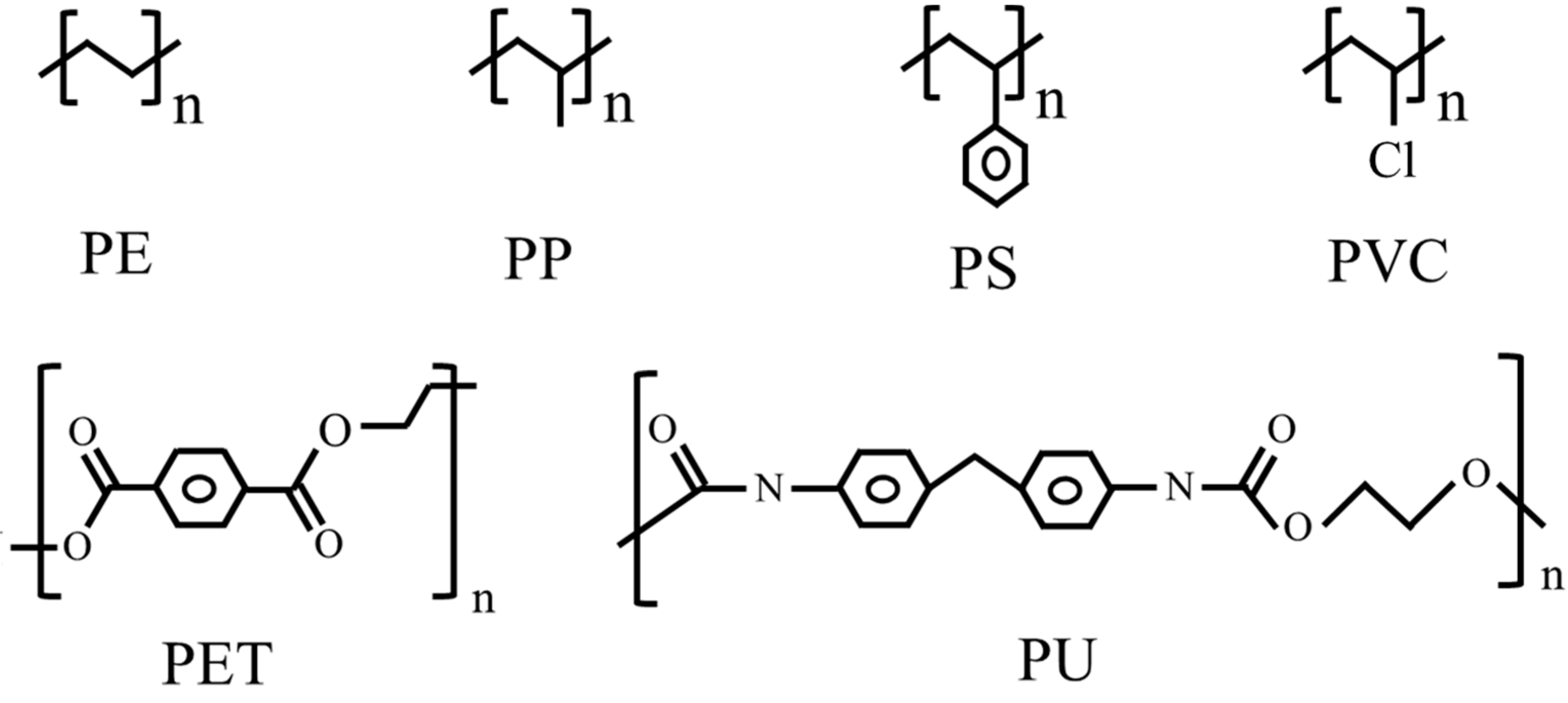
Plastics are one of the most widely used and vital materials in the modern world. However, the enormous manufacture and abuse of plastics place a huge burden on the environment. It was reported that about 58% of plastic waste was placed in landfills or discharged directly into the environment, 24% was burned and only 18% of plastic waste was recycled globally [1]. Some researchers have estimated that there would be roughly 12,000 Mt of plastic waste in the natural environment by 2050 [2]. Petroleum-derived polymers (polyethylene (PE), polyethylene terephthalate (PET), polyurethane (PU), polystyrene (PS), polypropylene (PP) and polyvinyl chloride (PVC)), especially polyolefins (PE, PP and PS) are widely used for packaging materials, plastic carry bags, plastic films, packaging foam, disposable cups and food containers [3] and are extremely recalcitrant to natural biodegradation [4]. As shown in Figure 1, polyolefins are linked by C–C and C–H bonds, the bond energy of which is much higher than that of C–O and C–N bonds, which means that polyolefins are more recalcitrant to degradation than plastics that consist of ester bonds, such as PET and PU [5]. There are different types of PE, including HDPE (high-density polyethylene), LDPE (low-density polyethylene) and LLDPE (liner low-density polyethylene). It is more difficult to degrade HDPE than LDPE or LLDPE due to its high density and crystallinity [1,2]. Moreover, because of its branches, PP is more difficult to degrade than PE. The benzene ring in PS made it more difficult to degrade than PE and PP. Although chemical treatment and thermal pyrolysis have been reported for polyolefin degradation, the use of harsh reaction conditions and toxic organic solvents can lead to secondary pollution in the environment. In recent years, many types of PE- and PS-degrading living species (mainly microbes) have been identified. These findings provide us with a mild and ecofriendly method for the degradation of polyolefins. Although the biodegradation efficiency of polyolefin is not high, polyolefin biodegradation is a promising method of recycling polyolefins, since various biotechnologies are being developed rapidly [5].
It was reported that the degradation rate of PP and PS was improved after pretreatment by UV and high temperature [6]. Therefore, many researchers have pretreated plastics with UV or high temperature prior to biodegradation. Some microorganisms, including pure strains and microbial consortia, have been isolated from the environment for the degradation of polyolefins. In 2006, Sivan et al. [7] isolated Rhodococcus ruber C208, which adhered to PE films and utilized LDPE films as the sole carbon source. Later, researchers found that R. ruber C208 could secrete laccase in vitro and that laccase also play a vital role in PE degradation [8]. In addition to laccase, manganese peroxidase [9], soybean peroxidase [10] and alkane hydroxylase [11,12] are also capable of degrading PE. Few PP-degrading enzymes have been reported to date. Hydroquinone peroxidase, which is produced by Azotobacter beijerinckii HM121, was reported to degrade PS [13]. Moreover, many microbial consortia have been isolated from nature with the capacity to degrade polyolefins. Some microbial consortia can degrade polyolefins into long-chain aliphatic compounds, which include alkanes, alkenes, alcohols and acids [14,15]. Many alkane hydroxylases that can degrade aliphatic alkanes have been identified, as well as alkene mono-oxygenases, which can degrade short-chain alkenes [16,17,18,19,20]. These researchers have provided a basis for the construction of artificial microbial consortia for the highly efficient degradation of polyolefins.
This review summarizes natural strains, enzymes and engineered microbial chassis for the biodegradation of polyolefins. Based on the former content, we provide an idea for polyolefin degradation by artificial microbial consortia and discussed the prospect.
2. Biodegradation of Polyolefins
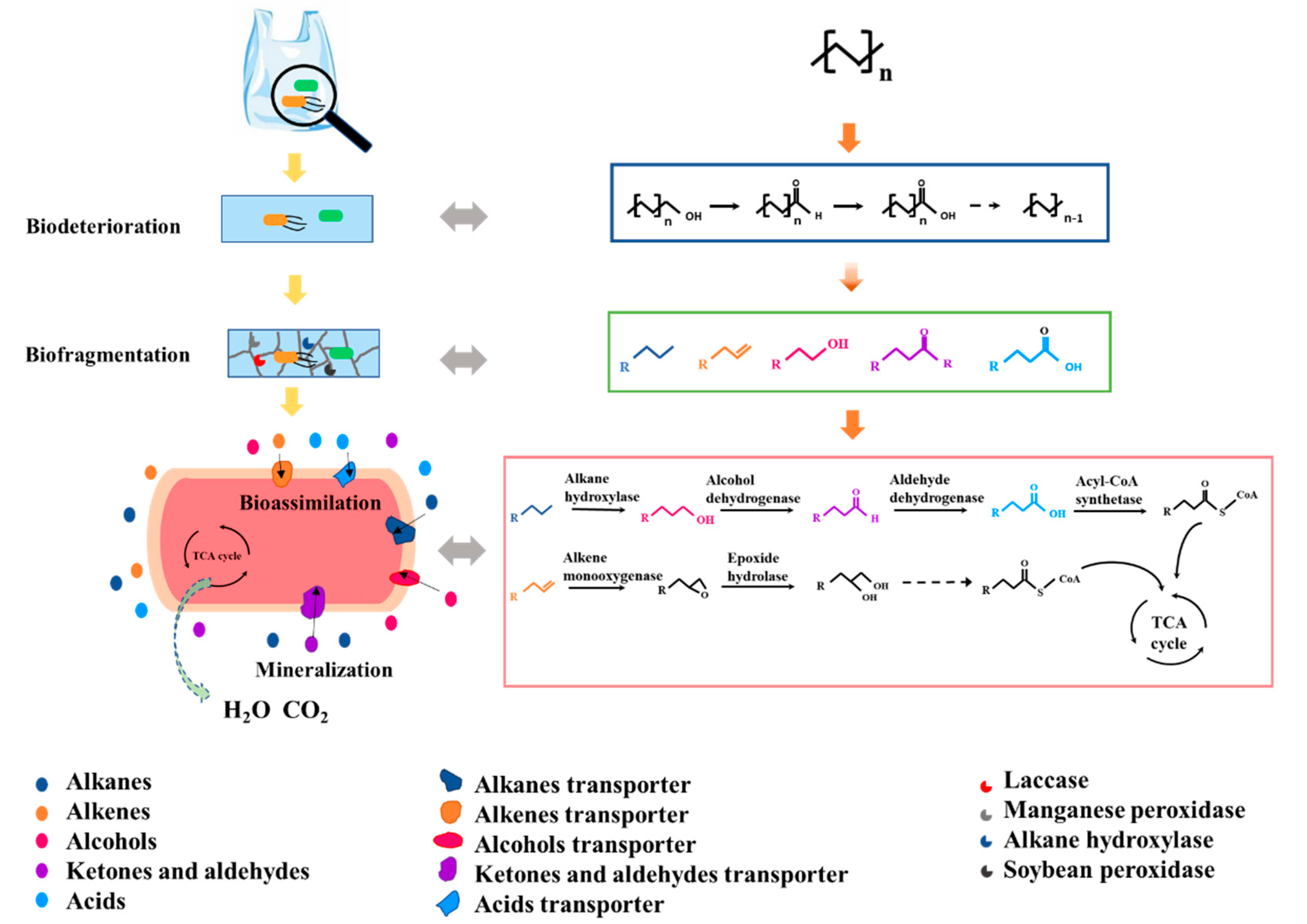
Polyolefins, including PE, PP and PS, are composed of C–C and C–H bonds, which are more stable against degradation than ester bonds. During polyolefin biodegradation, C–C and C–H bonds are oxidized. Many types of microorganisms have been isolated from sea water, compost and activated sludge with the capacity for polyolefin biodegradation. The oxidation of polyolefins can be divided into four stages, including biodeterioration, biofragmentation, bioassimilation and mineralization [4]. As shown in Figure 2, (taking PE as an example), degradation begins with the formation of biofilms [3]. In the biodeterioration stage, the surface of polyolefins is initially oxidized by the action of oxidative enzymes released by microorganisms or induced by exterior agents, such as sunlight (ultraviolet) exposure [4,5]. Biodeterioration reduces the number of carbonyl-groups and turns them into carboxylic acids, facilitates the further oxidation of polyolefins. During the biofragmentation stage, the polymer carbon chains are hydrolyzed into fragments with the release of intermediate products, which includes long-chain aliphatic compounds, such as alkanes and alkenes. The enzymes (e.g., laccase, manganese peroxidase and alkane hydroxylase) secreted by microorganisms capable of oxidizing polyolefins are involved in the biofragmentation stage [5]. Small hydrocarbon fragments with 10–50 carbon atoms released by biofragmentation are taken up and metabolized by microorganisms in the bioassimilation stage [21]. The hydrolysis products are transferred within the cell and degraded by the enzymes shown in Figure 2 and converted to microbial biomass with the associated release of carbon dioxide and water in the mineralization stage [4,5].
2.1. Isolated Microorganisms
Numerous of microorganisms with the capacity for polyolefin degradation, including bacteria, fungi and microbial consortia, have been isolated from the environment, such as soil containing plastic waste, the ocean and the guts of plastic-eating worms. These microorganisms are capable of utilizing polyolefins as sole carbon source or can generate depolymerases involved in polyolefin degradation. Therefore, the screening of these microorganisms is vital for the further degradation of polyolefins. Polyolefin-degrading bacteria can be identified by the following procedure. Soil samples collected from an area containing polyolefin waste was mixed with water and shaken [7]. In order to isolate strains from waxworm guts, worm gust need to be isolated and suspended in water [22]. Then, a portion of the suspension was transferred to polyolefin-containing medium. After incubation for several days, the polyolefin-degrading strains would be isolated through gradient dilution the of medium or collecting the strains on the agar media on which PE fragments were spread [22,23,24].
2.1.1. Single Bacteria
At present, most single strains that are capable of degrading polyolefins are bacteria, which can form biofilms on the surface of polyolefins or destroy the surface of polyolefins. Table 1 summarizes some bacteria capable of degrading polyolefins, including R. ruber [8,25,26], Pseudomonas [11,27,28], Bacillus [23,29], Acinetobacter [30] etc. R. ruber C208, one of the most efficient bacteria for PE biodegradation, is a Gram-positive bacterium isolated from soil and is capable of degrading unpretreated LDPE at a rate of 0.9% per week [7,31]. An increased rate of PE degradation was detected after LDPE was pretreated with UV light [32]. Changes in the molecular weight and molecular number of LDPE were also detected after incubation with R. ruber C208 [8]. It was reported that R. ruber C208 is also capable of degrading PS, achieving a weight loss of 0.8% after 8 weeks [25]. Moreover, R. rhodochrous ATCC 29672 exhibited the ability to degrade PP based on the characterization of changes in the metabolic activity of bacteria, such as ATP content, ADP/ATP ratio and cell viability [33].
As shown in Table 1, in addition to bacteria, fungi have the capacity to degrade polyolefin. Generally, the polyolefin-degrading capacity of fungi is better than that of bacteria, as fungi can generate hydrophobins, which can strengthen the fungal contact with the substrate and enable fungi to use polyolefins as carbon source [34]. Aspergillus clavatus JASK1, Phanerochaete chrysosporium NCIM 1170, Engyodontium album MTP091 and Curvularia sp. isolated from landfill soil have also been shown to play an important role in PE biodegradation [35,36,37].
Biodegradation should not only be determined by weight loss. Chemical properties, as well as changes in Mw (molecular weight) and Mn (molecular number), which can be characterized Fourier transform infrared spectroscopy (FTIR), X-ray photoelectron spectroscopy (XPS), nuclear magnetic resonance (NMR) and gel permeation chromatography (GPC) analyses, are also vital for the measurement of the degree of polyolefin degradation. After the biodeterioration stage, the nature and occurrence of functional groups on the surface of polyethylene substrates are changed, which can be studied by FTIR [21] spectroscopy, XPS [23] and NMR [38]. After the biofragmentation stage, the length of polyolefins is shorted. The result of biofragmentation can be proven by using GPC to measure changes in Mw and Mn [39]. After the biofragmentation stage, degradation products (such alkanes, alkenes, etc.) can be determined by GC-MS (gas chromatography mass spectrometry) [40]. Moreover, the result of bioassimilation can be determined through measurement of dry biomass weight of polyethylene-containing media. Finally, the percent of mineralization can be analyzed by CO2 measurement [28]. Furthermore, scanning electron microscopy (SEM) is another vital method used to characterize the surface features of polyolefins.
As previously reported, some waxworms can chew and eat plastics [23,41,42,43,44]. Enterobacter asburiae YT1 and Bacillus sp. YP1 were isolated from the guts of waxworms with the capability of degrading PE. Over 28-day incubation of the two strains on PE films, the physical properties (tensile strength and surface topography), chemical structure (hydrophobicity and appearance of carbonyl groups), Mw (accompanied by the formation of daughter products) and weight loss were detected [23]. Kyaw et al. [11] incubated Pseudomonas aeruginosa PAO1 with LDPE films. After exposure to P. aeruginosa PAO1, the LDPE sample turned into a mixture of long-chain fatty acids, esters, hydrocarbons, oxygenated chemical compounds predominantly containing aldehydes, ketones, esters and ether groups, unsaturated fatty acids and certain unknown compounds. Given that the structure of polyolefins is similar to that of alkanes, some strains with an ability to degrade alkanes also have an effect on PE degradation. Alcanivorax borkumensis, a bacterial strain isolated from the sea that can utilize alkane as carbon source [45] was found to induce a weight loss of 3.5% in 7 days [46].
2.1.2. Microbial Consortia
In addition to single bacteria, there are many microbial consortia that are capable of degrading PE isolated from various environments (Table 2). A mixed microbial consortium consisting of two Bacillus sp. and two Paenibacillus sp. was isolated from a landfill site. After incubation with the mixed microbial consortium for 30 days, the weight and the mean diameter of the PE sample were reduced 16.7% and 22.8%, respectively [29]. A microbial consortium consisting of Lysinibacillus xylanilyticus and Aspergillus niger was found to be capable of degrading PE [47]. After UV irradiation, the mineralization percentage increased from 15.8% to 29.5% after incubation for 126 days in soil, and according to FTIR and XRD, the chemical properties also improved. Skariyachan et al. [48] isolated a microbial consortium comprising Brevibacillus sps. and Aneurinibacillus sp. from waste landfills. During a 140-day incubation of the two strains on PE samples, weight loss for LDPE and HDPE strips reached 58.2% and 46.6% respectively, and weight loss for LDPE and HDPE pellets were 45.7% and 37.2%, respectively. As with a single bacterium, UV pretreatment aided in the degradation of polyolefin samples. In addition, the result of GC-MS analysis indicated that PE samples were degraded into cis-2-chlorovinylacetate, tri-decanoic acid and octadecanoic acid. Muenmee et al. [14] pretreated HDPE, LDPE, PP and PS samples with UV for 200 h, then mixed the plastics together. To simulate a landfill environment, the plastics mixture was placed in simulated lysimeters with a synthetic landfill gas (60% CH4:40% CO2) and a microbial consortium composed of Methylocyctis sp., Methylocella sp., Methylobactor sp., Methyloccus capsulatus., Nitrosomonas sp., Nitrosomonas europaea, Nitrobacter winogradskyi, Nitrobacter hamburgensis, Burkholderia sp., Pseudomonas sp. and Xanthobacter sp. After a reaction period of 3 months under semi-aerobic landfill conditions where different aeration rates were supplied, the plastic samples were degraded into hydrocarbon and oxygenated compounds, such as aliphatic alkanes, alkenes, alcohols and esters. The degradation products of different plastic types, such as the degraded products of HDPE, are mainly alkanes (C24H50, C32H66), alkenes (C15H30, C19H38) and some alcohols (C15H32O, C20H42O) were found, while for LDPE, only one alcohol (C11H24O) was found.

{kind=link}
{kind=link}
{kind=link}
Table 1.
Wild strains capable of polyolefin degradation.
| Polyolefin | Phylum/Class | Microorganism | Microorganism Source | Pretreatment | Experimental Condition | Biodegradation Result | Reference |
|---|---|---|---|---|---|---|---|
| PE | Bacteria | ||||||
| Terrabacteria group/Actinobacteria | R. ruber C208 | PE agricultural waste in soil | Unpretreated LDPE film | Incubation for 8 weeks at 37 °C | Weight loss: 7.5% | [7] | |
| Terrabacteria group/Actinobacteria | R. ruber C208 | PE agricultural waste in soil | UV-pretreated LDPE film | Incubation for 4 weeks at 30 °C | Weight loss: 8% | [32] | |
| Terrabacteria group/Actinobacteria | R. ruber C208 | PE agricultural waste in soil | Unpretreated LDPE film | Incubation for 30 days at 30 °C | Weight loss: 1.5–2.5%; reduction of 20.0% in Mw and 15.0% in Mn | [8] | |
| Terrabacteria group/Actinobacteria | Rhodococcus sp. | Three forest soils | Preoxidized LDPE film | Incubation for 30 days at 25 °C | Confirmation of adherence | [24] | |
| Terrabacteria group/Firmicutes | Staphylococcus arlettae | Various soil environments | Unpretreated PE film and PE powder | Incubation for 30 days at 37 °C | Weight loss: 13.6% | [22] | |
| Proteobacteria/Gammaproteobacteria | Enterobacter asburiae YT1 and Bacillus sp. YP1 | Guts of plastic-eating waxworms | Unpretreated LLDPE film | Shaken flasks incubated for 60 days at 30 °C | Weight losses of 6.1% and 10.7% after incubation with E. asburiae YT1 and Bacillus sp. YP1, respectively | [23] | |
| Proteobacte-ria/Gammaproteobacteria | Stentrophomonas sp. | Plastic debris in soil | Unpretreated LDPE film | Incubation for 30 days at 28 °C | Change in chemical properties | [49] | |
| Proteobacte-ria/Gammaproteobacteria | Stentrophomonas pavanii | Solid waste dump site | Modified LDPE | Incubation for 56 days at 30 °C | Confirmed by FTIR | [50] | |
| Proteobacte-ria/Gammaproteobacteria | Serratia marcescens | Soil | LLDPE powder made of LLDPE film | Incubation for 70 days at 30 °C | Weight loss: 36.0% | [51] | |
| Proteobacte-ria/Gammaproteobacteria | Alcanivorax borkumensis | Mediterranean Sea | Unpretreated LDPE film | Incubation for 7 days at 30 °C | Weight loss: 3.5% | [46] | |
| Terrabacteria group/Actinobacteria | Streptomyces spp. | Nile River Delta | 30 °C heat-treated degradable PE film | Incubation for 1 month at 30 °C | Three species showed slight weight loss. | [52] | |
| Terrabacteria group/Firmicutes | Pseudomonas aeruginosa PAO1 | ATCC | Unpretreated LDPE film | Incubation for 120 days at 37 °C | Maximum weight loss: 20.0% | [11] | |
| Terrabacteria group/Firmicute Dikarya/Ascomycota | Pseudomonas, Bacillus, Brevibacillus, Cellulosimicrobium, Lysinibacillus and Aspergillus | Dump site | Unpretreated PE films | Incubation for 16 weeks in shaken flasks at 37 °C and 28 °C | Gravimetric weight reductions of up to 36.4 % and 35.7% recorded for Aspergillus sp. and Bacillus sp. isolates, respectively. | [53] | |
| Fungi | |||||||
| Dikarya/Ascomycota | Aspergillus clavatus JASK1 | Landfill soil | Unpretreated LDPE films (bags) | Shaken flasks incubated for 90 days | Weight loss: 35.0% | [35] | |
| PP | Bacteria | ||||||
| Proteobacte-ria/Gammaproteobacteria | Stenotrophomonas panacihumi PA3–2 | Soil | Unpretreated PP powder | Incubation for 90 days at 37 °C | Mw decreased | [54] | |
| Terrabacteria group/Actinobacteria | R. rhodochrous ATCC 29672 | ATCC | PP film with pro-oxidant (Mn, Mn/Fe or Co) additives | Incubation for 180 days | Changes in ATP levels | [33] | |
| Terrabacteria group/Firmicute | Bacillus flexus | A soil consortium enriched from a plastic dumping site | UV-pretreated PP film | Incubation for 1 year | Weight loss: 2.5% | [55] | |
| Terrabacteria group/Firmicute | Bacillus cereus | Mangrove sediments | UV-pretreated PP granules | Incubation for 40 days at 3 °C | Weight loss: 12.0% | [56] | |
| Sporosarcina globispora | Mangrove sediments | UV-pretreated PP granules | incubation for 40 days at 33 °C | Weight loss: 11.0% | [56] | ||
| Terrabacteria group/Firmicute | Bacillus sp. | Municipal compost waste | Unpretreated PP powder | Incubation for 15 days at 37 °C | Weight loss: 10.0–12.0% | [57] | |
| Fungi | |||||||
| Proteobacteria /Gammaproteobacteria | Phanerochaete chrysosporium NCIM 1170, Engyodontium album MTP091 | 100 °C or UV for 10 days | Shaken flasks incubated for 12 months | Weight loss: 18.8% and 9.4% with P. chrysosporium and E. album, respectively | [37] | ||
| PS | Bacteria | ||||||
| Terrabacteria group/Firmicute | Exiguobacterium sp. strain YT2 | Guts of the larvae of Tenebrio molitor Linnaeus | Unpretreated styrofoam PS films | Incubation for 60 days | Weight loss: 7.4 % Mw decrease: 11.0% | [58] | |
| Terrabacteria group/Firmicute | Pseudomonas sp. | Soil | Unpretreated high-impact PS films | Incubation for 30 days at 30 °C | Weight loss: more than 10.0% | [59] | |
| Terrabacteria group/Firmicute | Bacillus sp. | Soil | Unpretreated high-impact PS films | Incubation for 30 days at 30 °C | Weight loss: 23.7% | [59] | |
| Terrabacteria group/Firmicute | Pseudomonas aeruginosa | Degraded polymer nanocomposite | PS: PLA and PS: PLA:organically modified montmorillonite (OMMT) composites | Incubation for 28 days at 30 °C in MSM | 9.9% degradation at 10 and 25% PS: PLA composites | [60] | |
| Terrabacteria group/Firmicute | Pseudomonas putida CA-3 | Industrial bioreactor | Pyrolyzed PS | 48 h of fermentation at 30 °C, 500 rpm | A single pyrolysis run and four fermentation runs resulted in the conversion of 64 g of PS to 6.4 g of PHA | [25] | |
| Terrabacteria group/Firmicute | Exiguobacterium sp. strain YT2 | Degraded plastic waste | High-impact PS | Incubation for 30 days at 30 °C | Weight loss: 12.4% | [61] | |
| Terrabacteria group/Actinobacteria | R. ruber C208 | Unpretreated styrofoam PS films | Incubation for 8 weeks at 28 °C | Weight loss: 0.8% | [62] | ||
| Fungi | |||||||
| Dikarya/Ascomycota | Curvularia sp. | Soil samples | Chemically oxidized PS | Incubation for 9 weeks at 30 °C | Microscopic examination showed adherence and penetrance to the polymer | [36] | |
2.2. Engineered Strains
2.2.1. Hydrolases Capable of Polyolefin Degradation
Most the enzymes capable of degrading PE are oxidoreductases. As shown in Table 3, well-known identified enzymes with the ability to oxidize polyolefins include laccase, manganese peroxidase, alkane hydroxylase and soybean peroxidase. Furthermore, a PS-degrading enzyme called hydroquinone peroxidase was identified from Azotobacter beijerinckii HM121. However, no PP-degrading enzymes have been identified to date. The result of enzymatic degradation of PE is that oxidation groups are introduced into the PE chain, which means polyolefins cannot be oxidized into monomers as the only act of these enzymes. Most PE-degrading enzymes can only perform terminal oxidation (the terminal carbon in polyolefins be oxidized) and subterminal oxidation (the carbon adjacent to the terminal carbon in polyolefins be oxidized) of PE. For example, laccase and manganese peroxidase can perform terminal oxidation, and the AlkB family can degrade n-alkanes, the main component of polyethylene, through either terminal or subterminal hydroxylation reactions [71]. Therefore, an ideal polyolefin-degrading enzyme has high hydroxylation activity against any carbon in the carbon chain so as to achieve the efficient transformation of polyolefin to its oligomer or monomer.
Laccases (EC 1.10.3.2), which belong to the so-called blue-copper family of oxidases, can catalyze the oxidation of a wide range of phenols and arylamines. Laccases, which are glycoproteins, have been reported in higher plants, fungi and bacteria. A laccase was purified from R. ruber C208 with the ability to degrade LDPE. As laccases contain four copper ion bonding sites, copper markedly affects their induction and activity, resulting in PE degradation. mRNA quantification by RT-PCR revealed a 13-fold increase in laccase mRNA levels in copper-treated cultures compared with an untreated control. The addition of copper to C208 cultures containing PE enhanced the biodegradation of PE by 75% [7,72]. A laccase mediator system (LMS) is composed of laccase and some small-molecule compounds that are easily oxidized by laccase, such as HBT, ABTS and DMP. In the process of LMS oxidation, laccase oxidizes the mediator first; then, the oxidized mediator oxidizes the substrate. It was reported that in the presence of a mediator, laccase can oxidize some substrates that it cannot oxidize alone. HBT, which has been used for PE degradation, reacts with non-phenolic models by a radical mechanism involving hydrogen atom abstraction [73]. Some researchers treated polyethylene with LMS using HBT (0.2 mM) as a mediator; after 3 days, the polyethylene membrane exhibited no elongation, and its relative tensile strength decreased by about 60%, which is higher than in the absence of HBT (20%) [74]. Johnnie et al. used laccase from Trichoderma viride fungus and 1-HBT to degrade LDPE. After incubation for 10 days, the weight loss of LDPE came to 2.3% [75].
Cytochrome P450 (CYP, P450), a member of a superfamily of heme–thiolate proteins, is distributed in most living organisms. There are more than 300,000 P450 genes. However, no P450 genes were found in E. coli, which means that E. coil is a good chassis for the heterologous expression of P450 genes. P450 enzymes can identify multiple substrates and catalyze diverse reactions, such as C–H hydroxylation; C=C double-bond epoxidation; heteroatom oxygenation; O-, N- and S-dealkylation; aromatic coupling; and C–C bond cleavage [76]. Because other PE-degrading enzymes can only perform terminal oxidation and subterminal oxidation of PE, the application of an ideal P450 enzyme that can cleave PE into short chains would contribute to the biodegradation of PE.
Alkane, which is composed of C–C bonds and C–H bonds, has a similar structure to that of PE. Therefore, alkane mono-oxygenase enzymes are potential candidates for the degradation of PE. One alkane hydroxylase, namely AlkB, has been reported to degrade PE [76]. AlkB, which was first identified in alkane-consuming Pseudomonas species isolated from oil-contaminated areas, is a membrane-bound, non-heme di-iron monooxygenase [77].
Manganese peroxidase was purified from a lignin-degrading fungus: P. chrysosporium. Manganese peroxidase was first identified as a lignin-degrading enzyme. It was reported that the addition of Mn (II) to nitrogen- or carbon-limited culture medium enhanced PE degradation [9].
2.2.2. Engineered Chassis for Polyolefin Biodegradation
As shown in Table 3, the microorganisms that are capable of secreting polyolefin biodegrading enzymes are not model organisms, which means that they are difficult to genetically engineer. Two model organisms, E. coli and Y. lipolytica, have been applied to the expression and secretion of polyolefin-biodegrading enzymes to date [12,78,79]. The heterologous expression of polyolefin-degrading enzymes in model organisms can efficiently increase the expression level of polyolefin-degrading enzymes through genetic engineering in model organisms.
E. coli:E. coli is one of the most widely used model microorganisms for production of recombinant proteins. As a model microorganism, E. coli has advantages in many aspects, such as a simple genetic background, ease of genetic modification and simple growth conditions. Engineered E. coli has been used to express alkane hydroxylase to degrade LMWPE. The alkB gene, which was cloned from Pseudomonas sp. E4, was introduced into E. coli BL21. After incubation for 80 days at 37 °C with engineered E. coli, 19.3% of the LMWPE was degraded [28]. The rubredoxin and rubredoxin reductase could help alkane monooxygenase to transfer electron. If the rubredoxin and rubredoxin reductase are co-expressed with alkane monooxygenase, the conversion rate can be increased. Researchers fused and expressed alkB with its coenzyme genes—rubA1, rubA2 and rub—in E. coli BL21. The result indicated that 30.5% of the carbon of LMWPE-1 degraded into CO2 after 78 days [80]. Another study revealed that an alkane-1-monooxygenase (AlkB) in Acinetobacter johnsonii JNU01 degraded PS, and this finding was later confirmed by recombinant alkB in E. coli BL21 [53]. These researchers also expressed alkB2 in E. coli, and the result indicates that alkB2 was more efficient for low-molecular-weight PE biodegradation than alkB1 [12]. A laccase gene isolated from a marine fungus was expressed in E. coli and showed PE degradation ability [81].
Many recent studies have shown that engineered E. coli can be used for laccase heterologous expression and secretion [82,83,84]. Ihssen et al. [78] expressed five novel bacterial laccase-like multicopper oxidases (LMCOs) of diverse origin. However, a potential issue with laccase expression in E. coli is that it is easy for E. coli to form inclusion bodies when expressing extracellular enzymes. Mo et al. [79] expressed three laccases from three different organisms, namely Lac1326 from marine sediment samples, fungal tvel5 laccase from Trametes versicolor and bacterial BPUL laccases from Bacillus pumilus for the purpose of degrading β-estradiol. The result of Western blot analysis indicates that laccase was detected both in vivo and in vitro in E. coli, which means that some laccase stayed in inclusion bodies instead of being secreted in vitro. Given that laccase has been widely used for the biodegradation of PE [38,75,85,86], the heterologous expression of laccase in E. coli is a potential method for the biodegradation of PE.
Yarrowia lipolytica:Y. lipolytica is a Crabtree-negative ascomycete yeast with good protein secretion capacities. Compared to other yeasts, Y. lipolytica lacks α-1,3-mannosyltransferase, a factor that limits the amount of excessive mannosylation of secreted heterologous glycoproteins and constitutes a valuable asset for the production of therapeutic proteins [87]. Y. lipolytica W29 is a wild-type strain with a remarkable characteristically high secretion level of proteins [88]. Y. lipolytica W29 ura302 was obtained through genetic convention of URA3 into ura3-302 in Y. lipolytica W29, and it was able to utilize sucrose and molasses as a carbon source under the control of XPR2 promoter. After the genetic convention of XPR2 into xpr2-322 and AXP1 into axp1-2, which indicates inactivation of alkaline extracellular protease and acid extracellular protease that would degrade foreign extracellular protein, a strain with high heterologous protein production capability called Y. lipolytica Po1f was obtained. A new strain called Y. lipolytica Po1g that carries a pBR322 docking platform was obtained through the integration of PINA300′ plasmid in Y. lipolytica Po1f [89]. Y. lipolytica Po1g was induced with a YLEX kit for expression/secretion of heterologous proteins [90]. A laccase from the white-rot fungus Trametes versicolor was expressed in Y. lipolytica Po1g for the biodegradation of PE. Compared to the yeast secretion signal, the native secretion signal showed higher enzyme activity in the culture medium. The yield of laccase reached 2.5 mg/L (0.23 units/mL) [72]. Laccase has been widely used for the biodegradation of PE, and the heterologous expression of laccase in Y. lipolytica is a promising method of PE degradation.
Table 3.
Enzymes capable of polyolefin degradation.
| Plastics | Enzymes | Enzyme Source | Pretreatment | Experimental Condition | Result | Reference |
|---|---|---|---|---|---|---|
| PE | Laccase | R. ruber C208 | Unpretreated LDPE film | Incubation for 30 days at 30 °C | Weight loss: 1.5–2.5%; reduction of 20% in Mw and 15% in Mn | [7] |
| Manganese peroxidase | Phanerochaete chrysosporium | Unpretreated PE film | Incubation for 12 days at 37 °C | Mw decreased | [9] | |
| Soybean peroxidase | Soybean | Unpretreated HDPE film | Reaction for 2 h at 60 °C | Hydrophilicity increased | [10] | |
| Alkane hydroxylase | Pseudomonas sp. E4 | Unpretreated LMWPE sheet | Incubation for 80 days at 37 °C | Weight loss: 19.3% | [28] | |
| Alkane hydroxylase | Pseudomonas aeruginosa E7(uniport Q9I0R2) | Unpretreated LMWPE film | Incubation for 50 days at 37 °C | Weight loss: 19.6–30.5% | [12] | |
| PS | Hydroquinone peroxidase | Azotobacter beijerinckii HM121 | Unpretreated PS film | Incubation for 20 min | Mw decreased | [13] |
| Alkane hydroxylase | A. johnsonii JNU01 | Unpretreated low-molecular-weight PS powder | Incubation for 7 days at 28 °C | Confirmed by FTIR and SEM | [72] |
3. Artificial Microbial Consortia in Polyolefin Biodegradation
Microorganisms do not exist independently in natural environments, and they usually live in complex communities. They communicate through quorum sensing and share metabolites and enzymes, expanding the substrate range compared with single bacteria. Therefore, artificial microbial consortia can be constructed to degrade plastics that are not easily degraded naturally [91]. The application of artificial microbial consortia for complex biological processes is an emerging field in synthetic biology. There are several specific advantages of plastic degradation by artificial microbial consortia compared to pure culture. Firstly, the synergies of different strains and different enzymic systems can improve the efficiency of polyolefin degradation and reduce the metabolic burden [92,93]. Moreover, the effect of polyolefin degradation by artificial microbial consortia can be more complete than that of pure culture. Finally, the construction of artificial microbial consortia is a time-saving and efficient method relative to other metabolic engineering techniques [79]. Therefore, constructing artificial microbial consortia is regarded as a promising means of polyolefin degradation. The bioproduction of surfactin by coculture of B. amyloliquefaciens MT45 and B. amyloliquefaciens X82 improved substrate utilization and increased the product titer by 3.3-fold [94]. Another microbial consortium including a crude oil degrader and biosurfactant producer was constructed to degrade crude oil, achieving 95.8% degradation efficiency of crude oil and degrading various hydrocarbons more effectively than single strains [95]. In our laboratory, we constructed a four-microbe consortium comprising two metabolically engineered B. subtilis, Rhodococcus jostii and P. putida to degrade PET. The result showed that the microbial consortium could degrade PET film, with weight loss reaching 23.2% under ambient temperature. The artificial microbial consortium successfully relieved the metabolic inhibition of TPA and EG [96,97]. Some researchers also proposed a novel strategy combining enzymic and microbial degradation of PET, achieving a maximum degradation efficiency of approximately 91.4% [98]. As for PET degradation, artificial microbial consortia provide a potential means of polyolefin degradation.
Most microbial consortia with the ability to degrade polyolefins identified to date are natural microbial consortia. Skariyachan et al. [66] formulated a bacterial consortium composed of strains isolated from plastic garbage processing areas. After incubation with the bacterial consortium, LDPE strips and LDPE pellets showed weight loss of 81.0% and 38.0%, respectively. Another artificially formulated microbial consortium composed of four strains isolated from cow dung samples gathered from highly plastic-acclimated environments degraded 75.0%, 55.0%, 60.0% and 43.0% of LDPE strips, LDPE pellets, HDPE strips and HDPE pellets, respectively, over a period of 120 days at 55 °C. The weight loss of PE degraded by microbial consortia is much higher than that of pure culture (10.2% at most) [65]. Later, these researchers formulated a bacterial consortium from cow dung samples for the purpose of LDPE and PP degradation. After incubation with the potential consortium (CB3) at 37 °C, LDPE and PP films showed degradation of 64.2% and 63.0%, respectively, which is much higher than that of pure culture of any strains in this microbial consortium [99]. However, artificially formulated microbial consortia still have disadvantages, such as division of labor, metabolic imbalance, competition, cooperation and complex interactions [100].
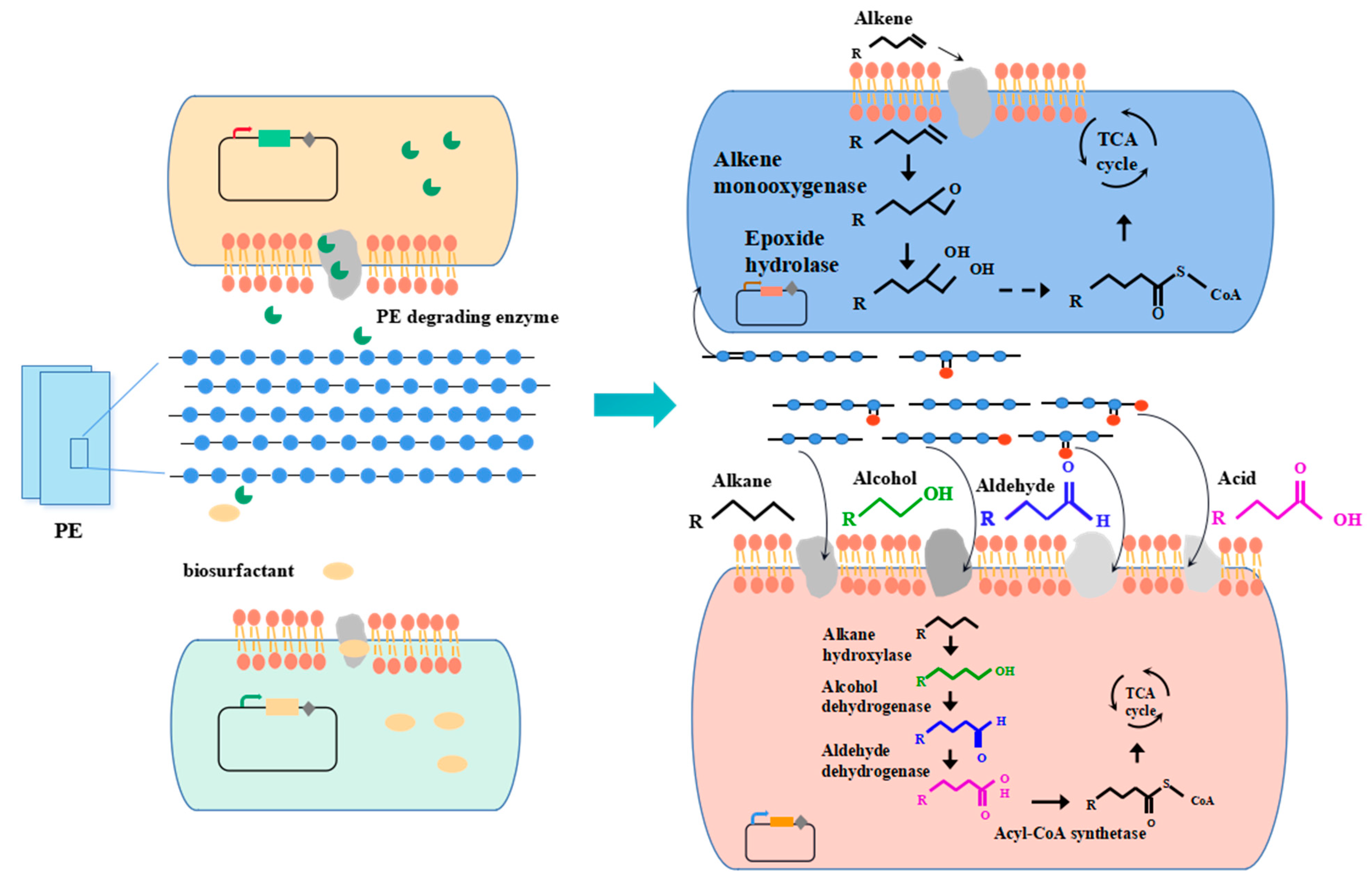
Most single bacteria or microbial consortia isolated from nature can only degrade polyolefins into long-chain aliphatic compounds (alkanes, alkenes, ketones, aldehydes, alcohols, acid, ketone acids, dicarboxylic acids and esters) at most. Few studies have achieved the goal of complete degradation of polyolefins. Building artificial microbial consortia provides a potential means for the complete degradation of polyolefins using polymer-degrading strains and long-chain aliphatic-compound-degrading strains. Combining polyolefin-degrading modules and long-chain aliphatic-compound-degrading module is promising means by which to completely degrade polyolefins (Figure 3). As shown in Figure 3 (taking PE as an example), an artificial microbial consortium that includes a biosurfactant producer [101], a polyolefin degrader [72], an alkene degrader and an alkane degrader [102] could be constructed for the biodegradation of polyolefins. In this artificial microbial consortium, the biosurfactant producer could produce biosurfactant, which can help the polyolefin degrader or polyolefin-degrading enzymes to make contact with polyolefins [15]. Then, some carbonyl groups in the carbon chain of polyolefin would be reduced under the action of the polyolefin-degrader or extracellular enzymes secreted by engineering strains. Subsequently, the polymer carbon chains would be hydrolyzed into fragments with the release of intermediate products, including alkane, alkene, ketones, aldehydes, alcohols, acids, etc. These intermediate products could be transported to alkene degraders and alkane degraders [21]. Alkanes could be gradually degraded into ketones, aldehydes, alcohols and acid and enter the tricarboxylic acid cycle (TCA cycle). As for alkenes, they could be degraded into epoxide, vicinal diol, aldehydes and acids and enter the metabolic pathway in the cell [5,76].
In contrast to polyolefins, long-chain aliphatic compounds can transfer into cells through active transport, passive transport, endocytosis and free diffusion and be metabolized in the cells. Many researchers have reported microorganisms isolated from environments that are capable of degrading alkanes, such as Aspergillus sp. [103], Bacillus sp. [104] and Yarrowia sp. [105]. Many enzymes have been identified for alkane degradation, such as alkane hydroxylase, which can oxidize alkanes into alcohol. Alcohols are then gradually oxidized into aldehydes, acids and acetyl-CoA and enter the tricarboxylic acid cycle (TCA cycle) or are converted into high-value chemicals, such as sophorolipid [101]. As for the degradation of alkene, the biodegrading enzymes are alkene monooxygenase, which can oxidize alkenes into epoxide [17]. Alkenes could be oxidized into vicinal diol [106] and then be oxidized into aldehydes, acids and acetyl-CoA and enter the TCA cycle.
Constructing an artificial microbial consortium that includes part or all of the following strains is a vital metabolic strategy to solve the current problems of polyolefin degradation: polyolefin degraders, long-chain aliphatic-compound degraders and biosurfactant producers. Artificial microbial consortia can perform more complex tasks compared to pure strains, and the division of labor is clearer than that of artificially formulated consortia.
4. Conclusions
In this review, we summarized the current advances in polyolefin biodegradation from pure natural strains, natural microbial consortia, enzymes, engineered chassis and artificial microbial consortia. The prospect of the biodegradation of polyolefins by artificial microbial consortia was also discussed. Constructing artificial microbial consortia is a promising strategy for the biodegradation of polyolefin. The use of artificial microbial consortia is a promising method for the degradation of polyolefins and long-chain aliphatic compounds. Compared to natural microbial consortia, the division of labor in artificial microbial consortia is clearer and more favorable for follow-up studies. The possibility of building an artificial microbial consortium including a biosurfactant producer, a polyolefin degrader, an alkene degrader and an alkane degrader was proposed in this review. There have been many recent advances in synthetic biology and metabolic engineering [107,108,109,110,111]. Therefore, it is possible to design rational and efficient artificial microbial consortia to degrade polyolefins, as well as more complex compounds.
Author Contributions
N.Z.: conceptualization, writing—original draft; M.D.: conceptualization, writing—review and editing, supervision; Y.Y.: supervision. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the National Key Research and Development Program of China (2019YFA0706900).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chamas, A.; Moon, H.; Zheng, J.J.; Qiu, Y.; Tabassum, T.; Jang, J.H.; Abu-Omar, M.; Scott, S.L.; Suh, S. Degradation Rates of Plastics in the Environment. ACS Sustain. Chem. Eng. 2020, 8, 3494–3511. [Google Scholar] [CrossRef] [Green Version]
- Geyer, R.; Jambeck, J.R.; Law, K.L. Production, use, and fate of all plastics ever made. Sci. Adv. 2017, 3, e1700782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellis, L.D.; Rorrer, N.A.; Sullivan, K.P.; Otto, M.; McGeehan, J.E.; Román-Leshkov, Y.; Wierckx, N.; Beckham, G.T. Chemical and biological catalysis for plastics recycling and upcycling. Nat. Catal. 2021, 4, 539–556. [Google Scholar] [CrossRef]
- Mohanan, N.; Montazer, Z.; Sharma, P.K.; Levin, D.B. Microbial and Enzymatic Degradation of Synthetic Plastics. Front. Microbiol. 2020, 11, 580709. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Pedersen, J.N.; Eser, B.E.; Guo, Z. Biodegradation of polyethylene and polystyrene: From microbial deterioration to enzyme discovery. Biotechnol. Adv. 2022, 60, 107991. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Jin, T.; Zou, T.; Xu, L.; Xi, B.; Xu, D.; He, J.; Xiong, L.; Tang, C.; Peng, J.; et al. Current progress on plastic/microplastic degradation: Fact influences and mechanism. Environ. Pollut. 2022, 304, 119159. [Google Scholar] [CrossRef]
- Sivan, A.; Szanto, M.; Pavlov, V. Biofilm development of the polyethylene-degrading bacterium Rhodococcus ruber. Appl. Microbiol. Biotechnol. 2006, 72, 346–352. [Google Scholar] [CrossRef]
- Santo, M.; Weitsman, R.; Sivan, A. The role of the copper-binding enzyme—Laccase—In the biodegradation of polyethylene by the actinomycete Rhodococcus ruber. Int. Biodeterior. Biodegrad. 2013, 84, 204–210. [Google Scholar] [CrossRef]
- Iiyoshi, Y.; Tsutsumi, Y.; Nishida, T. Polyethylene degradation by lignin-degrading fungi and manganese peroxidase. J. Wood Sci. 1998, 44, 222–229. [Google Scholar] [CrossRef]
- Zhao, J.C.; Guo, Z.; Ma, X.Y.; Liang, G.Z.; Wang, J.L. Novel surface modification of high-density polyethylene films by using enzymatic catalysis. J. Appl. Polym. Sci. 2004, 91, 3673–3678. [Google Scholar] [CrossRef]
- Kyaw, B.M.; Champakalakshmi, R.; Sakharkar, M.K.; Lim, C.S.; Sakharkar, K.R. Biodegradation of Low Density Polythene (LDPE) by Pseudomonas Species. Indian J. Microbiol. 2012, 52, 411–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeon, H.J.; Kim, M.N. Comparison of the functional characterization between alkane monooxygenases for low-molecular-weight polyethylene biodegradation. Int. Biodeterior. Biodegrad. 2016, 114, 202–208. [Google Scholar] [CrossRef]
- Nakamiya, K.; Ooi, T.; Kinoshita, S. Non-heme hydroquinone peroxidase from Azotobacter beijerinckii HM121. J. Ferment. Bioeng. 1997, 84, 14–21. [Google Scholar] [CrossRef]
- Muenmee, S.; Chiemchaisri, W.; Chiemchaisri, C. Enhancement of biodegradation of plastic wastes via methane oxidation in semi-aerobic landfill. Int. Biodeterior. Biodegrad. 2016, 113, 244–255. [Google Scholar] [CrossRef]
- Mukherjee, S.; RoyChaudhuri, U.; Kundu, P.P. Biodegradation of polyethylene via complete solubilization by the action of Pseudomonas fluorescens, biosurfactant produced by Bacillus licheniformis and anionic surfactant. J. Chem. Technol. Biotechnol. 2018, 93, 1300–1311. [Google Scholar] [CrossRef]
- Panasia, G.; Philipp, B. LaoABCR, a Novel System for Oxidation of Long-Chain Alcohols Derived from SDS and Alkane Degradation in Pseudomonas aeruginosa. Appl. Environ. Microbiol. 2018, 84, e00626-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brzeszcz, J.; Kaszycki, P. Aerobic bacteria degrading both n-alkanes and aromatic hydrocarbons: An undervalued strategy for metabolic diversity and flexibility. Biodegradation 2018, 29, 359–407. [Google Scholar] [CrossRef] [Green Version]
- Ensign, S.A. Microbial Metabolism of Aliphatic Alkenes. Biochemistry 2001, 40, 5845–5853. [Google Scholar] [CrossRef]
- Williams, S.C.; Forsberg, A.P.; Lee, J.; Vizcarra, C.L.; Lopatkin, A.J.; Austin, R.N. Investigation of the prevalence and catalytic activity of rubredoxin-fused alkane monooxygenases (AlkBs). J. Inorg. Biochem. 2021, 219, 111409. [Google Scholar] [CrossRef]
- Krum, J.G.; Ensign, S.A. Evidence that a Linear Megaplasmid Encodes Enzymes of Aliphatic Alkene and Epoxide Metabolism and Coenzyme M (2-Mercaptoethanesulfonate) Biosynthesis in Xanthobacter Strain Py2. J. Bacteriol. 2001, 183, 2172–2177. [Google Scholar] [CrossRef] [Green Version]
- Restrepo-Flórez, J.-M.; Bassi, A.; Thompson, M.R. Microbial degradation and deterioration of polyethylene—A review. Int. Biodeterior. Biodegrad. 2014, 88, 83–90. [Google Scholar] [CrossRef]
- Divyalakshmi, S.; Athikayala, S. Screening and isolation of polyethylene degrading bacteria from various soil environments. J. Environ. Sci. 2016, 12, 01–07. [Google Scholar] [CrossRef]
- Yang, J.; Yang, Y.; Wu, W.-M.; Zhao, J.; Jiang, L. Evidence of Polyethylene Biodegradation by Bacterial Strains from the Guts of Plastic-Eating Waxworms. Environ. Sci. Technol. 2014, 48, 13776–13784. [Google Scholar] [CrossRef] [PubMed]
- Koutny, M.; Amato, P.; Muchova, M.; Ruzicka, J.; Delort, A.-M. Soil bacterial strains able to grow on the surface of oxidized polyethylene film containing prooxidant additives. Int. Biodeterior. Biodegrad. 2009, 63, 354–357. [Google Scholar] [CrossRef] [Green Version]
- Ward, P.G.; Goff, M.; Donner, M.; Kaminsky, W.; O’Connor, K.E. A Two Step Chemo-biotechnological Conversion of Polystyrene to a Biodegradable Thermoplastic. Environ. Sci. Technol. 2006, 40, 2433–2437. [Google Scholar] [CrossRef]
- Jakubowicz, I. Evaluation of degradability of biodegradable polyethylene (PE). Polym. Degrad. Stab. 2003, 80, 39–43. [Google Scholar] [CrossRef]
- Nanda, S.; Smiti, S.S.; Jayanthi, A. Studies on the biodegradation of natural and synthetic polyethylene by Pseudomonas spp. J. Appl. Sci. Environ. Manag. 2010, 14, 57–60. [Google Scholar] [CrossRef] [Green Version]
- Yoon, M.G.; Jeon, H.J.; Kim, M.N. Biodegradation of Polyethylene by a Soil Bacterium and AlkB Cloned Recombinant Cell. J. Bioremediation Biodegrad. 2012, 3, 145. [Google Scholar] [CrossRef] [Green Version]
- Park, S.Y.; Kim, C.G. Biodegradation of micro-polyethylene particles by bacterial colonization of a mixed microbial consortium isolated from a landfill site. Chemosphere 2019, 222, 527–533. [Google Scholar] [CrossRef]
- Kim, H.-W.; Jo, J.H.; Kim, Y.-B.; Le, T.-K.; Cho, C.-W.; Yun, C.-H.; Chi, W.S.; Yeom, S.-J. Biodegradation of polystyrene by bacteria from the soil in common environments. J. Hazard. Mater. 2021, 416, 126239. [Google Scholar] [CrossRef]
- Fontanella, S.; Bonhomme, S.; Koutny, M.; Husarova, L.; Brusson, J.-M.; Courdavault, J.-P.; Pitteri, S.; Samuel, G.; Pichon, G.; Lemaire, J.; et al. Comparison of the biodegradability of various polyethylene films containing pro-oxidant additives. Polym. Degrad. Stab. 2010, 95, 1011–1021. [Google Scholar] [CrossRef]
- Gilan, I.; Hadar, Y.; Sivan, A. Colonization, biofilm formation and biodegradation of polyethylene by a strain of Rhodococcus ruber. Appl. Microbiol. Biotechnol. 2004, 65, 97–104. [Google Scholar] [CrossRef]
- Fontanella, S.; Bonhomme, S.; Brusson, J.-M.; Pitteri, S.; Samuel, G.; Pichon, G.; Lacoste, J.; Fromageot, D.; Lemaire, J.; Delort, A.-M. Comparison of biodegradability of various polypropylene films containing pro-oxidant additives based on Mn, Mn/Fe or Co. Polym. Degrad. Stab. 2013, 98, 875–884. [Google Scholar] [CrossRef]
- Sánchez, C. Fungal potential for the degradation of petroleum-based polymers: An overview of macro- and microplastics biodegradation. Biotechnol. Adv. 2020, 40, 107501. [Google Scholar] [CrossRef]
- Gajendiran, A.; Krishnamoorthy, S.; Abraham, J. Microbial degradation of low-density polyethylene (LDPE) by Aspergillus clavatus strain JASK1 isolated from landfill soil. 3 Biotech 2016, 6, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Motta, O.; Proto, A.; De Carlo, F.; Santoro, E.; Brunetti, L.; Capunzo, M. Utilization of chemically oxidized polystyrene as co-substrate by filamentous fungi. Int. J. Hyg. Environ. Health 2009, 212, 61–66. [Google Scholar] [CrossRef]
- Jeyakumar, D.; Chirsteen, J.; Doble, M. Synergistic effects of pretreatment and blending on fungi mediated biodegradation of polypropylenes. Bioresour. Technol. 2013, 148, 78–85. [Google Scholar] [CrossRef]
- Sarmah, P.; Rout, J. Efficient biodegradation of low-density polyethylene by cyanobacteria isolated from submerged polyethylene surface in domestic sewage water. Environ. Sci. Pollut. Res. 2018, 25, 33508–33520. [Google Scholar] [CrossRef]
- Montazer, Z.; Najafi, M.B.H.; Levin, D.B. Challenges with Verifying Microbial Degradation of Polyethylene. Polymers 2020, 12, 123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada-Onodera, K.; Mukumoto, H.; Katsuyaya, Y.; Saiganji, A.; Tani, Y. Degradation of polyethylene by a fungus, Penicillium simplicissimum YK. Polym. Degrad. Stab. 2001, 72, 323–327. [Google Scholar] [CrossRef]
- Ru, J.K.; Huo, Y.X.; Yang, Y. Microbial Degradation and Valorization of Plastic Wastes. Front. Microbiol. 2020, 11, 442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bombelli, P.; Howe, C.J.; Bertocchini, F. Polyethylene bio-degradation by caterpillars of the wax moth Galleria mellonella. Curr. Biol. 2017, 27, R292–R293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chalup, A.; Ayup, M.M.; Garzia, A.C.M.; Malizia, A.; Martin, E.; De Cristóbal, R.; Galindo-Cardona, A. First report of the lesser wax moth Achroia grisella F. (Lepidoptera: Pyralidae) consuming polyethylene (silo-bag) in northwestern Argentina. J. Apic. Res. 2018, 57, 569–571. [Google Scholar] [CrossRef]
- Kundungal, H.; Gangarapu, M.; Sarangapani, S.; Patchaiyappan, A.; Devipriya, S.P. Efficient biodegradation of polyethylene (HDPE) waste by the plastic-eating lesser waxworm (Achroia grisella). Environ. Sci. Pollut. Res. 2019, 26, 18509–18519. [Google Scholar] [CrossRef] [PubMed]
- Sabirova, J.S.; Ferrer, M.; Regenhardt, D.; Timmis, K.N.; Golyshin, P.N. Proteomic Insights into Metabolic Adaptations in Alcanivorax borkumensis Induced by Alkane Utilization. J. Bacteriol. 2006, 188, 3763–3773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delacuvellerie, A.; Cyriaque, V.; Gobert, S.; Benali, S.; Wattiez, R. The plastisphere in marine ecosystem hosts potential specific microbial degraders including Alcanivorax borkumensis as a key player for the low-density polyethylene degradation. J. Hazard. Mater. 2019, 380, 120899. [Google Scholar] [CrossRef] [PubMed]
- Esmaeili, A.; Pourbabaee, A.A.; Alikhani, H.A.; Shabani, F.; Esmaeili, E. Biodegradation of Low-Density Polyethylene (LDPE) by Mixed Culture of Lysinibacillus xylanilyticus and Aspergillus niger in Soil. PLoS ONE 2013, 8, e71720. [Google Scholar] [CrossRef] [Green Version]
- Skariyachan, S.; Patil, A.A.; Shankar, A.; Manjunath, M.; Bachappanavar, N.; Kiran, S. Enhanced polymer degradation of polyethylene and polypropylene by novel thermophilic consortia of Brevibacillus sps. and Aneurinibacillus sp. screened from waste management landfills and sewage treatment plants. Polym. Degrad. Stab. 2018, 149, 52–68. [Google Scholar] [CrossRef]
- Peixoto, J.; Silva, L.P.; Krüger, R.H. Brazilian Cerrado soil reveals an untapped microbial potential for unpretreated polyethylene biodegradation. J. Hazard. Mater. 2017, 324, 634–644. [Google Scholar] [CrossRef] [PubMed]
- Mehmood, C.T.; Qazi, I.A.; Hashmi, I.; Bhargava, S.; Deepa, S. Biodegradation of low density polyethylene (LDPE) modified with dye sensitized titania and starch blend using Stenotrophomonas pavanii. Int. Biodeterior. Biodegrad. 2016, 113, 276–286. [Google Scholar] [CrossRef]
- Azeko, S.T.; Etuk-Udo, G.A.; Odusanya, O.S.; Malatesta, K.; Anuku, N.; Soboyejo, W.O. Biodegradation of Linear Low Density Polyethylene by Serratia marcescens subsp. marcescens and its Cell Free Extracts. Waste Biomass-Valorization 2015, 6, 1047–1057. [Google Scholar] [CrossRef]
- El-Shafei, H.A.; El-Nasser, N.H.A.; Kansoh, A.L.; Ali, A.M. Biodegradation of disposable polyethylene by fungi and Streptomyces species. Polym. Degrad. Stab. 1998, 62, 361–365. [Google Scholar] [CrossRef]
- Muhonja, C.N.; Makonde, H.; Magoma, G.; Imbuga, M. Biodegradability of polyethylene by bacteria and fungi from Dandora dumpsite Nairobi-Kenya. PLoS ONE 2018, 13, e0198446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeon, H.J.; Kim, M.N. Isolation of mesophilic bacterium for biodegradation of polypropylene. Int. Biodeterior. Biodegrad. 2016, 115, 244–249. [Google Scholar] [CrossRef]
- Arkatkar, A.; Juwarkar, A.A.; Bhaduri, S.; Uppara, P.V.; Doble, M. Growth of Pseudomonas and Bacillus biofilms on pretreated polypropylene surface. Int. Biodeterior. Biodegrad. 2010, 64, 530–536. [Google Scholar] [CrossRef]
- Auta, H.S.; Emenike, C.U.; Fauziah, S.H. Screening of Bacillus strains isolated from mangrove ecosystems in Peninsular Malaysia for microplastic degradation. Environ. Pollut. 2017, 231, 1552–1559. [Google Scholar] [CrossRef] [PubMed]
- Jain, K.; Bhunia, H.; Reddy, M.S. Degradation of polypropylene–poly-L-lactide blend by bacteria isolated from compost. Bioremediation J. 2018, 22, 73–90. [Google Scholar] [CrossRef]
- Yang, Y.; Yang, J.; Wu, W.-M.; Zhao, J.; Song, Y.L.; Gao, L.C.; Yang, R.F.; Jiang, L. Biodegradation and Mineralization of Polystyrene by Plastic-Eating Mealworms: Part 2. Role of Gut Microorganisms. Environ. Sci. Technol. 2015, 49, 12087–12093. [Google Scholar] [CrossRef] [PubMed]
- Mohan, A.J.; Sekhar, V.C.; Bhaskar, T.; Nampoothiri, K.M. Microbial assisted High Impact Polystyrene (HIPS) degradation. Bioresour. Technol. 2016, 213, 204–207. [Google Scholar] [CrossRef] [PubMed]
- Shimpi, N.; Borane, M.; Mishra, S.; Kadam, M. Biodegradation of polystyrene (PS)-poly(lactic acid) (PLA) nanocomposites using Pseudomonas aeruginosa. Macromol. Res. 2012, 20, 181–187. [Google Scholar] [CrossRef]
- Sekhar, V.C.; Nampoothiri, K.M.; Mohan, A.J.; Nair, N.R.; Bhaskar, T.; Pandey, A. Microbial degradation of high impact polystyrene (HIPS), an e-plastic with decabromodiphenyl oxide and antimony trioxide. J. Hazard. Mater. 2016, 318, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Mor, R.; Sivan, A. Biofilm formation and partial biodegradation of polystyrene by the actinomycete Rhodococcus ruber. Biodegradation 2008, 19, 851–858. [Google Scholar] [CrossRef]
- Tribedi, P.; Dey, S. Pre-oxidation of low-density polyethylene (LDPE) by ultraviolet light (UV) promotes enhanced degradation of LDPE in soil. Environ. Monit. Assess. 2017, 189, 624. [Google Scholar] [CrossRef] [PubMed]
- Peng, G.Y.; Xu, P.; Zhu, B.S.; Bai, M.Y.; Li, D.J. Microplastics in freshwater river sediments in Shanghai, China: A case study of risk assessment in mega-cities. Environ. Pollut. 2018, 234, 448–456. [Google Scholar] [CrossRef] [PubMed]
- Skariyachan, S.; Setlur, A.S.; Naik, S.Y.; Naik, A.A.; Usharani, M.; Vasist, K.S. Enhanced biodegradation of low and high-density polyethylene by novel bacterial consortia formulated from plastic-contaminated cow dung under thermophilic conditions. Environ. Sci. Pollut. Res. 2017, 24, 8443–8457. [Google Scholar] [CrossRef] [PubMed]
- Skariyachan, S.; Manjunatha, V.; Sultana, S.; Jois, C.; Bai, V.; Vasist, K.S. Novel bacterial consortia isolated from plastic garbage processing areas demonstrated enhanced degradation for low density polyethylene. Environ. Sci. Pollut. Res. 2016, 23, 18307–18319. [Google Scholar] [CrossRef]
- Syranidou, E.; Karkanorachaki, K.; Amorotti, F.; Repouskou, E.; Kroll, K.; Kolvenbach, B.; Corvini, P.F.X.; Fava, F.; Kalogerakis, N. Development of tailored indigenous marine consortia for the degradation of naturally weathered polyethylene films. PLoS ONE 2017, 12, e0183984. [Google Scholar] [CrossRef] [Green Version]
- Aravinthan, A.; Arkatkar, A.; Juwarkar, A.A.; Doble, M. Synergistic growth of Bacillus and Pseudomonas and its degradation potential on pretreated polypropylene. Prep. Biochem. Biotechnol. 2016, 46, 109–115. [Google Scholar] [CrossRef]
- Arkatkar, A.; Arutchelvi, J.; Bhaduri, S.; Uppara, P.V.; Doble, M. Degradation of unpretreated and thermally pretreated polypropylene by soil consortia. Int. Biodeterior. Biodegrad. 2009, 63, 106–111. [Google Scholar] [CrossRef]
- Cacciari, I.; Quatrini, P.; Zirletta, G.; Mincione, E.; Vinciguerra, V.; Lupattelli, P.; Sermanni, G.G. Isotactic polypropylene biodegradation by a microbial community: Physicochemical characterization of metabolites produced. Appl. Environ. Microbiol. 1993, 59, 3695–3700. [Google Scholar] [CrossRef] [Green Version]
- Santacruz-Juárez, E.; Buendia-Corona, R.E.; Ramírez, R.E.; Sánchez, C. Fungal enzymes for the degradation of polyethylene: Molecular docking simulation and biodegradation pathway proposal. J. Hazard. Mater. 2021, 411, 125118. [Google Scholar] [CrossRef] [PubMed]
- Jolivalt, C.; Madzak, C.; Brault, A.; Caminade, E.; Malosse, C.; Mougin, C. Expression of laccase IIIb from the white-rot fungus Trametes versicolor in the yeast Yarrowia lipolytica for environmental applications. Appl. Microbiol. Biotechnol. 2005, 66, 450–456. [Google Scholar] [CrossRef] [PubMed]
- Crestini, C.; Jurasek, L.; Argyropoulos, D.S. On the Mechanism of the Laccase–Mediator System in the Oxidation of Lignin. Chem.—A Eur. J. 2003, 9, 5371–5378. [Google Scholar] [CrossRef] [PubMed]
- Fujisawa, M.; Hirai, H.; Nishida, T. Degradation of Polyethylene and Nylon-66 by the Laccase-Mediator System. J. Polym. Environ. 2001, 9, 103–108. [Google Scholar] [CrossRef]
- Johnnie, D.A.; Issac, R.; Prabha, M.L. Bio Efficacy Assay of Laccase Isolated and Characterized from Trichoderma viride in Biodegradation of Low Density Polyethylene (LDPE) and Textile Industrial Effluent Dyes. J. Pure Appl. Microbiol. 2021, 15, 410–420. [Google Scholar] [CrossRef]
- Yeom, S.-J.; Le, T.-K.; Yun, C.-H. P450-driven plastic-degrading synthetic bacteria. Trends Biotechnol. 2022, 40, 166–179. [Google Scholar] [CrossRef]
- Chen, C.-C.; Dai, L.H.; Ma, L.X.; Guo, R.-T. Enzymatic degradation of plant biomass and synthetic polymers. Nat. Rev. Chem. 2020, 4, 114–126. [Google Scholar] [CrossRef]
- Ihssen, J.; Reiss, R.; Luchsinger, R.; Thöny-Meyer, L.; Richter, M. Biochemical properties and yields of diverse bacterial laccase-like multicopper oxidases expressed in Escherichia coli. Sci. Rep. 2015, 5, 10465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mo, Y.; Lao, H.I.; Au, S.W.; Li, I.C.; Hu, J.; Yuen, H.M.; Cheong, W.M.; Lo, O.L.I.; Seak, L.C.U. Expression, secretion and functional characterization of three laccases in E. coli. Synth. Syst. Biotechnol. 2022, 7, 474–480. [Google Scholar] [CrossRef] [PubMed]
- Jeon, H.J.; Kim, M.N. Functional analysis of alkane hydroxylase system derived from Pseudomonas aeruginosa E7 for low molecular weight polyethylene biodegradation. Int. Biodeterior. Biodegradation 2015, 103, 141–146. [Google Scholar] [CrossRef]
- Gao, R.; Liu, R.; Sun, C. A marine fungus Alternaria alternata FB1 efficiently degrades polyethylene. J. Hazard. Mater. 2022, 431, 128671. [Google Scholar] [CrossRef]
- Wang, T.-N.; Zhao, M. A simple strategy for extracellular production of CotA laccase in Escherichia coli and decolorization of simulated textile effluent by recombinant laccase. Appl. Microbiol. Biotechnol. 2016, 101, 685–696. [Google Scholar] [CrossRef] [PubMed]
- Grandes-Blanco, A.I.; Tlecuitl-Beristain, S.; Díaz, R.; Sánchez, C.; Téllez-Téllez, M.; Márquez-Domínguez, L.; Santos-López, G.; Díaz-Godínez, G. Heterologous Expression of Laccase (LACP83) of Pleurotus ostreatus. BioResources 2017, 12, 3211–3221. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Zheng, B.; Jiang, D.; Qin, W.S. Overexpression of a Laccase with Dye Decolorization Activity from Bacillus sp. Induced in Escherichia coli. J. Mol. Microbiol. Biotechnol. 2017, 27, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Sowmya, H.V.; Ramalingappa; Krishnappa, M.; Thippeswamy, B. Degradation of polyethylene by Trichoderma harzianum—SEM, FTIR, and NMR analyses. Environ. Monit. Assess. 2014, 186, 6577–6586. [Google Scholar] [CrossRef] [PubMed]
- Sowmya, H.V.; Ramalingappa; Krishnappa, M.; Thippeswamy, B. Degradation of polyethylene by Penicillium simplicissimum isolated from local dumpsite of Shivamogga district. Environ. Dev. Sustain. 2015, 17, 731–745. [Google Scholar] [CrossRef]
- De Pourcq, K.; Vervecken, W.; Dewerte, I.; Valevska, A.; Van Hecke, A.; Callewaert, N. Engineering the yeast Yarrowia lipolytica for the production of therapeutic proteins homogeneously glycosylated with Man8GlcNAc2 and Man5GlcNAc2. Microb. Cell Factories 2012, 11, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogrydziak, D.M.; Nicaud, J.-M. Characterization of Yarrowia lipolyticaXPR2 multi-copy strains over-producing alkaline extracellular protease—A system for rapidly increasing secretory pathway cargo loads. FEMS Yeast Res. 2012, 12, 938–948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madzak, C.; Tréton, B.; Blanchin-Roland, S. Strong hybrid promoters and integrative expression/secretion vectors for quasi-constitutive expression of heterologous proteins in the yeast Yarrowia lipolytica. J. Mol. Microbiol. Biotechnol. 2000, 2, 207–216. [Google Scholar] [PubMed]
- Madzak, C.; Beckerich, J.-M. Heterologous Protein Expression and Secretion in Yarrowia lipolytica. In Yarrowia lipolytica: Biotechnological Applications; Springer: Berlin/Heidelberg, Germany, 2013; Volume 25, pp. 1–76. [Google Scholar] [CrossRef]
- Jenkins, S.; Quer, A.M.; Fonseca, C.; Varrone, C. Microbial Degradation of Plastic: New Plastic Degraders, Mixed Cultures and Engineering Strategies. Soil Microenvironment for Bioremediation and Polymer Production; Wiley: Hoboken, NJ, USA, 2019; pp. 215–238. [Google Scholar] [CrossRef]
- Li, Z.H.; Wang, X.N.; Zhang, H.R. Balancing the non-linear rosmarinic acid biosynthetic pathway by modular co-culture engineering. Metab. Eng. 2019, 54, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Shahab, R.L.; Brethauer, S.; Davey, M.P.; Smith, A.G.; Vignolini, S.; Luterbacher, J.S.; Studer, M.H. A heterogeneous microbial consortium producing short-chain fatty acids from lignocellulose. Science 2020, 369, 1073. [Google Scholar] [CrossRef] [PubMed]
- Zhi, Y.; Wu, Q.; Xu, Y. Production of surfactin from waste distillers’ grains by co-culture fermentation of two Bacillus amyloliquefaciens strains. Bioresour. Technol. 2017, 235, 96–103. [Google Scholar] [CrossRef]
- Chen, W.; Kong, Y.C.; Li, J.D.; Sun, Y.Y.; Min, J.; Hu, X.K. Enhanced biodegradation of crude oil by constructed bacterial consortium comprising salt-tolerant petroleum degraders and biosurfactant producers. Int. Biodeterior. Biodegrad. 2020, 154, 105047. [Google Scholar] [CrossRef]
- Qi, X.; Ma, Y.; Chang, H.; Li, B.; Ding, M.; Yuan, Y. Evaluation of PET Degradation Using Artificial Microbial Consortia. Front. Microbiol. 2021, 12, 778828. [Google Scholar] [CrossRef]
- Ma, Y.; Yao, M.D.; Li, B.Z.; Ding, M.Z.; He, B.; Chen, S.; Zhou, X.; Yuan, Y.J. Enhanced Poly(ethylene terephthalate) Hydrolase Activity by Protein Engineering. Engineering 2018, 4, 888–893. [Google Scholar] [CrossRef]
- Huang, Q.-S.; Yan, Z.-F.; Chen, X.-Q.; Du, Y.-Y.; Li, J.; Liu, Z.-Z.; Xia, W.; Chen, S.; Wu, J. Accelerated biodegradation of polyethylene terephthalate by Thermobifida fusca cutinase mediated by Stenotrophomonas pavanii. Sci. Total Environ. 2022, 808, 152107. [Google Scholar] [CrossRef] [PubMed]
- Skariyachan, S.; Taskeen, N.; Kishore, A.P.; Krishna, B.V.; Naidu, G. Novel consortia of Enterobacter and Pseudomonas formulated from cow dung exhibited enhanced biodegradation of polyethylene and polypropylene. J. Environ. Manag. 2021, 284, 112030. [Google Scholar] [CrossRef] [PubMed]
- Wondraczek, L.; Pohnert, G.; Schacher, F.H.; Köhler, A.; Gottschaldt, M.; Schubert, U.S.; Küsel, K.; Brakhage, A.A. Artificial Microbial Arenas: Materials for Observing and Manipulating Microbial Consortia. Adv. Mater. 2019, 31, e1900284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Bogaert, I.N.A.; Demey, M.; Develter, D.; Soetaert, W.; Vandamme, E.J. Importance of the cytochrome P450 monooxygenase CYP52 family for the sophorolipid-producing yeast Candida bombicola. FEMS Yeast Res. 2009, 9, 87–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zinjarde, S.; Apte, M.; Mohite, P.; Kumar, A.R. Yarrowia lipolytica and pollutants: Interactions and applications. Biotechnol. Adv. 2014, 32, 920–933. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Ma, C.; Wang, D.; Liu, H.; Zhu, C.C.; Xu, H.N. Nutrient drip irrigation for refractory hydrocarbon removal and microbial community shift in a historically petroleum-contaminated soil. Sci. Total Environ. 2020, 713, 136331. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Wang, J.; Gao, C.M.; Zhang, Y.M.; Du, W. A novel exopolysaccharide-producing and long-chain n-alkane degrading bacterium Bacillus licheniformis strain DM-1 with potential application for in-situ enhanced oil recovery. Sci. Rep. 2020, 10, 8519. [Google Scholar] [CrossRef] [PubMed]
- Iwama, R.; Hara, M.; Mizuike, A.; Horiuchi, H.; Fukuda, R. Osh6p, a homologue of the oxysterol-binding protein, is involved in production of functional cytochrome P450 belonging to CYP52 family in n-alkane-assimilating yeast Yarrowia lipolytica. Biochem. Biophys. Res. Commun. 2018, 499, 836–842. [Google Scholar] [CrossRef]
- Ensign, S.A.; Allen, J.R. Aliphatic Epoxide Carboxylation. Annu. Rev. Biochem. 2003, 72, 55–76. [Google Scholar] [CrossRef]
- Choi, K.R.; Jang, W.D.; Yang, D.; Cho, J.; Park, D.; Lee, S.Y. Systems metabolic engineering strategies: Integrating systems and synthetic biology with metabolic engineering. Trends Biotechnol. 2019, 37, 817–837. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Liu, Z.; Jiang, H.; Mao, X.Z. Biotechnology advances in β-carotene production by microorganisms. Trends Food Sci. Technol. 2021, 111, 322–332. [Google Scholar] [CrossRef]
- Zheng, Y.; Chen, J.-C.; Ma, Y.-M.; Chen, G.-Q. Engineering biosynthesis of polyhydroxyalkanoates (PHA) for diversity and cost reduction. Metab. Eng. 2020, 58, 82–93. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.Y.; Rhie, M.N.; Kim, H.T.; Joo, J.C.; Cho, I.J.; Son, J.; Jo, S.Y.; Sohn, Y.J.; Baritugo, K.-A.; Pyo, J.; et al. Metabolic engineering for the synthesis of polyesters: A 100-year journey from polyhydroxyalkanoates to non-natural microbial polyesters. Metab. Eng. 2020, 58, 47–81. [Google Scholar] [CrossRef]
- Zhao, J.T.; Li, F.; Cao, Y.X.; Zhang, X.B.; Chen, T.; Song, H.; Wang, Z.W. Microbial extracellular electron transfer and strategies for engineering electroactive microorganisms. Biotechnol. Adv. 2021, 53, 107682. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Structures of major synthetic polymers.

Figure 2.
The process of PE biodegradation.

Figure 3.
Artificial microbial consortia for complete PE biodegradation.

Table 2.
Microbial consortia capable of polyolefin degradation.
| Polyolefin | Microorganisms | Microorganism Source | Pretreatment | Experimental Condition | Biodegradation Result | Reference |
|---|---|---|---|---|---|---|
| PE | Mixed microorganisms | Microbial activated soil | Thermally pretreated | Incubation for 180 days at 60 °C | Mineralization percentage: 60.0% | [26] |
| Lysinibacillus xylanilyticus and Aspergillus niger | Landfill soils | UV-irradiated and non-UV-irradiated LDPE films | Incubation for 126 days in soil | Mineralization percentage: 29.5% (UV-irradiated) | [29] | |
| Soil microorgannisms | Soil | UV-irradiated and non-UV-irradiated | Incubation for 28 days | Weight loss: 6.0% and 3.5% | [63] | |
| Comamonas, Delftia, and Stenotrophomonas | Degraded plastic debris | Unpretreated LDPE films | Shaken flasks incubated for 90 days at 28 °C | Changes in chemical properties | [64] | |
| Brevibacillus sp. and Aneurinibacillus sp. | Waste management landfills and sewage treatment plants | Unpretreated HDPE, LDPE films and pellets | Incubation for 140 days at 50 °C | Weight loss for LDPE and HDPE strips was 58.2% and 46.6% respectively; weight loss for LDPE and HDPE pellets was 45.7 % and 37.2%, respectively | [47] | |
| Bacillus sp. and Paenibacillus sp. | Landfill site | Unpretreated PE microplastic granules | Incubation for 60 days at 30 °C | Weight loss: 16.7%; mean diameter reduction: 22.8% | [46] | |
| Artificial thermophilic bacterial consortium composed of bacterial isolates (Bacillus vallismortis, Pseudomonas protegens, Stenotrophomonas sp. and Paenibacillus sp.) | Dung of cows fed off plastic-contaminated pastures | LDPE and HDPE films and pellets | Incubation for 120 days at 55 °C | Gravimetric weight loss percentages of 75.0%, 55.0%, 60.0% and 43.0% for LDPE film, pellets, HDPE film and pellets, respectively | [65] | |
| Two Enterobacter sp. and one Pantoea sp. | Plastic garbage processing areas | LDPE films and pellets | Incubation for 120 days at 37 °C | Maximum weight loss: 81.0% | [66] | |
| Lysinibacillus. sp. and Salinibacterium sp. | Plastic samples and surface water | LDPE and HDPE pieces | Incubation at 25 °C for 6 months | Weight loss: 15.0% for LDPE after 4 months and 5.5% for HDPE after 6 months | [67] | |
| pp | Bacillus and Pseudomonas | UV- or thermally pretreated PP films | Flasks incubated at 28 ± 2 °C and 180 rpm for 12 months | Weight loss: 1.9% | [68] | |
| Microbial consortium | Plastic dumping site | Thermally pretreated PP films | Incubation for 1 year | Weight loss: 10.7% | [69] | |
| Mixed soil community | Soil samples rich in plastic waste | Isotactic PP films | Incubation for 5 months | The film had 40% methylene chloride extractable compounds, and a mixture of hydrocarbons (between C10H22 and C31H64) was detected and identified in the extract | [70] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhang, N.; Ding, M.; Yuan, Y. Current Advances in Biodegradation of Polyolefins. Microorganisms 2022, 10, 1537. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10081537
AMA Style
Zhang N, Ding M, Yuan Y. Current Advances in Biodegradation of Polyolefins. Microorganisms. 2022; 10(8):1537. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10081537
Chicago/Turabian StyleZhang, Ni, Mingzhu Ding, and Yingjin Yuan. 2022. "Current Advances in Biodegradation of Polyolefins" Microorganisms 10, no. 8: 1537. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10081537
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.