
Toxoplasma gondii Serotypes in Italian and Foreign Populations: A Cross-Sectional Study Using a Homemade ELISA Test

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design and Setting
2.2. Study Population and Data
- Sex: female or male;
- Nationality: Italian (patients born in Italy) or foreign (patients born in other countries);
- Place of birth: Italy, Africa, South America, Asia, or Europe (except Italy).
2.3. Identification of T. gondii Antibodies in Sera Samples
2.4. ELISA Protocol for the T. gondii Serotype Determination
2.5. Statistical Methods
3. Results
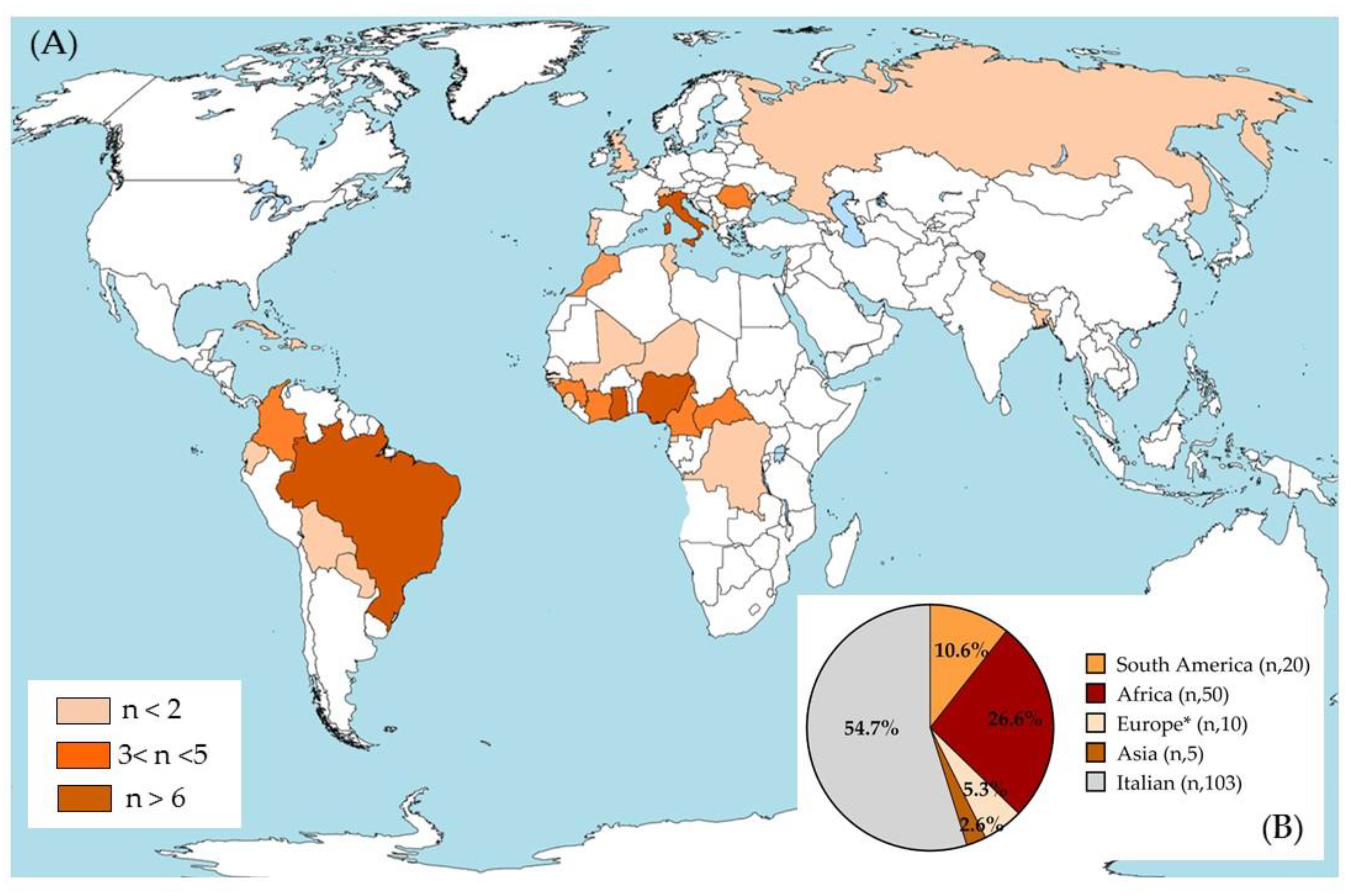
3.1. Demographic Characteristics of the Overall Study Population Included
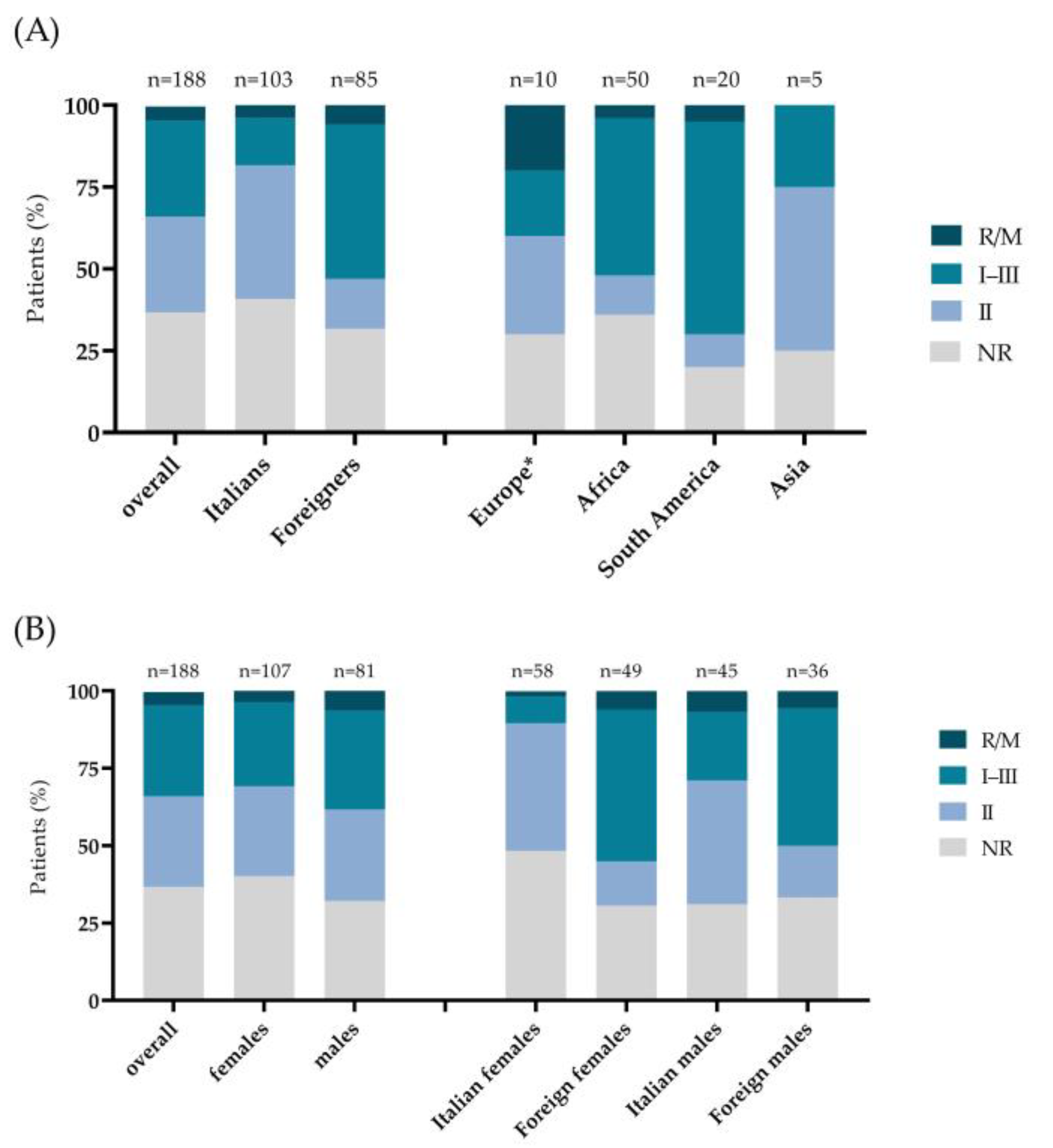
3.2. Distribution of T. gondii Serotypes in the Overall Sample and in the Italian, Foreigner, Male, and Female Subgroups
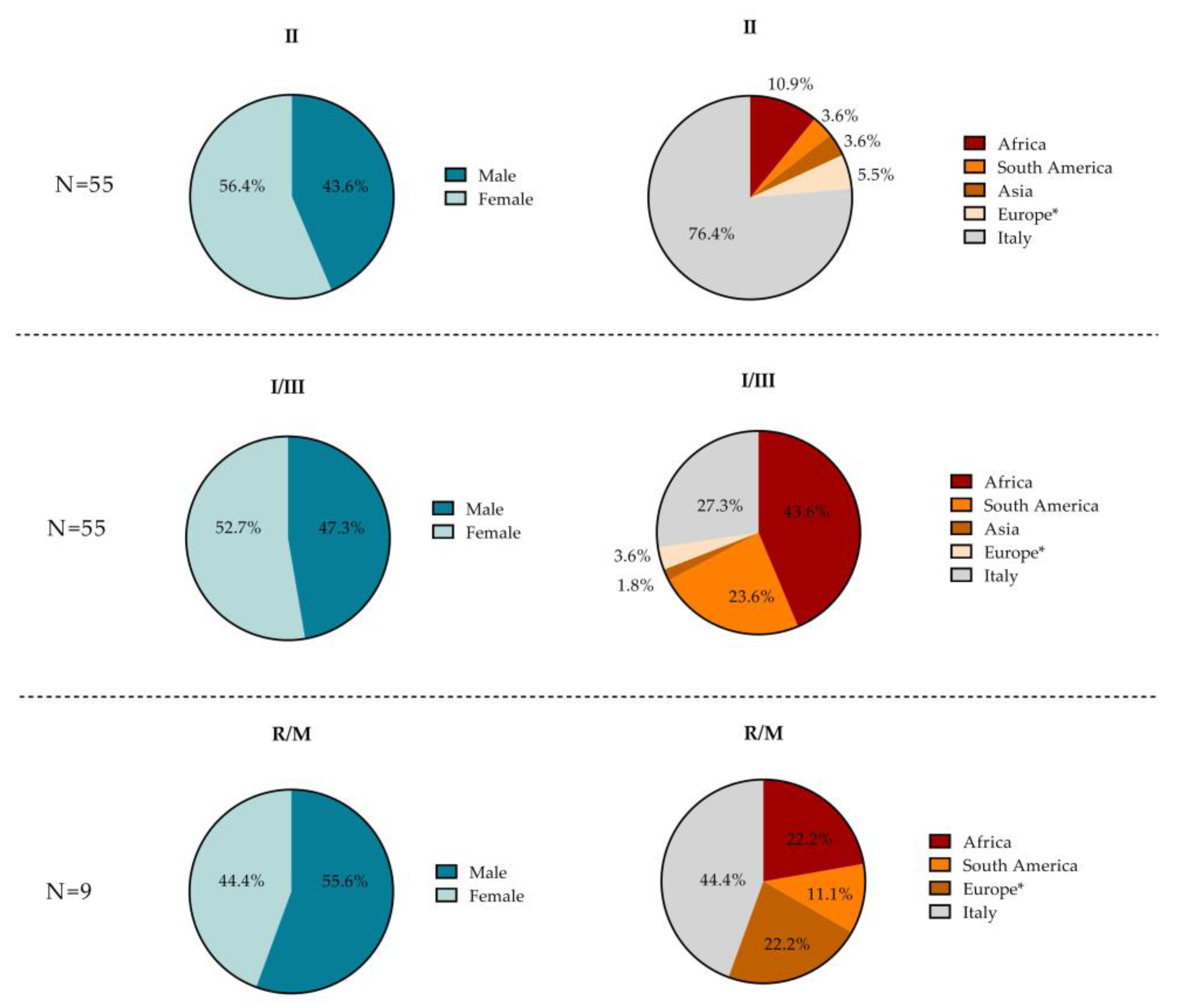
3.3. Demographic Characteristics According to T. gondii Serotype
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Robert-Gangneux, F.; Dardé, M.-L. Epidemiology of and Diagnostic Strategies for Toxoplasmosis. Clin. Microbiol. Rev. 2012, 25, 264–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shwab, E.K.; Zhu, X.-Q.; Majumdar, D.; Pena, H.F.J.; Gennari, S.M.; Dubey, J.P.; Su, C. Geographical Patterns of Toxoplasma Gondii Genetic Diversity Revealed by Multilocus PCR-RFLP Genotyping. Parasitology 2014, 141, 453–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorenzi, H.; Khan, A.; Behnke, M.S.; Namasivayam, S.; Swapna, L.S.; Hadjithomas, M.; Karamycheva, S.; Pinney, D.; Brunk, B.P.; Ajioka, J.W.; et al. Local Admixture of Amplified and Diversified Secreted Pathogenesis Determinants Shapes Mosaic Toxoplasma Gondii Genomes. Nat. Commun. 2016, 7, 10147. [Google Scholar] [CrossRef] [PubMed]
- Galal, L.; Hamidović, A.; Dardé, M.L.; Mercier, M. Diversity of Toxoplasma Gondii Strains at the Global Level and Its Determinants. Food Waterborne Parasitol. 2019, 15, e00052. [Google Scholar] [CrossRef] [PubMed]
- Mercier, A.; Devillard, S.; Ngoubangoye, B.; Bonnabau, H.; Bañuls, A.-L.; Durand, P.; Salle, B.; Ajzenberg, D.; Dardé, M.-L. Additional Haplogroups of Toxoplasma Gondii out of Africa: Population Structure and Mouse-Virulence of Strains from Gabon. PLoS Negl. Trop. Dis. 2010, 4, e876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shwab, E.K.; Saraf, P.; Zhu, X.-Q.; Zhou, D.-H.; McFerrin, B.M.; Ajzenberg, D.; Schares, G.; Hammond-Aryee, K.; van Helden, P.; Higgins, S.A.; et al. Human Impact on the Diversity and Virulence of the Ubiquitous Zoonotic Parasite. Proc. Natl. Acad. Sci. USA 2018, 115, E6956–E6963. [Google Scholar] [CrossRef] [Green Version]
- Lehmann, T.; Marcet, P.L.; Graham, D.H.; Dahl, E.R.; Dubey, J.P. Globalization and the Population Structure of Toxoplasma Gondii. Proc. Natl. Acad. Sci. USA 2006, 103, 11423–11428. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Escobar, M.; Schares, G.; Maksimov, P.; Joeres, M.; Ortega-Mora, L.M.; Calero-Bernal, R. Toxoplasma Gondii Genotyping: A Closer Look into Europe. Front. Cell. Infect. Microbiol. 2022, 12, 842595. [Google Scholar] [CrossRef]
- Bertranpetit, E.; Jombart, T.; Paradis, E.; Pena, H.; Dubey, J.; Su, C.; Mercier, A.; Devillard, S.; Ajzenberg, D. Phylogeography of Toxoplasma Gondii Points to a South American Origin. Infect. Genet. Evol. 2017, 48, 150–155. [Google Scholar] [CrossRef]
- Su, C.; Khan, A.; Zhou, P.; Majumdar, D.; Ajzenberg, D.; Dardé, M.-L.; Zhu, X.-Q.; Ajioka, J.W.; Rosenthal, B.M.; Dubey, J.P.; et al. Globally Diverse Toxoplasma Gondii Isolates Comprise Six Major Clades Originating from a Small Number of Distinct Ancestral Lineages. Proc. Natl. Acad. Sci. USA 2012, 109, 5844–5849. [Google Scholar] [CrossRef] [Green Version]
- Galal, L.; Ajzenberg, D.; Hamidović, A.; Durieux, M.-F.; Dardé, M.-L.; Mercier, A. Toxoplasma and Africa: One Parasite, Two Opposite Population Structures. Trends Parasitol. 2018, 34, 140–154. [Google Scholar] [CrossRef] [PubMed]
- Chaichan, P.; Mercier, A.; Galal, L.; Mahittikorn, A.; Ariey, F.; Morand, S.; Boumédiène, F.; Udonsom, R.; Hamidovic, A.; Murat, J.B.; et al. Geographical Distribution of Toxoplasma Gondii Genotypes in Asia: A Link with Neighboring Continents. Infect. Genet. Evol. 2017, 53, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Barragan, A.; Sibley, L.D. Transepithelial Migration of Toxoplasma Gondii Is Linked to Parasite Motility and Virulence. J. Exp. Med. 2002, 195, 1625–1633. [Google Scholar] [CrossRef] [PubMed]
- Barragan, A.; David Sibley, L. Migration of Toxoplasma Gondii across Biological Barriers. Trends Microbiol. 2003, 11, 426–430. [Google Scholar] [CrossRef]
- Sibley, L.D.; Boothroyd, J.C. Virulent Strains of Toxoplasma Gondii Comprise a Single Clonal Lineage. Nature 1992, 359, 82–85. [Google Scholar] [CrossRef]
- Lambert, H.; Barragan, A. Modelling Parasite Dissemination: Host Cell Subversion and Immune Evasion by Toxoplasma Gondii. Cell. Microbiol. 2010, 12, 292–300. [Google Scholar] [CrossRef]
- Carme, B.; Bissuel, F.; Ajzenberg, D.; Bouyne, R.; Aznar, C.; Demar, M.; Bichat, S.; Louvel, D.; Bourbigot, A.M.; Peneau, C.; et al. Severe Acquired Toxoplasmosis in Immunocompetent Adult Patients in French Guiana. J. Clin. Microbiol. 2002, 40, 4037–4044. [Google Scholar] [CrossRef] [Green Version]
- De Salvador-Guillouet, F.; Ajzenberg, D.; Chaillou-Opitz, S.; Saint-Paul, M.C.; Dunais, B.; Dellamonica, P.; Marty, P. Severe Pneumonia during Primary Infection with an Atypical Strain of Toxoplasma Gondii in an Immunocompetent Young Man. J. Infect. 2006, 53, e47–e50. [Google Scholar] [CrossRef]
- Carme, B.; Demar, M.; Ajzenberg, D.; Dardé, M.L. Severe Acquired Toxoplasmosis Caused by Wild Cycle of Toxoplasma gondii, French Guiana. Emerg. Infect. Dis. 2009, 15, 656–658. [Google Scholar] [CrossRef]
- Demar, M.; Hommel, D.; Djossou, F.; Peneau, C.; Boukhari, R.; Louvel, D.; Bourbigot, A.-M.; Nasser, V.; Ajzenberg, D.; Darde, M.-L.; et al. Acute Toxoplasmoses in Immunocompetent Patients Hospitalized in an Intensive Care Unit in French Guiana. Clin. Microbiol. Infect. 2012, 18, E221–E231. [Google Scholar] [CrossRef] [Green Version]
- de-la-Torre, A.; González, G.; Díaz-Ramirez, J.; Gómez-Marín, J.E. Screening by Ophthalmoscopy for Toxoplasma Retinochoroiditis in Colombia. Am. J. Ophthalmol. 2007, 143, 354–356. [Google Scholar] [CrossRef] [PubMed]
- Khan, A. Genetic Divergence of Toxoplasma Gondii Strains Associated with Ocular Toxoplasmosis, Brazil. Emerg. Infect. Dis. 2006, 12, 942–949. [Google Scholar] [CrossRef] [PubMed]
- Grigg, M.E.; Boothroyd, J.C. Rapid Identification of Virulent Type I Strains of the Protozoan Pathogen Toxoplasma Gondii by PCR-Restriction Fragment Length Polymorphism Analysis at the B1 Gene. J. Clin. Microbiol. 2001, 39, 398–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ajzenberg, D.; Bañuls, A.-L.; Tibayrenc, M.; Dardé, M.L. Microsatellite Analysis of Toxoplasma Gondii Shows Considerable Polymorphism Structured into Two Main Clonal Groups. Int. J. Parasitol. 2002, 32, 27–38. [Google Scholar] [CrossRef]
- Sousa, S.; Ajzenberg, D.; Vilanova, M.; Costa, J.; Dardé, M.-L. Use of GRA6-Derived Synthetic Polymorphic Peptides in an Immunoenzymatic Assay to Serotype Toxoplasma Gondii in Human Serum Samples Collected from Three Continents. Clin. Vaccine Immunol. 2008, 15, 1380–1386. [Google Scholar] [CrossRef] [Green Version]
- Parmley, S.F.; Gross, U.; Sucharczuk, A.; Windeck, T.; Sgarlato, G.D.; Remington, J.S. Two Alleles of the Gene Encoding Surface Antigen P22 in 25 Strains of Toxoplasma Gondii. J. Parasitol. 1994, 80, 293–301. [Google Scholar] [CrossRef]
- Kong, J.-T.; Grigg, M.E.; Uyetake, L.; Parmley, S.; Boothroyd, J.C. Serotyping of Toxoplasma Gondii Infections in Humans Using Synthetic Peptides. J. Infect. Dis. 2003, 187, 1484–1495. [Google Scholar] [CrossRef] [Green Version]
- Arranz-Solís, D.; Cordeiro, C.; Young, L.H.; Dardé, M.L.; Commodaro, A.G.; Grigg, M.E.; Saeij, J.P.J. Serotyping of Toxoplasma Gondii Infection Using Peptide Membrane Arrays. Front. Cell. Infect. Microbiol. 2019, 9, 408. [Google Scholar] [CrossRef]
- Piffer, S.; Lauriola, A.L.; Pradal, U.; Collini, L.; Dell’Anna, L.; Pavanello, L. Toxoplasma Gondii Infection during Pregnancy: A Ten-Year Observation in the Province of Trento, Italy. Le Infez. Med. 2020, 28, 603–610. [Google Scholar]
- Tomasoni, L.R.; Sosta, E.; Beltrame, A.; Rorato, G.; Bigoni, S.; Frusca, T.; Zanardini, C.; Driul, L.; Magrini, F.; Viale, P.; et al. Antenatal Screening for Mother to Child Infections in Immigrants and Residents: The Case of Toxoplasmosis in Northern Italy. J. Immigr. Minority Health 2010, 12, 834–840. [Google Scholar] [CrossRef]
- Fanigliulo, D.; Marchi, S.; Montomoli, E.; Trombetta, C.M. Toxoplasma Gondii in Women of Childbearing Age and during Pregnancy: Seroprevalence Study in Central and Southern Italy from 2013 to 2017. Parasite 2020, 27, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Battisti, E.; Zanet, S.; Trisciuoglio, A.; Bruno, S.; Ferroglio, E. Circulating Genotypes of Toxoplasma Gondii in Northwestern Italy. Vet. Parasitol. 2018, 253, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Ferroglio, E.; Bosio, F.; Trisciuoglio, A.; Zanet, S. Toxoplasma Gondii in Sympatric Wild Herbivores and Carnivores: Epidemiology of Infection in the Western Alps. Parasites Vectors 2014, 7, 196. [Google Scholar] [CrossRef] [Green Version]
- Formenti, N.; Gaffuri, A.; Trogu, T.; Viganò, R.; Ferrari, N.; Lanfranchi, P. Spread and Genotype of Toxoplasma Gondii in Naturally Infected Alpine Chamois (Rupicapra r. Rupicapra). Parasitol. Res. 2016, 115, 2115–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sgroi, G.; Viscardi, M.; Santoro, M.; Borriello, G.; D’Alessio, N.; Boccia, F.; Pacifico, L.; Fioretti, A.; Veneziano, V.; Fusco, G. Genotyping of Toxoplasma Gondii in Wild Boar (Sus Scrofa) in Southern Italy: Epidemiological Survey and Associated Risk for Consumers. Zoonoses Public Health 2020, 67, 805–813. [Google Scholar] [CrossRef] [PubMed]
- Pagliuca, C.; Pastore, G.; Scaglione, E.; Migliucci, A.; Maruotti, G.M.; Cicatiello, A.G.; Salvatore, E.; Picardi, M.; Camilla Sammartino, J.; Consiglio Buonocore, M.; et al. Genotyping of Toxoplasma Gondii Strain Directly from Human CSF Samples of Congenital Toxoplasmosis Clinical Case. New Microbiol 2017, 40, 151–154. [Google Scholar]
- Marcer, F.; Marchiori, E.; Centelleghe, C.; Ajzenberg, D.; Gustinelli, A.; Meroni, V.; Mazzariol, S. Parasitological and Pathological Findings in Fin Whales Balaenoptera Physalus Stranded along Italian Coastlines. Dis. Aquat. Org. 2019, 133, 25–37. [Google Scholar] [CrossRef]
- Migliore, S.; La Marca, S.; Stabile, C.; Di Marco Lo Presti, V.; Vitale, M. A Rare Case of Acute Toxoplasmosis in a Stray Dog Due to Infection of T. Gondii Clonal Type I: Public Health Concern in Urban Settings with Stray Animals? BMC Vet. Res. 2017, 13, 249. [Google Scholar] [CrossRef]
- Sousa, S.; Puime, A.; da Costa, J.M.C.; Marie-Laure, D. Serotyping Toxoplasma Gondii: A Cross-Sectional Study in Uruguay. J. Infect. Dis. Epidemiol 2017, 3, 039. [Google Scholar] [CrossRef] [Green Version]
- Jokelainen, P.; Isomursu, M.; Näreaho, A.; Oksanen, A. Natural Toxoplasma Gondii Infections In European Brown Hares And Mountain Hares In Finland: Proportional Mortality Rate, Antibody Prevalence, And Genetic Characterization. J. Wildl. Dis. 2011, 47, 154–163. [Google Scholar] [CrossRef]
- Verma, S.K.; Ajzenberg, D.; Rivera-Sanchez, A.; Su, C.; Dubey, J.P. Genetic Characterization of Toxoplasma Gondii Isolates from Portugal, Austria and Israel Reveals Higher Genetic Variability within the Type II Lineage. Parasitology 2015, 142, 948–957. [Google Scholar] [CrossRef] [PubMed]
- Waap, H.; Vilares, A.; Rebelo, E.; Gomes, S.; Ângelo, H. Epidemiological and Genetic Characterization of Toxoplasma Gondii in Urban Pigeons from the Area of Lisbon (Portugal). Vet. Parasitol. 2008, 157, 306–309. [Google Scholar] [CrossRef]
- Pomares, C.; Ajzenberg, D.; Bornard, L.; Bernardin, G.; Hasseine, L.; Darde, M.-L.; Marty, P. Toxoplasmosis and Horse Meat, France. Emerg. Infect. Dis. 2011, 17, 1327–1328. [Google Scholar] [CrossRef]
- Leroy, J.; Houzé, S.; Dardé, M.-L.; Yéra, H.; Rossi, B.; Delhaes, L.; Gabriel, F.; Loubet, P.; Deleplancque, A.-S.; Senneville, E.; et al. Severe Toxoplasmosis Imported from Tropical Africa in Immunocompetent Patients: A Case Series. Travel Med. Infect. Dis. 2020, 35, 101509. [Google Scholar] [CrossRef]
- Hamilton, C.M.; Black, L.; Oliveira, S.; Burrells, A.; Bartley, P.M.; Melo, R.P.B.; Chianini, F.; Palarea-Albaladejo, J.; Innes, E.A.; Kelly, P.J.; et al. Comparative Virulence of Caribbean, Brazilian and European Isolates of Toxoplasma Gondii. Parasites Vectors 2019, 12, 104. [Google Scholar] [CrossRef] [PubMed]
- Howe, D.K.; Sibley, L.D. Toxoplasma Gondii Comprises Three Clonal Lineages: Correlation of Parasite Genotype with Human Disease. J. Infect. Dis. 1995, 172, 1561–1566. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, S.A.; Amouei, A.; Sharif, M.; Sarvi, S.; Galal, L.; Javidnia, J.; Pagheh, A.S.; Gholami, S.; Mizani, A.; Daryani, A. Human Toxoplasmosis: A Systematic Review for Genetic Diversity of Toxoplasma Gondii in Clinical Samples. Epidemiol. Infect. 2019, 147, e36. [Google Scholar] [CrossRef] [Green Version]
- Velmurugan, G.V.; Dubey, J.P.; Su, C. Genotyping Studies of Toxoplasma Gondii Isolates from Africa Revealed That the Archetypal Clonal Lineages Predominate as in North America and Europe. Vet. Parasitol. 2008, 155, 314–318. [Google Scholar] [CrossRef]
- Hamidović, A.; Etougbétché, J.R.; Tonouhewa, A.B.N.; Galal, L.; Dobigny, G.; Houémènou, G.; Da Zoclanclounon, H.; Amagbégnon, R.; Laleye, A.; Fievet, N.; et al. A Hotspot of Toxoplasma Gondii Africa 1 Lineage in Benin: How New Genotypes from West Africa Contribute to Understand the Parasite Genetic Diversity Worldwide. PLoS Negl. Trop. Dis. 2021, 15, e0008980. [Google Scholar] [CrossRef]
- Nissapatorn, V.; Lau, Y.-L.; Fong, M.-Y. Toxoplasma Gondii: The Parasite in Trend. In Parasites and Their Vectors; Lim, Y.A.L., Vythilingam, I., Eds.; Springer: Vienna, Austria, 2013; pp. 155–175. ISBN 978-3-7091-1552-7. [Google Scholar]
- Lass, A.; Ma, L.; Kontogeorgos, I.; Zhang, X.; Li, X.; Karanis, P. First Molecular Detection of Toxoplasma Gondii in Vegetable Samples in China Using Qualitative, Quantitative Real-Time PCR and Multilocus Genotyping. Sci. Rep. 2019, 9, 17581. [Google Scholar] [CrossRef]
- Armand, B.; Solhjoo, K.; Kordshooli, M.S.; Davami, M.H.; Pourahmad, M.; Orfaee, V. Toxoplasma Gondii Type I, Predominant Genotype Isolated from Sheep in South of Iran. Vet. World 2017, 10, 386–392. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| T. gondii Serotypes | Females N = 107 n/N (%) | Males N = 81 n/N (%) | p § | Italians N = 103 n/N (%) | Foreigners N = 85 n/N (%) | p § | Italian Females N = 58 n/N (%) | Foreign Females N = 49 n/N (%) | p § | Italian Males N = 45 n/N (%) | Foreign Males N = 36 n/N (%) | p § |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| I–III, N = 55 | 29/107 (27.1) | 26/81 (32.1) | p = 1.000 | 15/103 (14.5) | 40/85 (47.0) | p < 0.001 | 5/58 (8.6) | 24/49 (49.0) | p < 0.001 | 10/45 (22.2) | 16/36 (44.4) | p = 0.218 |
| II, N = 55 | 31/107 (29.0) | 24/81 (29.6) | p = 1.000 | 42/103 (40.8) | 13/85 (15.3) | p = 0.001 | 24/58 (41.4) | 7/49 (14.3) | p < 0.011 | 18/45 (40.0) | 6/36 (16.7) | p = 0.113 |
| R/M, N = 9 | 4/107 (3.7) | 5/81 (6.2) | p = 1.000 | 4/103 (3.9) | 5/85 (5.9) | p = 1.000 | 1/58 (1.7) | 3/49 (6.1) | p = 1.000 | 3/45 (6.7) | 2/36 (5.6) | p = 1.000 |
| NR, N = 69 | 43/107 (40.2) | 26/81 (32.1) | p = 1.000 | 42/104 (40.8) | 27/85 (31.8) | p = 0.904 | 28/58 (48.3) | 15/49 (30.6) | p = 0.306 | 14/45 (31.1) | 12/36 (33.3) | p = 1.000 |
| T. gondii Serotypes | |||||
|---|---|---|---|---|---|
| I–III | II | R/M | NR | p | |
| Overall n/N (%) | 55/188 (29.3) | 55/188 (29.2) | 9/188 (4.8) | 69/188 (36.7) | |
| Females—n/N (%) | 29/55 (52.7) | 31/55 (56.4) | 4/9 (44.4) | 43/69 (62.3) | p = 0.632 § |
| Age, years median (Q1–Q3) | 30 (23–44) | 31 (21–38) | 45 (34–57) | 33 (27–38) | p = 0.068 ‡ |
| Foreigners—n/N (%) | 40/55 (72.7) | 13/55 (23.6) | 5/9 (55.6) | 27/69 (39.1) | p < 0.001 § |
| Place of birth—n/N (%) | p < 0.001 § | ||||
| Europe | 2/55 (3.6) | 3/55 (5.4) | 2/9 (22.2) | 3/69 (4.4) | |
| Asia | 1/55 (1.8) | 2/55 (3.6) | 0 (0.0) | 2/69 (2.9) | |
| South America | 13/55 (23.6) | 2/55 (3.6) | 1/9 (11.1) | 4/69 (5.8) | |
| Africa | 24/55 (43.6) | 6/55 (10.9) | 2/9 (22.2) | 18/69 (26.1) | |
| Italy | 15/55 (27.3) | 42/55 (76.4) | 4/9 (44.4) | 42/69 (60.9) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caldrer, S.; Vola, A.; Ferrari, G.; Ursini, T.; Mazzi, C.; Meroni, V.; Beltrame, A. Toxoplasma gondii Serotypes in Italian and Foreign Populations: A Cross-Sectional Study Using a Homemade ELISA Test. Microorganisms 2022, 10, 1577. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10081577
Caldrer S, Vola A, Ferrari G, Ursini T, Mazzi C, Meroni V, Beltrame A. Toxoplasma gondii Serotypes in Italian and Foreign Populations: A Cross-Sectional Study Using a Homemade ELISA Test. Microorganisms. 2022; 10(8):1577. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10081577
Chicago/Turabian StyleCaldrer, Sara, Ambra Vola, Guglielmo Ferrari, Tamara Ursini, Cristina Mazzi, Valeria Meroni, and Anna Beltrame. 2022. "Toxoplasma gondii Serotypes in Italian and Foreign Populations: A Cross-Sectional Study Using a Homemade ELISA Test" Microorganisms 10, no. 8: 1577. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10081577