
Epigallocatechin Gallate Stabilized by Cyclodextrin Inactivates Influenza Virus and Human Coronavirus 229E


Abstract
:1. Introduction
2. Materials and Methods
2.1. EGCg Stabilization
2.2. Cells
2.3. Stock Virus
2.4. Cell Viability Assay
2.5. Plaque Assay
2.6. TCID50 Assay
2.7. CD-EGCg Treatment for Influenza Virus
2.8. CD-EGCg Treatment for HCoV-229E
2.9. Statistical Analysis
2.10. Reagents and Chemicals
3. Results
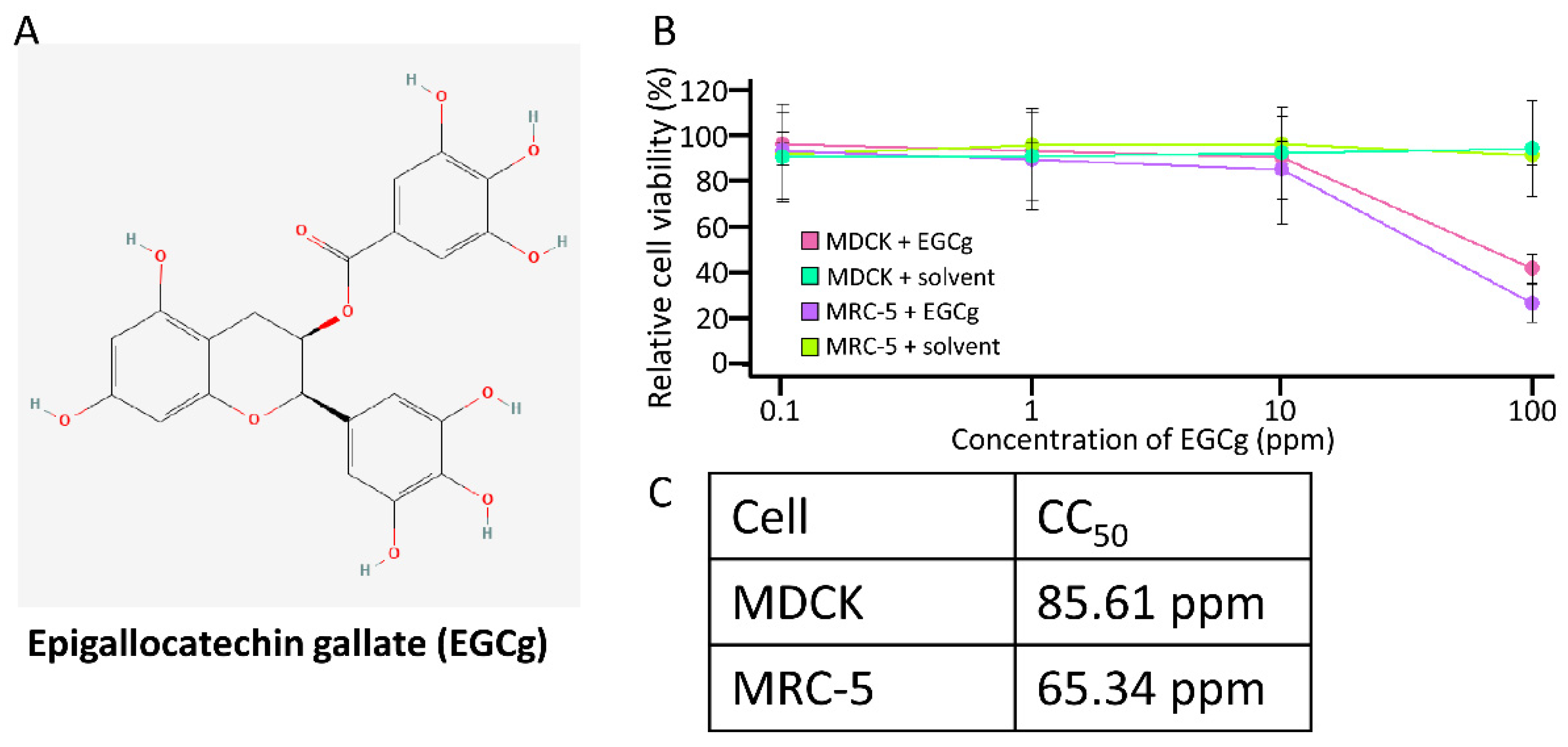
3.1. Cytotoxicity of Stabilized EGCg
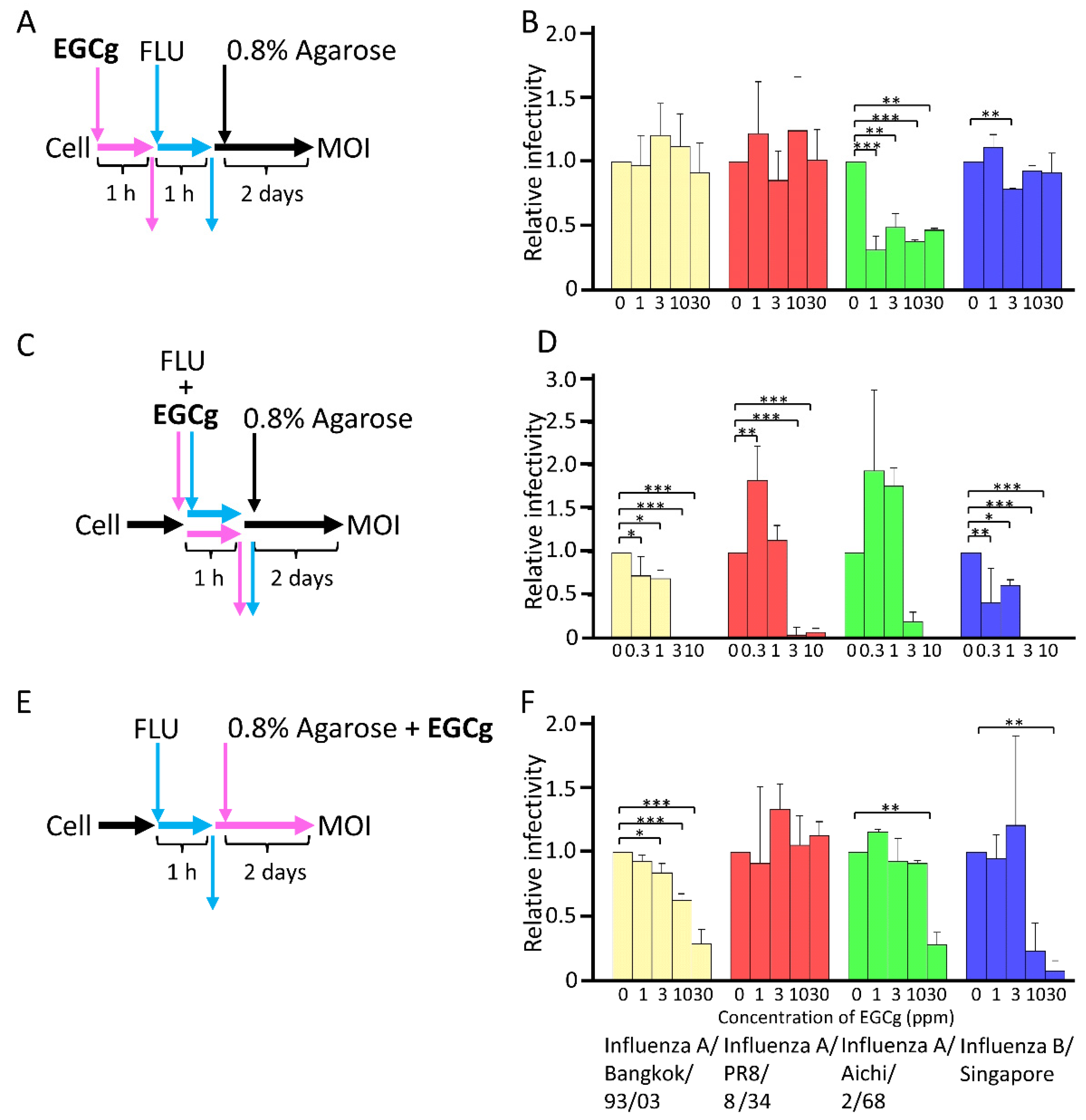
3.2. Antiviral Activity of CD-EGCg against Influenza Virus
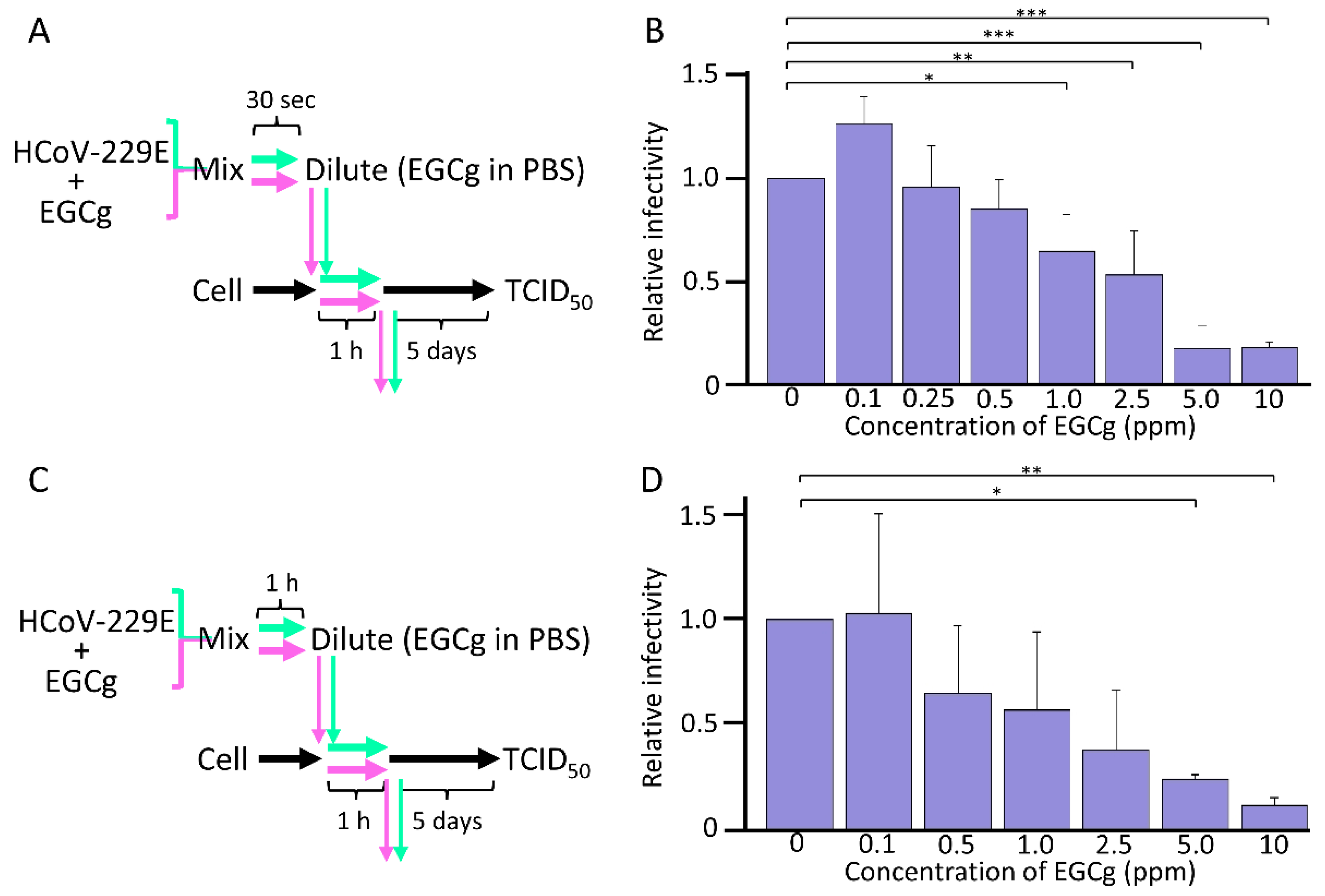
3.3. Antiviral Activity of EGCg against HCoV-229E
3.4. Point of Action of CD-EGCg
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sato, H.; Murakami, T.; Matsuura, R.; Abe, M.; Matsuoka, S.; Yashiroda, Y.; Yoshida, M.; Akari, H.; Nagasawa, Y.; Takei, M.; et al. A Novel Class of HIV-1 Inhibitors Targeting the Vpr-Induced G2-Arrest in Macrophages by New Yeast- and Cell-Based High-Throughput Screening. Viruses 2022, 14, 1321. [Google Scholar] [CrossRef] [PubMed]
- Matsuura, R.; Lo, C.W.; Wada, S.; Somei, J.; Ochiai, H.; Murakami, T.; Saito, N.; Ogawa, T.; Shinjo, A.; Benno, Y.; et al. SARS-CoV-2 Disinfection of Air and Surface Contamination by TiO2 Photocatalyst-Mediated Damage to Viral Morphology, RNA, and Protein. Viruses 2021, 13, 942. [Google Scholar] [CrossRef] [PubMed]
- Lo, C.W.; Matsuura, R.; Iimura, K.; Wada, S.; Shinjo, A.; Benno, Y.; Nakagawa, M.; Takei, M.; Aida, Y. UVC disinfects SARS-CoV-2 by induction of viral genome damage without apparent effects on viral morphology and proteins. Sci. Rep. 2021, 11, 13804. [Google Scholar] [CrossRef] [PubMed]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Pérez, M.G.; Moreira, E.D.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 mRNA COVID-19 Vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef] [PubMed]
- Baden, L.R.; El Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.A.; Rouphael, N.; Creech, C.B.; et al. Efficacy and Safety of the mRNA-1273 SARS-CoV-2 Vaccine. N. Engl. J. Med. 2021, 384, 403–416. [Google Scholar] [CrossRef]
- Zhao, Z.; Feng, M.; Wan, J.; Zheng, X.; Teng, C.; Xie, X.; Pan, W.; Hu, B.; Huang, J.; Liu, Z.; et al. Research progress of epigallocatechin-3-gallate (EGCG) on anti-pathogenic microbes and immune regulation activities. Food Funct. 2021, 12, 9607–9619. [Google Scholar] [CrossRef]
- Nakayama, M.; Suzuki, K.; Toda, M.; Okubo, S.; Hara, Y.; Shimamura, T. Inhibition of the infectivity of influenza virus by tea polyphenols. Antivir. Res. 1993, 21, 289–299. [Google Scholar] [CrossRef]
- Song, J.M.; Lee, K.H.; Seong, B.L. Antiviral effect of catechins in green tea on influenza virus. Antivir. Res. 2005, 68, 66–74. [Google Scholar] [CrossRef]
- Kuzuhara, T.; Iwai, Y.; Takahashi, H.; Hatakeyama, D.; Echigo, N. Green tea catechins inhibit the endonuclease activity of influenza A virus RNA polymerase. PLoS Curr. 2009, 1, RRN1052. [Google Scholar] [CrossRef]
- Fassina, G.; Buffa, A.; Benelli, R.; Varnier, O.E.; Noonan, D.M.; Albini, A. Polyphenolic antioxidant (-)-epigallocatechin-3-gallate from green tea as a candidate anti-HIV agent. Aids 2002, 16, 939–941. [Google Scholar] [CrossRef]
- He, W.; Li, L.X.; Liao, Q.J.; Liu, C.L.; Chen, X.L. Epigallocatechin gallate inhibits HBV DNA synthesis in a viral replication—Inducible cell line. World J. Gastroenterol. 2011, 17, 1507–1514. [Google Scholar] [CrossRef] [PubMed]
- Jang, M.; Park, R.; Park, Y.I.; Cha, Y.E.; Yamamoto, A.; Lee, J.I.; Park, J. EGCG, a green tea polyphenol, inhibits human coronavirus replication in vitro. Biochem. Biophys. Res. Commun. 2021, 547, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Ohgitani, E.; Shin-Ya, M.; Ichitani, M.; Kobayashi, M.; Takihara, T.; Kawamoto, M.; Kinugasa, H.; Mazda, O. Significant Inactivation of SARS-CoV-2 In Vitro by a Green Tea Catechin, a Catechin-Derivative, and Black Tea Galloylated Theaflavins. Molecules 2021, 26, 3572. [Google Scholar] [CrossRef]
- Li, F.; Qasim, S.; Li, D.; Dou, Q.P. Updated review on green tea polyphenol epigallocatechin-3-gallate as a cancer epigenetic regulator. Semin. Cancer Biol. 2022, 83, 335–352. [Google Scholar] [CrossRef]
- Lam, W.H.; Kazi, A.; Kuhn, D.J.; Chow, L.M.; Chan, A.S.; Dou, Q.P.; Chan, T.H. A potential prodrug for a green tea polyphenol proteasome inhibitor: Evaluation of the peracetate ester of (-)-epigallocatechin gallate [(-)-EGCG]. Bioorg. Med. Chem. 2004, 12, 5587–5593. [Google Scholar] [CrossRef] [PubMed]
- Mamenzigou, U.M.; Ikeda, T.; Morohoshi, T.; Nakayama, M.; Yui, K. The Inclusion Complex Formation of Epigallocatechin Gallate in γ-Cyclodextrin and Its Effect on the Antioxidant Activity. Trans. Mater. Res. Soc. Jpn. 2013, 38, 681–685. [Google Scholar] [CrossRef]
- Jansook, P.; Ogawa, N.; Loftsson, T. Cyclodextrins: Structure, physicochemical properties and pharmaceutical applications. Int. J. Pharm. 2018, 535, 272–284. [Google Scholar] [CrossRef]
- Paiva-Santos, A.C.; Ferreira, L.; Peixoto, D.; Silva, F.; Soares, M.J.; Zeinali, M.; Zafar, H.; Mascarenhas-Melo, F.; Raza, F.; Mazzola, P.G.; et al. Cyclodextrins as an encapsulation molecular strategy for volatile organic compounds- Pharmaceutical applications. Colloids Surf. B Biointerfaces 2022, 218, 112758. [Google Scholar] [CrossRef]
- Aree, T.; Jongrungruangchok, S. Enhancement of antioxidant activity of green tea epicatechins in β-cyclodextrin cavity: Single-crystal X-ray analysis, DFT calculation and DPPH assay. Carbohydr. Polym. 2016, 151, 1139–1151. [Google Scholar] [CrossRef]
- Sawamura, R.; Shimizu, T.; Sun, Y.; Yasukawa, K.; Miura, M.; Toriyama, M.; Motohashi, S.; Watanabe, W.; Konno, K.; Kurokawa, M. In vitro and in vivo anti-influenza virus activity of diarylheptanoids isolated from Alpinia officinarum. Antivir. Chem. Chemother. 2010, 21, 33–41. [Google Scholar] [CrossRef]
- Yamada, K.; Koyama, H.; Hagiwara, K.; Ueda, A.; Sasaki, Y.; Kanesashi, S.N.; Ueno, R.; Nakamura, H.K.; Kuwata, K.; Shimizu, K.; et al. Identification of a novel compound with antiviral activity against influenza A virus depending on PA subunit of viral RNA polymerase. Microbes Infect. 2012, 14, 740–747. [Google Scholar] [CrossRef] [PubMed]
- PubChem Identifier: CID 65064. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/65064 (accessed on 22 August 2022).
- Ntamo, Y.; Jack, B.; Ziqubu, K.; Mazibuko-Mbeje, S.E.; Nkambule, B.B.; Nyambuya, T.M.; Mabhida, S.E.; Hanser, S.; Orlando, P.; Tiano, L.; et al. Epigallocatechin gallate as a nutraceutical to potentially target the metabolic syndrome: Novel insights into therapeutic effects beyond its antioxidant and anti-inflammatory properties. Crit. Rev. Food Sci. Nutr. 2022. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Virus | Strain | EGCg Treatment Pre-Adsorption | EGCg Treatment during Adsorption | EGCg Treatment Post-Adsorption |
|---|---|---|---|---|
| Influenza virus | A/Bangkok/93/03 | >30 | 1.41 ± 0.17 | 19.32 ± 1.79 |
| A/PR8/8/34 | >30 | 2.79 ± 0.09 | >30 | |
| A/Aichi/2/68 | >30 | 2.78 ± 0.23 | 22.89 ± 1.44 | |
| B/Singapore | >30 | 0.93 ± 0.35 | 11.08 ± 1.56 | |
| HCoV-229E | N. T. 1 | 2.5 (Short contact) 0.5 (Long contact) | N. T. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matsuura, R.; Kawamura, A.; Matsumoto, Y.; Iida, Y.; Kanayama, M.; Kurokawa, M.; Aida, Y. Epigallocatechin Gallate Stabilized by Cyclodextrin Inactivates Influenza Virus and Human Coronavirus 229E. Microorganisms 2022, 10, 1796. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10091796
Matsuura R, Kawamura A, Matsumoto Y, Iida Y, Kanayama M, Kurokawa M, Aida Y. Epigallocatechin Gallate Stabilized by Cyclodextrin Inactivates Influenza Virus and Human Coronavirus 229E. Microorganisms. 2022; 10(9):1796. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10091796
Chicago/Turabian StyleMatsuura, Ryosuke, Arisa Kawamura, Yasunobu Matsumoto, Yoshiki Iida, Masanori Kanayama, Masahiko Kurokawa, and Yoko Aida. 2022. "Epigallocatechin Gallate Stabilized by Cyclodextrin Inactivates Influenza Virus and Human Coronavirus 229E" Microorganisms 10, no. 9: 1796. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10091796