
Characterization of Phytopythium Species Involved in the Establishment and Development of Kiwifruit Vine Decline Syndrome
 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Oomycete Isolation
2.2. Molecular Identification
2.3. Morphological Observation
2.4. Effect of Temperature and pH on In Vitro Growth
2.5. Pathogenicity Test
2.6. Statistical Analysis
3. Results
3.1. Oomycete Isolation and Identification
3.2. Morphological Observations
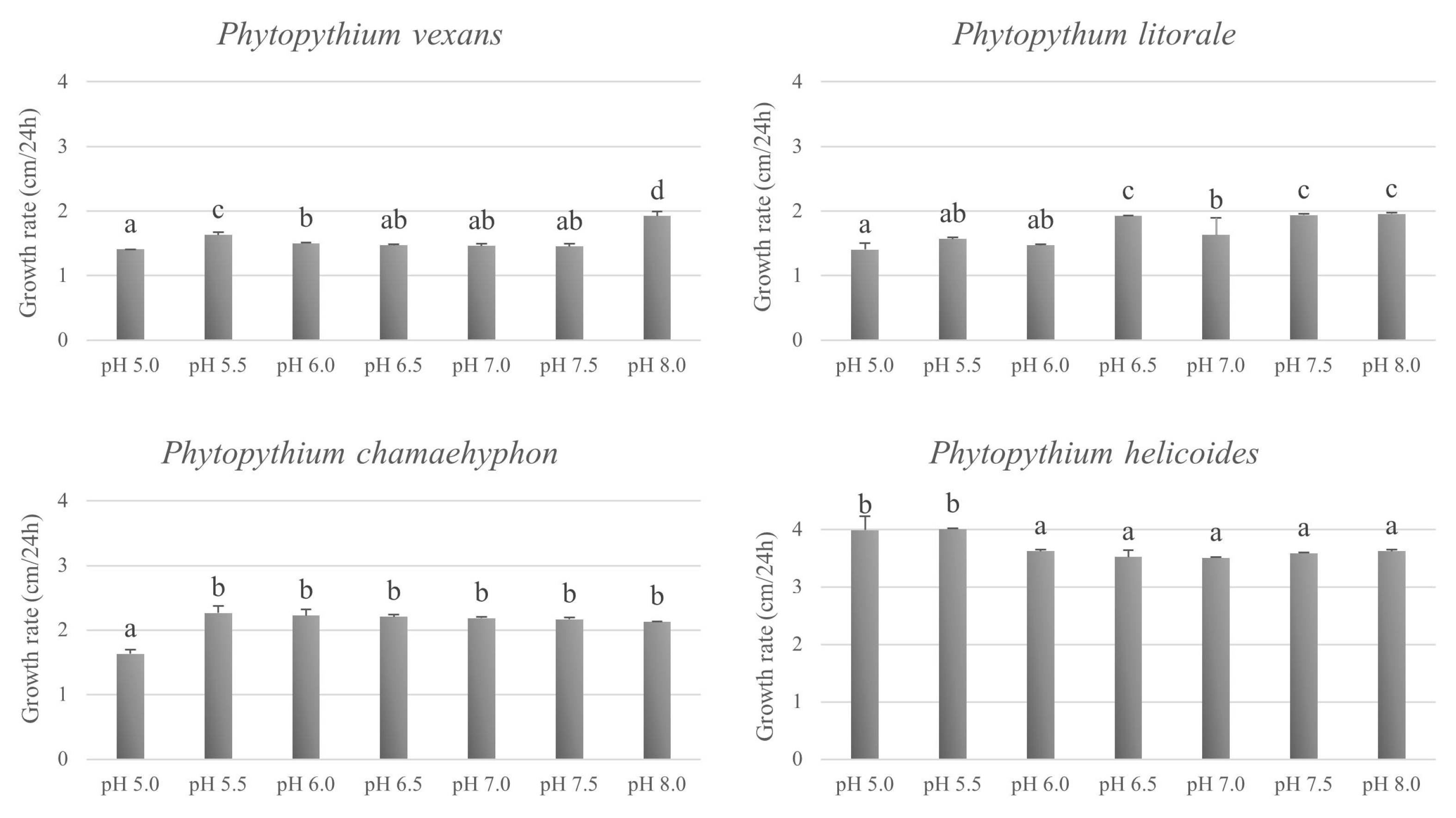
3.3. Effect of Temperature and pH on In Vitro Growth
3.4. Pathogenicity Test
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Sorrenti, G.; Tacconi, G.; Tosi, L.; Vittone, G.; Nari, L.; Savian, F.; Saro, S.; Ermacora, P.; Graziani, S.; Toselli, M. Avanza la “moria del kiwi”: Evoluzione e primi riscontri della ricerca. Riv. Fruttic. 2019, 2, 34–42. [Google Scholar]
- Balestra, G.M.; Costa, G. La situazione in Italia: Evoluzione varietale sì, ma con tante avversità. Riv. Fruttic. 2020, 82, 10–16. [Google Scholar]
- Savian, F.; Ginaldi, F.; Musetti, R.; Sandrin, N.; Tarquini, G.; Pagliari, L.; Firrao, G.; Martini, M.; Ermacora, P. Studies on the aetiology of kiwifruit decline: Interaction between soil-borne pathogens and waterlogging. Plant Soil 2020, 456, 113–128. [Google Scholar] [CrossRef]
- Xiloyannis, C.; Baroni, G.; Bovo, M.; Angelini, P.; Pezzarossa, B.; Pagliai, M. L’irrigazione per scorrimento nel Veronese: Spreco di acqua e di concimi; effetti sulla porosità del terreno. Riv. Fruttic. 1992, 6, 55–62. [Google Scholar]
- Tacconi, G.; Paltrinieri, S.; Mejia, J.F.; Fuentealba, S.P.; Bertaccini, A.; Tosi, L.; Giacopini, A.; Mazzucchi, U.; Favaron, F.; Sella, L.; et al. Vine decline in kiwifruit: Climate change and effect on waterlogging and Phytophthora in north Italy. Acta Hortic. 2015, 1096, 93–97. [Google Scholar] [CrossRef]
- Akilli, S.; Serçe, Ç.U.; Zekaİ Katircioğlu, Y.; Karakaya, A.; Maden, S. Involvement of Phytophthora citrophthora in kiwifruit decline in Turkey. J. Phytopathol. 2011, 159, 579–581. [Google Scholar] [CrossRef]
- Kurbetli, İ.; Ozan, S. Occurrence of Phytophthora root and stem rot of kiwifruit in Turkey. J. Phytopathol. 2013, 161, 887–889. [Google Scholar] [CrossRef]
- Reid, J.B.; Tate, K.G.; Brown, N.S. Effects of flooding and alluvium deposition on kiwifruit (Actinidia deliciosa). N. Z. J. Crop Hortic. Sci. 1991, 19, 247–257. [Google Scholar] [CrossRef]
- McAneney, K.J.; Clough, A.; Green, A.; Harris, B.; Richardson, A. Waterlogging and vine death at Kerikeri. N. Z. Kiwifruit 1989, 56, 15. [Google Scholar]
- Prencipe, S.; Savian, F.; Nari, L.; Ermacora, P.; Spadaro, D.; Martini, M. First report of Phytopythium vexans causing decline syndrome of Actinidia deliciosa ‘Hayward’ in Italy. Plant Dis. 2020, 104, 2032. [Google Scholar] [CrossRef] [Green Version]
- Donati, I.; Cellini, A.; Sangiorgio, D.; Caldera, E.; Sorrenti, G.; Spinelli, F. Pathogens associated to kiwifruit vine decline in Italy. Agriculture 2020, 10, 119. [Google Scholar] [CrossRef] [Green Version]
- Bardi, L. Early kiwifruit decline: A soil-borne disease syndrome or a climate change effect on plant–soil relations? Front. Agron. 2020, 2, 3. [Google Scholar] [CrossRef]
- Tosi, L.; Tacconi, G.; Giacopini, A. La moria del kiwi, situazione e prospettive. L’Informatore Agrar. 2015, 44, 67–70. [Google Scholar]
- Sorrenti, G.; Toselli, M.; Reggidori, G.; Spinelli, F.; Tosi, L.; Giacopini, A.; Tacconi, G. Implicazioni della gestione idrica nella “moria del kiwi” del veronese. Riv. Fruttic. 2016, 3, 45–51. [Google Scholar]
- Bardi, L.; Nari, L.; Morone, C.; Faga, M.G.; Malusà, E. Possible Role of High Temperature and Soil Biological Fertility on Kiwifruit Early Decline Syndrome. Front. Agron. 2020, 2, 580659. [Google Scholar] [CrossRef]
- Tacconi, G.; Giacopini, A.; Vittone, G.; Nari, L.; Spadaro, D.; Savian, F.; Ermacora, P.; Saro, S.; Morone, C.; Bardi, L.; et al. “Moria del kiwi”: Situazione disastrosa al nord, preoccupante nel resto d’Italia. Kiwi Inf. 2019, 2, 32–37. [Google Scholar]
- Savian, F.; Prencipe, S.; Filippini, N.; Nari, L.; Martini, M.; Ermacora, P.; Spadaro, D. Pathogenicity of Phytopythium chamaehyphon: A New Player in Kiwifruit Vine Decline Syndrome of Actinidia chinensis var. deliciosa ‘Hayward’ in Italy. Plant Dis. 2021, 105, 2781–2784. [Google Scholar] [CrossRef]
- Spigaglia, P.; Barbanti, F.; Marocchi, F.; Mastroleo, M. Clostridium bifermentans and C. subterminale are associated with kiwifruit vine decline, known as moria, in Italy. Plant Pathol. 2020, 69, 765–774. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.D.; Lee, S.B.; Taylor, J.W. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Cooke, D.E.L.; Duncan, J.M. Phylogenetic analysis of Phytophthora species based on ITS1and ITS2 sequences of the ribosomal RNA gene repeat. Mycol. Res. 1997, 101, 667–677. [Google Scholar] [CrossRef]
- Baten, M.A.; Asano, T.; Motohashi, K.; Ishiguro, Y.; Rahman, M.Z.; Inaba, S.; Suga, H.; Kageyama, K. Phylogenetic relationships among Phytopythium species, and re-evaluation of Phytopythium fagopyri comb. nov., recovered from damped-off buckwheat seedlings in Japan. Mycol. Prog. 2014, 13, 1003. [Google Scholar] [CrossRef]
- Robideau, G.P.; de Cock, A.W.A.M.; Coffey, M.D.; Voglmayr, H.; Brouwer, H.; Bala, K.; Chitty, D.W.; Désaulniers, N.; Eggertson, Q.A.; Gachon, C.M.M.; et al. DNA barcoding of oomycetes with cytochrome c oxidase subunit I and internal transcribed spacer. Mol. Ecol. Resour. 2011, 11, 1002–1011. [Google Scholar] [CrossRef] [PubMed]
- de Cock, A.W.A.M.; Lodhi, A.M.; Rintoul, T.L.; Bala, K.L.; Robideau, G.P.; Gloria Abad, Z.; Coffey, M.D.; Shahzad, S.; Lévesque, C.A. Phytopythium: Molecular phylogeny and systematics. Peersonia 2015, 34, 25–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crous, P.W.; Verkley, G.J.M.; Groenewald, J.Z.; Samson, R.A. Fungal Biodiversity. In CBS Laboratory Manual Series 1; CBS-KNAW Fungal Biodiversity Centre: Utrecht, The Netherlands, 2009; p. 269. [Google Scholar]
- de Cock, A.W.A.M.; Lévesque, C.A. New species of Pythium and Phytophthora. Stud. Mycol. 2004, 50, 481–487. [Google Scholar]
- Adhikari, B.N.; Hamilton, J.P.; Zerillo, M.M.; Tisserat, N.; Lévesque, C.A.; Buell, C.R. Comparative genomics reveals insight into virulence strategies of plant pathogenic oomycetes. PLoS ONE 2013, 8, e75072. [Google Scholar] [CrossRef]
- Polat, Z.; Awan, Q.N.; Hussain, M.; Akgul, D.S. First Report of Phytopythium vexans causing root and collar rot of kiwifruit in Turkey. Plant Dis. 2017, 101, 1058. [Google Scholar] [CrossRef]
- Baten, M.A.; Li, M.; Motohashi, K.; Ishiguro, Y.; Rahman, M.Z.; Suga, H.; Kageyama, K. Two new species, Phytopythium iriomotense sp. nov. and P. aichiense sp. nov., isolated from river water and water purification sludge in Japan. Mycol. Prog. 2015, 14, 2. [Google Scholar] [CrossRef]
- Rodríguez Padrón, C.; Siverio, F.; Pérez-Sierra, A.; Rodríguez, A. Isolation and pathogenicity of Phytophthora species and Phytopythium vexans recovered from avocado orchards in the Canary Islands, including Phytophthora niederhauserii as a new pathogen of avocado. Phytopathol. Mediterr. 2018, 57, 89–106. [Google Scholar]
- Kageyama, K.; Senda, M.; Asano, T.; Suga, H.K.; Ishiguro, K. Intro isolate heterogeneity of the ITS region of rDNA in Pythium helicoides. Mycol. Res. 2007, 111, 416–423. [Google Scholar] [CrossRef]
- Chen, X.R.; Liu, B.B.; Xing, Y.P.; Cheng, B.P.; Liu, M.L.; Tong, Y.H.; Xu, J.Y. Identification and characterization of Phytopythium helicoides causing stem rot of Shatangju mandarin seedlings in China. Eur. J. Plant Pathol. 2016, 146, 715–727. [Google Scholar] [CrossRef]
- Marano, A.V.; Jesus, A.L.; De Souza, J.I.; Leãno, E.M.; James, T.Y.; Jerônimo, G.H.; de Cock, A.W.A.M.; Pires-Zottarelli, C.L.A. A new combination in Phytopythium: P. kandeliae (Oomycetes, Straminipila). Mycosphere 2014, 5, 510–522. [Google Scholar] [CrossRef]
- Bouket, C.A.; Babai-Ahari, A.; Arzanlou, M.; Tojo, M. Morphological and molecular characterization of Phytopythium litorale and Pp. oedochilum from Iran. Nova Hedwig. 2016, 102, 257–270. [Google Scholar] [CrossRef]
- Sideris, C.P. Taxonomic studies in the family Pythiaceae II. Pythium. Mycologia 1932, 24, 14–61. [Google Scholar] [CrossRef]
- Bennett, R.M.; Bora, N.; Dedeles, G.R.; Thines, M. Phytopythium leanoi sp. nov. and Phytopythium dogmae sp. nov., Phytopythium species associated with mangrove leaf litter from the Philippines. Acta Mycol. 2017, 52, 1103. [Google Scholar] [CrossRef] [Green Version]
- Nam, B.; Choi, Y.J. Phytopythium and Pythium Species (Oomycota) isolated from freshwater environments of Korea. Mycobiology 2019, 47, 261–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derviş, S.; Türkölmez, S.; Çiftçi, O.; Özer, G.; Ulubaş Serçe, Ç.; Dikilitas, M. Phytopythium litorale: A novel killer pathogen of plane (Platanus orientalis) causing canker stain and root and collar rot. Plant Dis. 2020, 104, 2642–2648. [Google Scholar] [CrossRef]
- Li, Y.; Feng, Y.; Wu, C.; Xue, J.; Jiao, B.; Li, B.; Dai, T. First Report of Phytopythium litorale Causing Crown and Root Rot on Rhododendron pulchrum in China. Plant Dis. 2021, 105, 4173. [Google Scholar] [CrossRef]
- Jabiri, S.; Bahra, C.; MacLean, D.; Radouane, N.; Barka, E.A.; Bendriss Amraoui, M.; Lahlali, R. Phytopythium vexans Associated with Apple and Pear Decline in the Saïss Plain of Morocco. Microorganisms 2021, 9, 1916. [Google Scholar] [CrossRef] [PubMed]
- Beluzán, F.; Miarnau, X.; Torguet, L.; Armengol, J.; Abad-Campos, P. Survey of Oomycetes Associated with Root and Crown Rot of Almond in Spain and Pathogenicity of Phytophthora niederhauserii and Phytopythium vexans to ‘Garnem’ Rootstock. Agriculture 2022, 12, 294. [Google Scholar] [CrossRef]
- Wang, K.X.; Xie, Y.L.; Yuan, G.Q.; Li, Q.Q.; Lin, W. First report of root and collar rot caused by Phytopythium helicoides on kiwifruit (Actinidia chinensis). Plant Dis. 2015, 99, 725–726. [Google Scholar] [CrossRef]
- Tao, Y.H.; Zeng, F.; Ho, H.H.; Wei, J.; Wu, Y.; Yang, L.; He, Y. Pythium vexans causing stem rot of Dendrobium in Yunnan Province, China. J. Phytopathol. 2011, 159, 255–259. [Google Scholar] [CrossRef]
- Afandi, A.; Murayama, E.; Hieno, A.; Suga, H.; Kageyama, K. Population structures of the waterborne plant pathogen Phytopythium helicoides reveal its possible origins and transmission modes in Japan. PLoS ONE 2018, 13, e0209667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nechwatal, J.; Mendgen, K. Pythium litorale sp. nov., a new species from the littoral of Lake Constance, Germany. FEMS Microbiol. Lett. 2006, 255, 96–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cantrell, H.F.; Dowler, W.M. Effects of temperature and pH on growth and composition of Pythium irregulare and Pythium vexans. Mycologia 1971, 63, 31–37. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain * | Species | Cultivar | Year of Isolation | GenBank Accession Numbers ** | ||
|---|---|---|---|---|---|---|
| ITS | LSU | COI | ||||
| PH1 | Phytopythium vexans | Hayward | 2016 | OL891590 | OL957094 | ON228590 |
| PH6 | Phytopythium chamaehyphon | Hayward | 2016 | OL891528 | OL957144 | ON228528 |
| PH3 | Phytopythium vexans | Hayward | 2016 | MN510425 * | MN510427 | MN510423 |
| PH2 | Phytopythium vexans | Hayward | 2016 | OL891591 | OL957095 | ON228591 |
| GBI | Phytopythium vexans | Hayward | 2016 | OL891578 | OL957096 | ON228578 |
| 4SRE1 | Phytopythium vexans | Hayward | 2016 | OL891579 | OL957097 | ON228579 |
| 4SRE2 | Phytopythium vexans | Hayward | 2016 | OL891580 | OL957098 | ON228580 |
| 4SBE_2C1 | Phytopythium vexans | Hayward | 2016 | OL891581 | OL957099 | ON228581 |
| 4SBE_C2A | Phytopythium litorale | Hayward | 2016 | OL891534 | OL957153 | ON228534 |
| 4SBE_C2B | Phytopythium litorale | Hayward | 2016 | OL891536 | OL957154 | ON228536 |
| 4SBE_C2D | Phytopythium litorale | Hayward | 2016 | OL891533 | OL957155 | ON228533 |
| 4SBE_1C1 | Phytopythium vexans | Hayward | 2016 | OL891587 | OL957100 | ON228587 |
| 4SBE_C1A | Phytopythium vexans | Hayward | 2016 | OL891586 | OL957101 | ON228586 |
| 4SBE_C2C | Phytopythium litorale | Hayward | 2016 | OL891535 | OL957162 | ON228535 |
| 4SBE_4C2 | Phytopythium vexans | Hayward | 2016 | OL891588 | OL957102 | ON228588 |
| 4SBE_3C2 | Phytopythium vexans | Hayward | 2016 | OL891589 | OL957103 | ON228589 |
| 4/16_DRE | Phytopythium chamaehyphon | Hayward | 2016 | OL891529 | OL957145 | ON228529 |
| 5/16_CBE | Phytopythium vexans | Hayward | 2016 | OL891582 | OL957104 | ON228582 |
| 6/16_BRE | Phytopythium vexans | Soreli | 2016 | OL891583 | OL957105 | ON228583 |
| 11/16_AB1 | Phytopythium chamaehyphon | Hayward | 2016 | OL891531 | OL957146 | ON228531 |
| 12/16_DR2 | Phytopythium vexans | Hayward | 2016 | MN510426 | MN510428 | MN510424 |
| 12/16_CR1 | Phytopythium vexans | Hayward | 2016 | OL891584 | OL957106 | ON228584 |
| 12/16_AB1 | Phytopythium vexans | Hayward | 2016 | OL891585 | OL957107 | ON228585 |
| 13/16_DR2 | Phytopythium vexans | Hayward | 2016 | OL891564 | OL957108 | ON228564 |
| 13/16_DR3 | Phytopythium vexans | Hayward | 2016 | OL891565 | OL957109 | ON228565 |
| 10/16_BR | Phytopythium chamaehyphon | Hayward | 2016 | OL891530 | OL957147 | ON228530 |
| PP1 | Phytopythium vexans | Hayward | 2018 | OL891571 | OL957110 | ON228571 |
| PP2 | Phytopythium vexans | Hayward | 2018 | OL891572 | OL957111 | ON228572 |
| PP3 | Phytopythium vexans | Hayward | 2018 | OL891573 | OL957112 | ON228573 |
| PP4 | Phytopythium vexans | Hayward | 2018 | OL891574 | OL957113 | ON228574 |
| PP5 | Phytopythium vexans | Hayward | 2018 | OL891575 | OL957114 | ON228575 |
| PP6 | Phytopythium vexans | Hayward | 2018 | OL891576 | OL957115 | ON228576 |
| PP8 | Phytopythium vexans | Hayward | 2018 | OL891577 | OL957116 | ON228577 |
| PPA | Phytopythium vexans | Hayward | 2018 | OL891566 | OL957117 | ON228566 |
| PPC | Phytopythium vexans | Hayward | 2018 | OL891567 | OL957118 | ON228567 |
| PPD | Phytopythium vexans | Hayward | 2018 | OL891568 | OL957119 | ON228568 |
| PPE | Phytopythium vexans | Hayward | 2018 | OL891569 | OL957120 | ON228569 |
| PPF | Phytopythium vexans | Hayward | 2018 | OL891570 | OL957121 | ON228570 |
| CA1 | Phytopythium helicoides | Hayward | 2019 | OL891523 | OL957151 | ON228523 |
| CA2 | Phytopythium helicoides | Hayward | 2019 | OL891524 | OL957152 | ON228524 |
| CA3 | Phytopythium chamaehyphon | Hayward | 2019 | OL891525 | OL957148 | ON228525 |
| CA4 | Phytopythium chamaehyphon | Hayward | 2019 | OL891526 | OL957149 | ON228526 |
| CA5 | Phytopythium chamaehyphon | Hayward | 2019 | OL891527 | OL957150 | ON228527 |
| R1A | Phytopythium vexans | Hayward | 2019 | OL891543 | OL957122 | ON228543 |
| R1B | Phytopythium vexans | Hayward | 2019 | OL891544 | OL957123 | ON228544 |
| R1C | Phytopythium vexans | Hayward | 2019 | OL891545 | OL957124 | ON228545 |
| R1D | Phytopythium vexans | Hayward | 2019 | OL891546 | OL957125 | ON228546 |
| R1E | Phytopythium vexans | Hayward | 2019 | OL891547 | OL957126 | ON228547 |
| R1F | Phytopythium vexans | Hayward | 2019 | OL891548 | OL957127 | ON228548 |
| R1G | Phytopythium vexans | Hayward | 2019 | OL891549 | OL957128 | ON228549 |
| R1H | Phytopythium vexans | Hayward | 2019 | OL891550 | OL957129 | ON228550 |
| R3A | Phytopythium litorale | Hayward | 2019 | OL891532 | OL957156 | ON228532 |
| T4A | Phytopythium vexans | Hayward | 2019 | OL891562 | OL957130 | ON228562 |
| T4B | Phytopythium vexans | Hayward | 2019 | OL891563 | OL957131 | ON228563 |
| R7A | Phytopythium vexans | Hayward | 2019 | OL891551 | OL957132 | ON228551 |
| R7B | Phytopythium vexans | Hayward | 2019 | OL891552 | OL957133 | ON228552 |
| R7C | Phytopythium vexans | Hayward | 2019 | OL891553 | OL957134 | ON228553 |
| R7D | Phytopythium vexans | Hayward | 2019 | OL891554 | OL957135 | ON228554 |
| R7E | Phytopythium vexans | Hayward | 2019 | OL891555 | OL957136 | ON228555 |
| R7F | Phytopythium vexans | Hayward | 2019 | OL891556 | OL957137 | ON228556 |
| P8A | Phytopythium vexans | Soreli | 2019 | OL891557 | OL957138 | ON228557 |
| P8B | Phytopythium vexans | Soreli | 2019 | OL891558 | OL957139 | ON228558 |
| P8D | Phytopythium vexans | Soreli | 2019 | OL891559 | OL957140 | ON228559 |
| P8E | Phytopythium vexans | Soreli | 2019 | OL891560 | OL957141 | ON228560 |
| P8F | Phytopythium vexans | Soreli | 2019 | OL891561 | OL957142 | ON228561 |
| P8G | Phytopythium litorale | Soreli | 2019 | OL891537 | OL957157 | ON228537 |
| P8H | Phytopythium litorale | Soreli | 2019 | OL891538 | OL957158 | ON228538 |
| P8I | Phytopythium litorale | Soreli | 2019 | OL891539 | OL957159 | ON228539 |
| R10A | Phytopythium litorale | Dong Hong | 2019 | OL891540 | OL957160 | ON228540 |
| R10B | Phytopythium litorale | Dong Hong | 2019 | OL891541 | OL957161 | ON228541 |
| RE3 | Phytopythium vexans | Hayward | 2019 | OL891542 | OL957143 | ON228542 |
| DS 12 dpi | DS 26 dpi | |||||
|---|---|---|---|---|---|---|
| Treatment | Mean ± SD | Tukey Test | Mean ± SD | Tukey Test | ||
| P. vexans strain PP1 | 1.50 ± 0.50 | bc | 2.17 ± 0.58 | b | ||
| P. chamaehyphon strain PH6 | 1.00 ± 0.50 | ab | 2.67 ± 0.29 | bc | ||
| P. helicoides strain CA2 | 3.67 ± 0.58 | d | 4.00 ± 0.00 | e | ||
| P. litorale strain R3A | 2.17 ± 0.76 | c | 3.17 ± 0.76 | cd | ||
| Positive control (infected soil) | 3.50 ± 0.87 | d | 3.83 ± 0.29 | de | ||
| Negative control (healthy soil) | 0.00 | - | a | 0.00 | - | a |
| Treatment | Mean ± SD | Tukey Test |
|---|---|---|
| P. vexans strain PP1 | 23.94 ± 9.16 | b |
| P. chamaehyphon strain PH6 | 26.06 ± 2.77 | b |
| P. helicoides strain CA2 | 7.35 ± 0.76 | a |
| P. litorale strain R3A | 17.13 ± 0.90 | ab |
| Positive control (infected soil) | 13.43 ± 2.91 | a |
| Negative control (healthy soil) | 40.05 ± 8.32 | c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prencipe, S.; Schiavon, G.; Rosati, M.; Nari, L.; Schena, L.; Spadaro, D. Characterization of Phytopythium Species Involved in the Establishment and Development of Kiwifruit Vine Decline Syndrome. Microorganisms 2023, 11, 216. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms11010216
Prencipe S, Schiavon G, Rosati M, Nari L, Schena L, Spadaro D. Characterization of Phytopythium Species Involved in the Establishment and Development of Kiwifruit Vine Decline Syndrome. Microorganisms. 2023; 11(1):216. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms11010216
Chicago/Turabian StylePrencipe, Simona, Giada Schiavon, Marco Rosati, Luca Nari, Leonardo Schena, and Davide Spadaro. 2023. "Characterization of Phytopythium Species Involved in the Establishment and Development of Kiwifruit Vine Decline Syndrome" Microorganisms 11, no. 1: 216. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms11010216