
Fungus under a Changing Climate: Modeling the Current and Future Global Distribution of Fusarium oxysporum Using Geographical Information System Data

Abstract
:1. Introduction
2. Materials and Methods
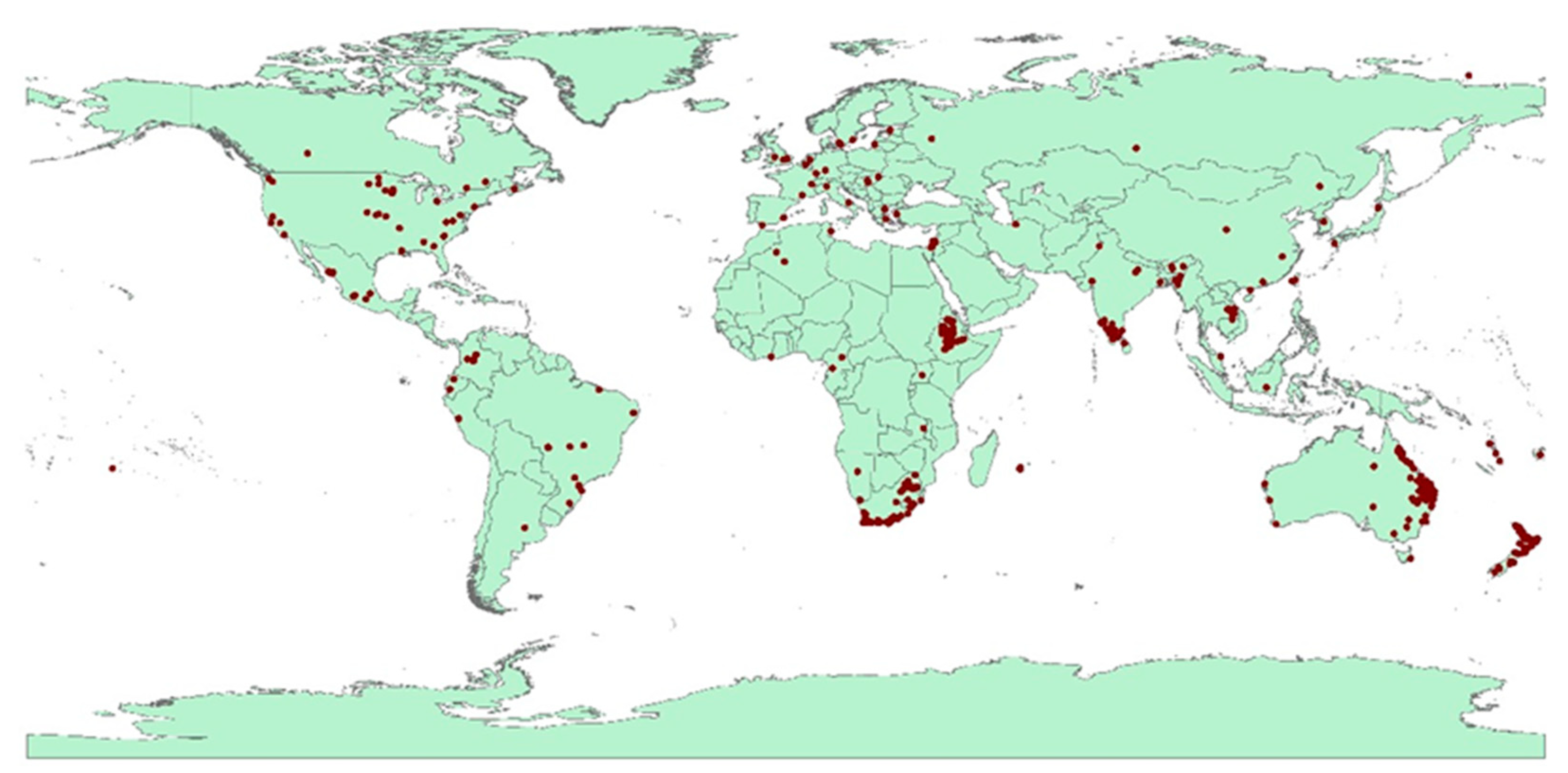
2.1. Occurrence Records for F. oxysporum
2.2. Climatological Data
2.3. Modeling and Data Analysis
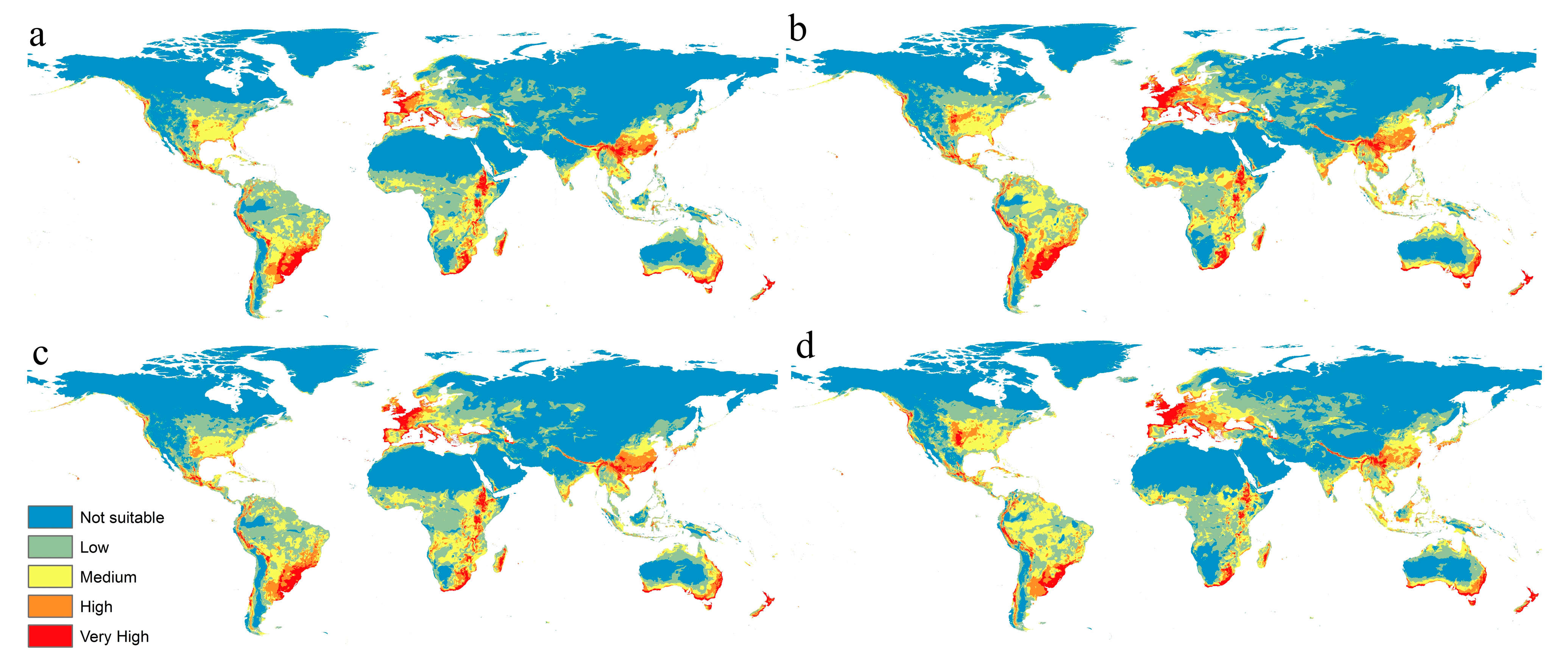
3. Results
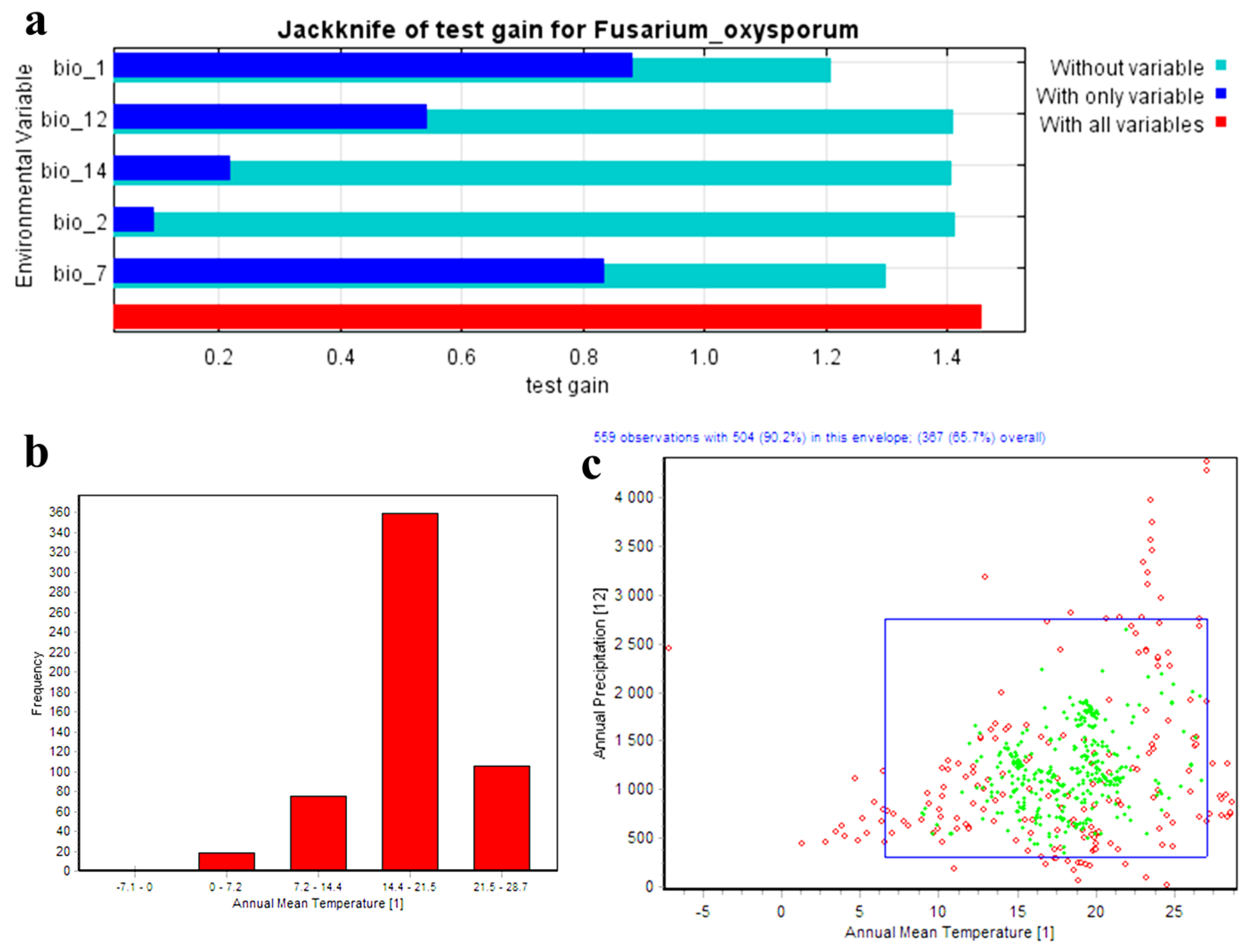
3.1. Model Accuracy and Environmental Variables’ Effects
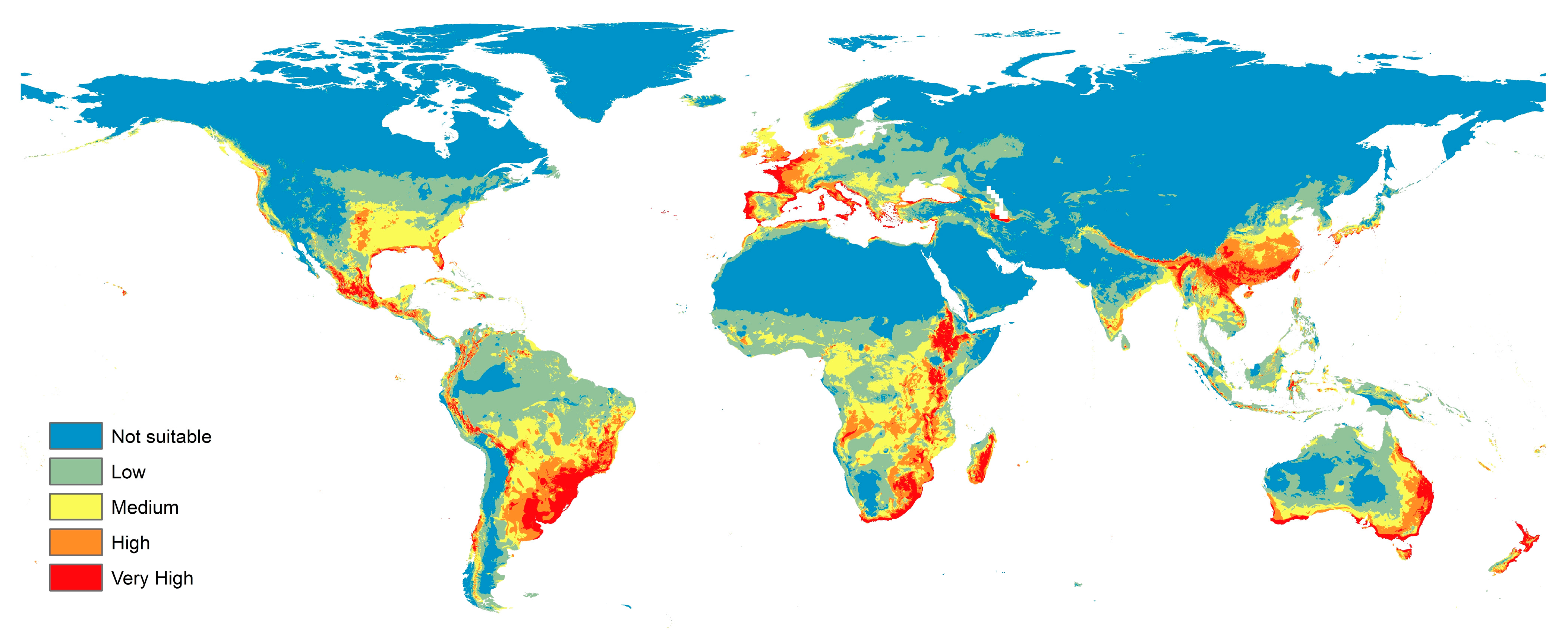
3.2. Current Prediction Map of F. oxysporum Status
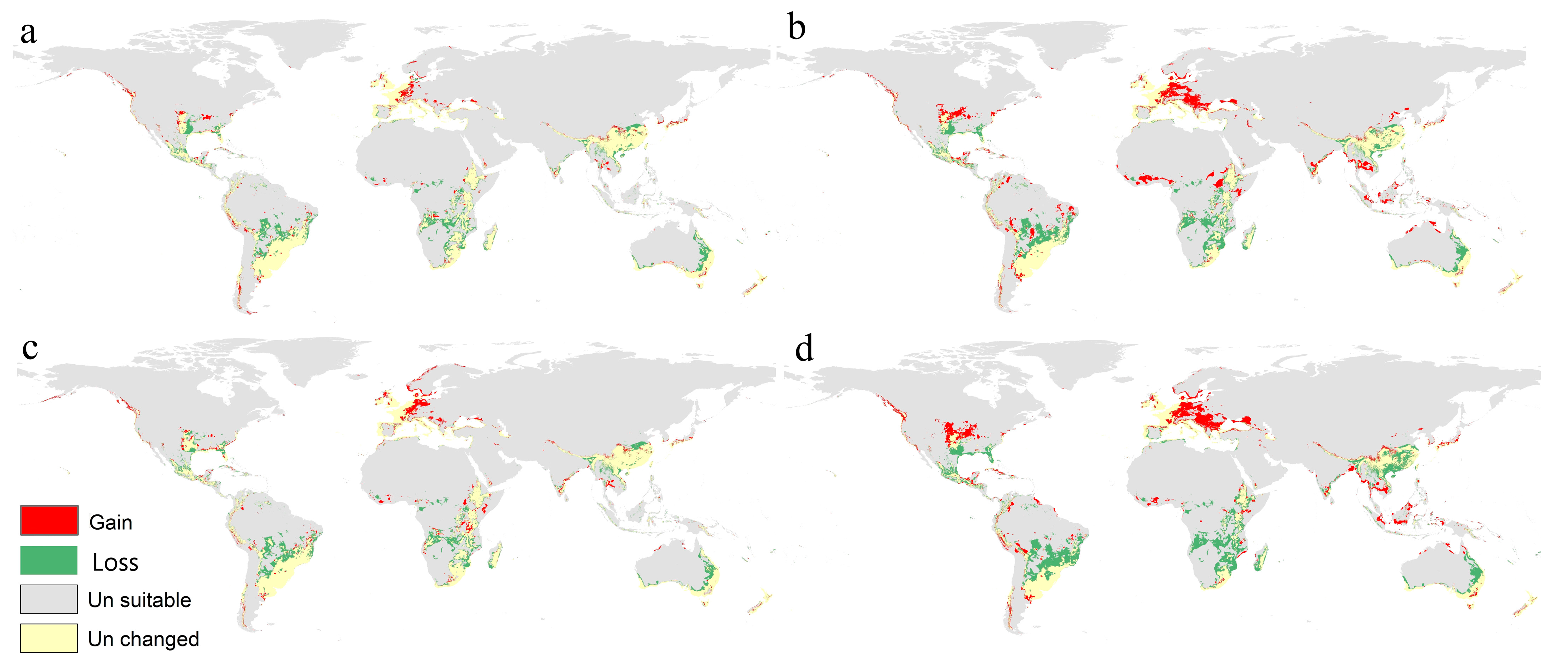
3.3. Prediction of F. oxysporum Status under Different Climate Change Scenarios
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tiedje, J.M.; Bruns, M.A.; Casadevall, A.; Criddle, C.S.; Eloe-Fadrosh, E.; Karl, D.M.; Nguyen, N.K.; Zhou, J. Microbes and Climate Change: A Research Prospectus for the Future. mBio 2022, 13, e0080022. [Google Scholar] [CrossRef] [PubMed]
- IPCC. Climate Change. Impacts, Adaptation, and Vulnerability. Contribution of Working Group II to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; In press; Cambridge University Press: Cambridge, UK, 2022. [Google Scholar]
- Parikka, P.; Hakala, K.; Tiilikkala, K. Expected shifts in Fusarium species’ composition on cereal grain in Northern Europe due to climatic change. Food Addit. Contam. Part A 2012, 29, 1543–1555. [Google Scholar] [CrossRef] [PubMed]
- Crowther, T.W.; van den Hoogen, J.; Wan, J.; Mayes, M.A.; Keiser, A.D.; Mo, L.; Averill, C.; Maynard, D.S. The global soil community and its influence on biogeochemistry. Science 2019, 365, eaav0550. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Zhou, X.; Hale, L.; Yuan, M.; Ning, D.; Feng, J.; Shi, Z.; Li, Z.; Feng, B.; Gao, Q.; et al. Climate warming accelerates temporal scaling of grassland soil microbial biodiversity. Nat. Ecol. Evol. 2019, 3, 612–619. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Xue, K.; Xie, J.; Deng, Y.; Wu, L.; Cheng, X.; Fei, S.; Deng, S.; He, Z.; Van Nostrand, J.D.; et al. Microbial mediation of carbon-cycle feedbacks to climate warming. Nat. Clim. Chang. 2012, 2, 106–110. [Google Scholar]
- Cavicchioli, R.; Ripple, W.J.; Timmis, K.N.; Azam, F.; Bakken, L.R.; Baylis, M.; Behrenfeld, M.J.; Boetius, A.; Boyd, P.W.; Classen, A.T.; et al. Scientists’ warning to humanity: Microorganisms and climate change. Nat. Rev. Microbiol. 2019, 17, 569–586. [Google Scholar] [CrossRef]
- Burgess, L.W.; Bryden, W.L. Fusarium: A ubiquitous fungus of global significance. Microbiol. Aust. 2012, 33, 22–25. [Google Scholar] [CrossRef]
- Desjardins, A.E. Fusarium Mycotoxins: Chemistry Genetics and Biology; American Phytopathological Society Press: St. Paul, MN, USA, 2006. [Google Scholar]
- Pitt, J.I.; Hocking, A.D. Current mycotoxin issues in Australia and Southeast Asia. Microbiol. Aust. 2003, 24, 4–6. [Google Scholar] [CrossRef]
- Bryden, W.L. Mycotoxins and mycotoxicoses: Significance, occurrence and mitigation in the food chain. In General and Applied Toxicology, 3rd ed.; Ballantyne, B., Marrs, T., Syversen, T., Eds.; John Wiley & Sons Ltd.: Chichester, UK, 2009; pp. 3529–3553. [Google Scholar]
- Nelson, P.E.; Dignani, M.C.; Anaisse, E.J. Taxonomy, biology, and clinical aspects of Fusarium species. Clin. Microbiol. Rev. 1994, 7, 479–504. [Google Scholar]
- Gordon, T.R. Fusarium oxysporum and the Fusarium Wilt Syndrome. Annu. Rev. Phytopathol. 2017, 55, 23–39. [Google Scholar] [CrossRef]
- Smith, S.N. An overview of ecological and habitat aspects in the genus Fusarium with special emphasis on the soil-borne pathogenic forms. Plant Pathol. Bull. 2007, 16, 97–120. [Google Scholar]
- McGovern, R.J. Management of tomato diseases caused by Fusarium oxysporum. Crop Prot. 2015, 73, 78–92. [Google Scholar] [CrossRef]
- Baysal, Ö.; Siragusa, M.; Gümrükcü, E.; Zengin, S.; Carimi, F.; Sajeva, M.; da Silva, J.A.T. Molecular characterization of Fusarium oxysporum f. melongenae by ISSR and RAPD markers on eggplant. Biochem. Genet. 2010, 48, 524–537. [Google Scholar] [CrossRef] [PubMed]
- Shabani, F.; Kumar, L.; Esmaeili, A. Future distributions of Fusarium oxysporum f. spp. in European, Middle Eastern and North African agricultural regions under climate change. Agric. Ecosyst. Environ. 2014, 197, 96–105. [Google Scholar] [CrossRef]
- McMahon, S.M.; Harrison, S.; Armbruster, W.S.; Bartlein, P.; Beale, C.M.; Edwards, M.E.; Kattge, J.; Midgley, G.; Morin, X.; Prentice, I.C. Improving assessment and modelling of climate change impacts on global terrestrial biodiversity. Trends Ecol. Evol. 2011, 26, 249–259. [Google Scholar] [CrossRef]
- Parmesan, C.; Duarte, C.M.; Poloczanska, E.; Richardson, A.J.; Singer, M.C. Overstretching attribution. Nat. Clim. Chang. 2011, 1, 2–4. [Google Scholar]
- Runquist, R.D.B.; Lake, T.; Tiffin, P.; Moeller, D.A. Species distribution models throughout the invasion history of Palmer amaranth predict regions at risk of future invasion and reveal challenges with modeling rapidly shifting geographic ranges. Sci. Rep. 2019, 9, 2426. [Google Scholar]
- Wilson, C.D.; Roberts, D.; Reid, N. Applying species distribution modelling to identify areas of high conservation value for endangered species: A case study using Margaritifera margaritifera (L.). Biol. Conserv. 2011, 144, 821–829. [Google Scholar] [CrossRef]
- Guo, S.; Ge, X.; Zou, Y.; Zhou, Y.; Wang, T.; Zong, S. Projecting the Potential Global Distribution of Carpomya vesuviana (Diptera: Tephritidae), Considering Climate Change and Irrigation Patterns. Forests 2019, 10, 355. [Google Scholar]
- Byeon, D.H.; Jung, J.M.; Jung, S.; Lee, W.H. Prediction of global geographic distribution of Metcalfa pruinosa using CLIMEX. Entomol. Res. 2018, 48, 99–107. [Google Scholar]
- Phillips, S.J.; Dudík, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Zurell, D.; Franklin, J.; König, C.; Bouchet, P.J.; Dormann, C.F.; Elith, J.; Fandos, G.; Feng, X.; Guillera-Arroita, G.; Guisan, A.; et al. A standard protocol for reporting species distribution models. Ecography 2020, 43, 1261–1277. [Google Scholar]
- Alkhalifah, D.H.M.; Damra, E.; Khalaf, S.M.H.; Hozzein, W.N. Biogeography of Black Mold Aspergillus niger: Global Situation and Future Perspective under Several Climate Change Scenarios Using MaxEnt Modeling. Diversity 2022, 14, 845. [Google Scholar] [CrossRef]
- Hosni, E.M.; Nasser, M.; Al-Ashaal, S.; Rady, M.H.; Kenawy, M.A. Modeling current and future global distribution of Chrysomya bezziana under changing climate. Sci. Rep. 2020, 10, 4947. [Google Scholar] [CrossRef] [PubMed]
- Abou-Shaara, H.; Alashaal, S.A.; Hosni, E.M.; Nasser, M.G.; Ansari, M.J.; Alharbi, S.A. Modeling the Invasion of the Large Hive Beetle, Oplostomus fuligineus, into North Africa and South Europe under a Changing Climate. Insects 2021, 12, 275. [Google Scholar] [CrossRef] [PubMed]
- Escobar, L.E.; Lira-Noriega, A.; Medina-Vogel, G.; Peterson, A.T. Potential for spread of the white-nose fungus (Pseudogymnoascus destructans) in the Americas: Use of Maxent and Niche A to assure strict model transference. Geospat. Health 2014, 9, 221–229. [Google Scholar]
- Hosni, E.M.; Nasser, M.; Al-Khalaf, A.A.; Al-Shammery, K.A.; Al-Ashaal, S.; Soliman, D. Invasion of the Land of Samurai: Potential Spread of Old-World Screwworm to Japan under Climate Change. Diversity 2022, 14, 99. [Google Scholar]
- Hosni, E.M.; Al-Khalaf, A.A.; Nasser, M.G.; Abou-Shaara, H.F.; Radwan, M.H. Modeling the Potential Global Distribution of Honeybee Pest, Galleria mellonella under Changing Climate. Insects 2022, 13, 484. [Google Scholar]
- Hosni, E.M.; Al-Khalaf, A.A.; Naguib, R.M.; Afify, A.E.; Abdalgawad, A.A.; Faltas, E.M.; Hassan, M.A.; Mahmoud, M.A.; Naeem, O.M.; Hassan, Y.M.; et al. Evaluation of Climate Change Impacts on the Global Distribution of the Calliphorid Fly Chrysomya albiceps Using GIS. Diversity 2022, 14, 578. [Google Scholar]
- Kessler, W.H.; Ganser, C.; Glass, G.E. Modeling the Distribution of Medically Important Tick Species in Florida. Insects 2019, 10, 190. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar]
- Elith, J.H.; Graham, C.P.H.; Anderson, R.P.; Dudík, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A.; et al. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar]
- Mulieri, P.R.; Patitucci, L.D. Using ecological niche models to describe the geographical distribution of the myiasis-causing Cochliomyia hominivorax (Diptera: Calliphoridae) in southern South America. Parasitol. Res. 2019, 118, 1077–1086. [Google Scholar] [CrossRef] [PubMed]
- Soliman, M.M.; Al-Khalaf, A.A.; El-Hawagry, M.S.A. Effects of Climatic Change on Potential Distribution of Spogostylum ocyale (Diptera: Bombyliidae) in the Middle East Using Maxent Modelling. Insects 2023, 14, 120. [Google Scholar] [CrossRef]
- Tyagi, S.; Singh, R.; Javeria, S. Effect of Climate Change on Plant-Microbe Interaction: An Overview. Eur. J. Mol. Biotechnol. 2014, 5, 149–156. Available online: https://goo.gl/tvo1AC (accessed on 13 December 2022).
- Weiman, S. Microbes help to drive global carbon cycling and climate change. Microbe Mag. 2015, 10, 233–238. [Google Scholar]
- Castro, H.F.; Classen, A.T.; Austin, E.E.; Norby, R.J.; Schadt, C.W. Soil microbial community responses to multiple experimental climate change drivers. Appl. Environ. Microbiol. 2010, 76, 999–1007. [Google Scholar] [CrossRef]
- Allison, S.D.; Wallenstein, M.D.; Bradford, M.A. Soil carbon response to warming dependent on microbial physiology. Nat. Geosci. 2010, 3, 336–340. [Google Scholar]
- Sayer, E.J.; Oliver, A.E.; Fridley, J.D.; Askew, A.P.; Mills, R.T.E.; Grime, J.P. Links between soil microbial communities and plant traits in a species-rich grassland under long-term climate change. Ecol. Evolution. 2017, 7, 855–862. [Google Scholar]
- Vicuña, R.; González, B. The microbial world in a changing environment. Rev. Chil. Hist. Nat. 2021, 94, 2. [Google Scholar] [CrossRef]
- Gordon, T.R.; Okamoto, D.; Milgroom, M.G. The structure and interrelationship of fungal populations in native and cultivated soils. Mol. Ecol. 1992, 1, 241–249. [Google Scholar]
- Ganley, R.J.; Watt, M.S.; Kriticos, D.J.; Hopkins, A.J.M.; Manning, L.K. Increased risk of pitch canker to Australasia under climate change. Australas. Plant Pathol. 2011, 40, 228–237. [Google Scholar]
- Magan, N.; Medina-Vaya, A.; Aldred, D. Possible climate-change effects on mycotoxin contamination of food crops pre- and postharvest. Plant Pathol. 2011, 60, 150–163. [Google Scholar] [CrossRef]
- Matthews, M.C.; Mostert, D.; Ndayihanzamaso, P.; Rose, L.J.; Viljoen, A. Quantitative detection of economically important Fusarium oxysporum f. sp. cubense strains in Africa in plants, soil and water. PLoS ONE 2020, 15, e0236110. [Google Scholar]
- FAOSTAT. Bananamarket Review: Preliminary Results for 2017; Food and Agriculture Organizations of United Nations: Rome, Italy, 2017; Available online: http://www.fao.org/fileadmin/templates/est/COMM_MARKETS_MONITORING/Bananas/Documents/Banana_Market_Review_December_2017_update.pdf (accessed on 13 December 2022).
- Scheerer, L.; Pemsl, D.E.; Dita, M.; Vicente, L.P.; Staver, C. A quantified approach to projecting losses caused by Fusarium wilt tropical race4. Acta Hortic. 2018, 1196, 211–218. [Google Scholar]
- Tol, R.S.J.; Downing, T.E.; Kuik, O.J.; Smith, J.B. Distributional aspects of climate change impacts. Glob. Environ. Chang. 2004, 14, 259–272. [Google Scholar] [CrossRef]
- Callaway, J. Adaptation benefits and costs: Are they important in the global policy picture and how can we estimate them? Glob. Environ. Chang. 2004, 14, 275–284. [Google Scholar]
- Parry, M.; Arnell, N.; McMichael, T.; Nicholls, R.; Martens, P.; Kovats, S.; Livermore, M.; Rosenzweig, C.; Iglesias, A.; Fischer, G. Millions at risk: Defining critical climate change threats and targets. Glob. Environ. Chang. 2001, 11, 181–183. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Description |
|---|---|
| Bio 1 | Annual mean temperature |
| Bio 2 | Mean diurnal range (mean of monthly max temp–min temp) |
| Bio 3 | Isothermality (bio2/bio7 ) × 100 |
| Bio 4 | Temperature seasonality (standard deviation × 100) |
| Bio 5 | Max temperature of warmest month |
| Bio 6 | Min temperature of coldest month |
| Bio 7 | Temperature annual range |
| Bio 8 | Mean temperature of wettest quarter |
| Bio 9 | Mean temperature of driest quarter |
| Bio 10 | Mean temperature of warmest quarter |
| Bio 11 | Mean temperature of coldest quarter |
| Bio 12 | Annual precipitation |
| Bio 13 | Precipitation of wettest month |
| Bio 14 | Precipitation of driest month |
| Bio 15 | Precipitation seasonality (coefficient of variation) |
| Bio 16 | Precipitation of wettest quarter |
| Bio 17 | Precipitation of driest quarter |
| Bio 18 | Precipitation of warmest quarter |
| Bio 19 | Precipitation of coldest quarter |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alkhalifah, D.H.M.; Damra, E.; Melhem, M.B.; Hozzein, W.N. Fungus under a Changing Climate: Modeling the Current and Future Global Distribution of Fusarium oxysporum Using Geographical Information System Data. Microorganisms 2023, 11, 468. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms11020468
Alkhalifah DHM, Damra E, Melhem MB, Hozzein WN. Fungus under a Changing Climate: Modeling the Current and Future Global Distribution of Fusarium oxysporum Using Geographical Information System Data. Microorganisms. 2023; 11(2):468. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms11020468
Chicago/Turabian StyleAlkhalifah, Dalal Hussien M., Eman Damra, Moaz Beni Melhem, and Wael N. Hozzein. 2023. "Fungus under a Changing Climate: Modeling the Current and Future Global Distribution of Fusarium oxysporum Using Geographical Information System Data" Microorganisms 11, no. 2: 468. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms11020468