
Resistance to the Bacteriocin Lcn972 Deciphered by Genome Sequencing
 ,
,
Abstract
:1. Introduction
2. Material and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. Library Preparation and Sequencing
2.3. RNA Extraction and Quantitative Reverse Transcription-PCR (RT-qPCR)
2.4. Evolution Experiments
2.5. Phenotypic Testing during Evolution and Stabilization
2.6. PCR Detection of IS905::celB and IS981::LLMG_RS12285
2.7. MIC Determinations
2.8. Microscopy
3. Results and Discussion
3.1. Genome Analysis of L. lactis MG1614 Reveals Laboratory-Selected Mutations
3.2. Genome Analysis of the Lcn972R Mutant L. lactis D1-20
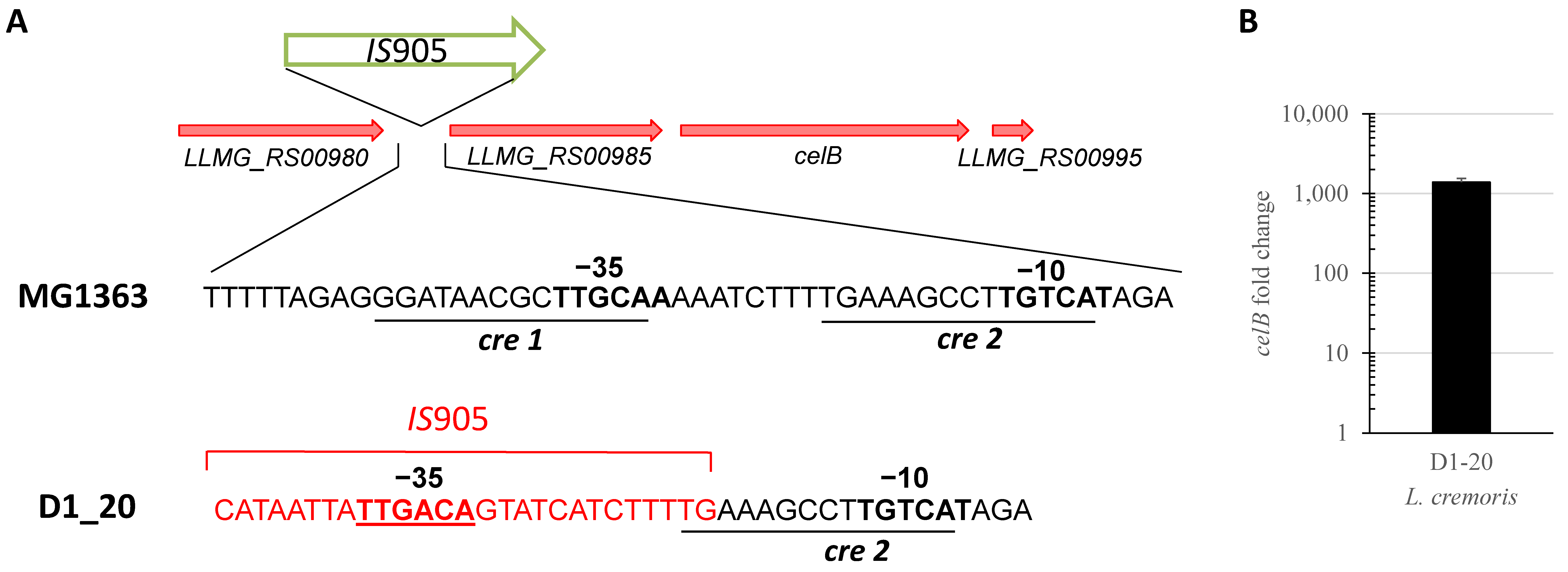
3.3. Cellobiose Uptake Is Activated in L. cremoris D1-20
3.4. Transposition of IS981 into the LLMG_RS12285 Promoter Is Selected Early during Evolution in the Presence of Lcn972 and Is Not Lost upon Successive Cultivation without Selective Pressure
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. 2021 Antibacterial Agents in Clinical and Preclinical Development: An Overview and Analysis; World Health Organization: Geneva, Switzerland, 2022; ISBN 978-92-4-004765-5. ISSN 978-92-4-004766-2. [Google Scholar]
- Cebrián, R.; Rodríguez-Cabezas, M.E.; Martín-Escolano, R.; Rubiño, S.; Garrido-Barros, M.; Montalbán-López, M.; Rosales, M.J.; Sánchez-Moreno, M.; Valdivia, E.; Martínez-Bueno, M.; et al. Preclinical Studies of Toxicity and Safety of the AS-48 Bacteriocin. J. Adv. Res. 2019, 20, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Martín, R.; Escobedo, S.; Martín, C.; Crespo, A.; Quiros, L.M.; Suarez, J.E. Surface glycosaminoglycans protect eukaryotic cells against membrane-driven peptide bacteriocins. Antimicrob. Agents Chemother. 2015, 59, 677–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soltani, S.; Boutin, Y.; Couture, F.; Biron, E.; Subirade, M.; Fliss, I. In vitro assessment of skin sensitization, irritability and toxicity of bacteriocins and reuterin for possible topical applications. Sci. Rep. 2022, 12, 4570. [Google Scholar] [CrossRef] [PubMed]
- Benítez-Chao, D.F.; León-Buitimea, A.; Lerma-Escalera, J.A.; Morones-Ramírez, J.R. Bacteriocins: An Overview of Antimicrobial, Toxicity, and Biosafety Assessment by in vivo Models. Front. Microbiol. 2021, 12, 677. [Google Scholar] [CrossRef]
- Heilbronner, S.; Krismer, B.; Brötz-Oesterhelt, H.; Peschel, A. The microbiome-shaping roles of bacteriocins. Nat. Rev. Microbiol. 2021, 19, 726–739. [Google Scholar] [CrossRef]
- Rea, M.C.; Sit, C.S.; Clayton, E.; O’Connor, P.M.; Whittal, R.M.; Zheng, J.; Vederas, J.C.; Ross, R.P.; Hill, C. Thuricin CD, a posttranslationally modified bacteriocin with a narrow spectrum of activity against Clostridium difficile. Proc. Natl. Acad. Sci. USA 2010, 107, 9352–9357. [Google Scholar] [CrossRef] [Green Version]
- Field, D.; Molloy, E.M.; Iancu, C.; Draper, L.A.; O’Connor, P.M.; Cotter, P.D.; Hill, C.; Ross, R.P. Saturation mutagenesis of selected residues of the alpha-peptide of the lantibiotic lacticin 3147 yields a derivative with enhanced antimicrobial activity. Microb. Biotechnol. 2013, 6, 564–575. [Google Scholar] [CrossRef]
- Li, Q.; Montalban-Lopez, M.; Kuipers, O.P. Increasing the antimicrobial activity of nisin-based lantibiotics against Gram-negative pathogens. Appl. Environ. Microbiol. 2018, 84, e00052-18. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Ramos, A.; Madi-Moussa, D.; Coucheney, F.; Drider, D. Current knowledge of the mode of action and immunity mechanisms of lab-bacteriocins. Microorganisms 2021, 9, 2107. [Google Scholar] [CrossRef]
- Acedo, J.Z.; Chiorean, S.; Vederas, J.C.; van Belkum, M.J. The expanding structural variety among bacteriocins from Gram-positive bacteria. FEMS Microbiol. Rev. 2018, 42, 805–828. [Google Scholar] [CrossRef]
- Campelo, A.B.; López-González, M.J.; Escobedo, S.; Janzen, T.; Neves, A.R.; Rodríguez, A.; Martínez, B. Mutations Selected After Exposure to Bacteriocin Lcn972 Activate a Bce-Like Bacitracin Resistance Module in Lactococcus lactis. Front. Microbiol. 2020, 11, 1805. [Google Scholar] [CrossRef] [PubMed]
- Tymoszewska, A.; Ovchinnikov, K.V.; Diep, D.B.; Słodownik, M.; Maron, E.; Martínez, B.; Aleksandrzak-Piekarczyk, T. Lactococcus lactis Resistance to Aureocin A53- and Enterocin L50-Like Bacteriocins and Membrane-Targeting Peptide Antibiotics Relies on the YsaCB-KinG-LlrG Four-Component System. Antimicrob. Agents Chemother. 2021, 65, e00921-21. [Google Scholar] [CrossRef] [PubMed]
- Katla, T.; Naterstad, K.; Vancanneyt, M.; Swings, J.; Axelsson, L. Differences in susceptibility of Listeria monocytogenes strains to sakacin P, sakacin A, pediocin PA-1, and nisin. Appl. Environ. Microbiol. 2003, 69, 4431–4437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bastos Mdo, C.; Coelho, M.L.; Santos, O.C. Resistance to bacteriocins produced by Gram-positive bacteria. Microbiology 2015, 161, 683–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Draper, L.A.; Cotter, P.D.; Hill, C.; Ross, R.P. Lantibiotic resistance. Microbiol. Mol. Biol. Rev. 2015, 79, 171–191. [Google Scholar] [CrossRef] [Green Version]
- Martínez, B.; Böttiger, T.; Schneider, T.; Rodríguez, A.; Sahl, H.G.; Wiedemann, I. Specific interaction of the unmodified bacteriocin Lactococcin 972 with the cell wall precursor lipid II. Appl. Environ. Microbiol. 2008, 74, 4666–4670. [Google Scholar] [CrossRef] [Green Version]
- Martínez, B.; Zomer, A.L.; Rodríguez, A.; Kok, J.; Kuipers, O.P. Cell envelope stress induced by the bacteriocin Lcn972 is sensed by the lactococcal two-component system CesSR. Mol. Microbiol. 2007, 64, 473–486. [Google Scholar] [CrossRef] [Green Version]
- Turner, D.L.; Lamosa, P.; Rodríguez, A.; Martínez, B. Structure and properties of the metastable bacteriocin Lcn972 from Lactococcus lactis. J. Mol. Struct. 2013, 1031, 207–210. [Google Scholar] [CrossRef]
- Rolhion, N.; Chassaing, B.; Nahori, M.-A.; de Bodt, J.; Moura, A.; Lecuit, M.; Dussurget, O.; Bérard, M.; Marzorati, M.; Fehlner-Peach, H.; et al. Listeria monocytogenes Bacteriocin Can Target the Commensal Prevotella copri and Modulate Intestinal Infection. Cell Host Microbe 2019, 26, 691–701.e5. [Google Scholar] [CrossRef] [Green Version]
- Roces, C.; Courtin, P.; Kulakauskas, S.; Rodríguez, A.; Chapot-Chartier, M.P.; Martínez, B. Isolation of Lactococcus lactis mutants simultaneously resistant to the cell wall-active bacteriocin Lcn972, lysozyme, nisin and bacteriophage c2. Appl. Environ. Microbiol. 2012, 78, 4157–4163. [Google Scholar] [CrossRef] [Green Version]
- López-González, M.J.; Escobedo, S.; Rodríguez, A.; Neves, A.R.; Janzen, T.; Martínez, B. Adaptive Evolution of Industrial Lactococcus lactis under Cell Envelope Stress Provides Phenotypic Diversity. Front. Microbiol. 2018, 9, 2654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roces, C.; Pérez, V.; Campelo, A.B.; Blanco, D.; Kok, J.; Kuipers, O.P.; Rodríguez, A.; Martínez, B. The putative lactococcal extracytoplasmic function anti-sigma factor llmg2447 determines resistance to the cell wall-active bacteriocin lcn972. Antimicrob. Agents Chemother. 2012, 56, 5520–5527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gasson, M.J. Plasmid complements of Streptococcus lactis NCDO 712 and other lactic streptococci after protoplast-induced curing. J. Bacteriol. 1983, 154, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. 2010. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc (accessed on 21 March 2022).
- Nurk, S.; Bankevich, A.; Antipov, D.; Gurevich, A.; Korobeynikov, A.; Lapidus, A.; Prjibelsky, A.; Pyshkin, A.; Sirotkin, A.; Sirotkin, Y.; et al. Assembling Genomes and Mini-metagenomes from Highly Chimeric Reads. In Research in Computational Molecular Biology; RECOMB 2013. Lecture Notes in Computer Science; Deng, M., Jiang, R., Sun, F., Zhang, X., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; Volume 7821. [Google Scholar] [CrossRef]
- Galardini, M.; Biondi, E.G.; Bazzicalupo, M.; Mengoni, A. CONTIGuator: A bacterial genomes finishing tool for structural insights on draft genomes. Source Code Biol. Med. 2011, 6, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darling, A.C.; Mau, B.; Blattner, F.R.; Perna, N.T. Mauve: Multiple alignment of conserved genomic sequence with rearrangements. Genome Res. 2004, 14, 1394–1403. [Google Scholar] [CrossRef] [Green Version]
- Deatherage, D.E.; Barrick, J.E. Identification of mutations in laboratory-evolved microbes from next-generation sequencing data using breseq. Methods Mol. Biol. 2014, 1151, 165–188. [Google Scholar]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Madera, C.; Monjardín, C.; Suárez, J.E. Milk contamination and resistance to processing conditions determine the fate of Lactococcus lactis bacteriophages in dairies. Appl. Environ. Microbiol. 2004, 70, 7365–7371. [Google Scholar] [CrossRef] [Green Version]
- Martínez, B.; Suárez, J.E.; Rodríguez, A. Lactococcin 972, a homodimeric lactococcal bacteriocin whose primary target is not the plasma membrane. Microbiology 1996, 142, 2393–2398. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Zhou, Y.N.; Goldstein, B.P.; Jin, D.J. Cross-resistance of Escherichia coli RNA polymerases conferring rifampin resistance to different antibiotics. J. Bacteriol. 2005, 187, 2783–2792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wittmann, H.G.; Apirion, D. Analysis of ribosomal proteins in streptomycin resistant and dependent mutants isolated from streptomycin independent Escherichia coli strains. Mol. Gen. Genet. 1975, 141, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Linares, D.M.; Kok, J.; Poolman, B. Genome sequences of Lactococcus lactis MG1363 (revised) and NZ9000 and comparative physiological studies. J. Bacteriol. 2010, 192, 5806–5812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solopova, A.; Kok, J.; Kuipers, O.P. Disruption of a transcriptional repressor by an insertion sequence element integration leads to activation of a novel silent cellobiose transporter in Lactococcus lactis MG1363. Appl. Environ. Microbiol. 2017, 83, e01279-17. [Google Scholar] [CrossRef] [Green Version]
- Gebhard, S. ABC transporters of antimicrobial peptides in Firmicutes bacteria—Phylogeny, function and regulation. Mol. Microbiol. 2012, 86, 1295–1317. [Google Scholar] [CrossRef]
- Consuegra, J.; Gaffé, J.; Lenski, R.E.; Hindré, T.; Barrick, J.E.; Tenaillon, O.; Schneider, D. Insertion-sequence-mediated mutations both promote and constrain evolvability during a long-term experiment with bacteria. Nat. Commun. 2021, 12, 980. [Google Scholar] [CrossRef]
- de Visser, J.A.; Akkermans, A.D.; Hoekstra, R.F.; de Vos, W.M. Insertion-sequence-mediated mutations isolated during adaptation to growth and starvation in Lactococcus lactis. Genetics 2004, 168, 1145–1157. [Google Scholar] [CrossRef] [Green Version]
- Aleksandrzak-Piekarczyk, T.; Polak, J.; Jezierska, B.; Renault, P.; Bardowski, J. Genetic characterization of the CcpA-dependent, cellobiose-specific PTS system comprising CelB, PtcB and PtcA that transports lactose in Lactococcus lactis IL1403. Int. J. Food Microbiol. 2011, 145, 186–194. [Google Scholar] [CrossRef]
- Campelo, A.B.; Gaspar, P.; Roces, C.; Rodríguez, A.; Kok, J.; Kuipers, O.P.; Neves, A.R.; Martínez, B. The Lcn972 bacteriocin-encoding plasmid pBL1 impairs cellobiose metabolism in Lactococcus lactis. Appl. Environ. Microbiol. 2011, 77, 7576–7585. [Google Scholar] [CrossRef] [Green Version]
- Solopova, A.; Bachmann, H.; Teusink, B.; Kok, J.; Neves, A.R.; Kuipers, O.P. A specific mutation in the promoter region of the silent cel cluster accounts for the appearance of lactose-utilizing Lactococcus lactis MG1363. Appl. Environ. Microbiol. 2012, 78, 5612–5621. [Google Scholar] [CrossRef] [Green Version]
- Chapot-Chartier, M.P.; Vinogradov, E.; Sadovskaya, I.; Andre, G.; Mistou, M.Y.; Trieu-Cuot, P.; Furlan, S.; Bidnenko, E.; Courtin, P.; Péchoux, C.; et al. Cell surface of Lactococcus lactis is covered by a protective polysaccharide pellicle. J. Biol. Chem. 2010, 285, 10464–10471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahony, J.; Randazzo, W.; Neve, H.; Settanni, L.; van Sinderen, D. Lactococcal 949 group phages recognize a carbohydrate receptor on the host cell surface. Appl. Environ. Microbiol. 2015, 81, 3299–3305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahony, J.; Oliveira, J.; Collins, B.; Hanemaaijer, L.; Lugli, G.A.; Neve, H.; Ventura, M.; Kouwen, T.R.; Cambillau, C.; van Sinderen, D. Genetic and functional characterisation of the lactococcal P335 phage-host interactions. BMC Genom. 2017, 18, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guinane, C.M.; Cotter, P.D.; Hill, C.; Ross, R.P. Spontaneous resistance in Lactococcus lactis IL1403 to the Lantibiotic lacticin 3147. FEMS Microbiol. Lett. 2006, 260, 77–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kramer, N.E.; Hasper, H.E.; van den Bogaard, P.T.; Morath, S.; de Kruijff, B.; Hartung, T.; Smid, E.J.; Breukink, E.; Kok, J.; Kuipers, O.P. Increased D-alanylation of lipoteichoic acid and a thickened septum are main determinants in the nisin resistance mechanism of Lactococcus lactis. Microbiology 2008, 154, 1755–1762. [Google Scholar] [CrossRef] [Green Version]
- Tymoszewska, A.; Aleksandrzak-Piekarczyk, T. The Lactococcal dgkB (yecE) and dxsA Genes for Lipid Metabolism Are Involved in the Resistance to Cell Envelope-Acting Antimicrobials. Int. J. Mol. Sci. 2021, 22, 1014. [Google Scholar] [CrossRef]
- McDonald, M.J. Microbial Experimental Evolution—A proving ground for evolutionary theory and a tool for discovery. EMBO Rep. 2019, 20, e46992. [Google Scholar] [CrossRef]
- Remigi, P.; Masson-Boivin, C.; Rocha, E. Experimental Evolution as a Tool to Investigate Natural Processes and Molecular Functions. Trends Microbiol. 2019, 27, 623–634. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Position | Change | Annotation | Locus_tag | Description | MG1614 | D1_20 |
|---|---|---|---|---|---|---|
| 218,269 | +C | coding (1042/1056 nt) | LLMG_RS01195→ | DUF2142 domain-containing protein | * | * |
| 269,158 | C→A | L633I (CTT→ATT) | LLMG_RS01465→ | Anaerobic ribonucleoside-triphosphate reductase | - | * |
| 446,872 | C→A | intergenic (−46/−65) | LLMG_RS02295←/→LLMG_RS02300 | trehalose operon repressor/PTS sugar transporter subunit IIA | * | - |
| 482,337 | C→T | H723Y (CAT→TAT) | LLMG_RS02480→ | glycoside hydrolase family 65 protein | * | * |
| 636,592 | (A)7→6 | coding (102/969 nt) | LLMG_RS03345← | ABC transporter permease | * | * |
| 643,970 | T→G | T7P (ACT→CCT) | LLMG_RS03375← | IS3 family transposase | * | * |
| 894,627 | +G | pseudogene (1075/1176 nt) | LLMG_RS04715→ | cell surface protein | * | * |
| 945,239 | +C | coding (307/330 nt) | LLMG_RS04990→ | hypothetical protein | * | * |
| 1,093,299 | +C | coding (3979/4050 nt) | LLMG_RS13005→ | cell wall anchor | * | * |
| 1,093,562 | +C | pseudogene (175/839 nt) | LLMG_RS13155→ | hypothetical protein | * | * |
| 1,134,220 | C→A | T147K (ACA→AAA) | LLMG_RS05920→ | alpha-glucuronidase | * | * |
| 1,210,283 | (A)5→6 | pseudogene (515/1046 nt) | LLMG_RS06265← | LacI family transcriptional regulator | * | * |
| 1,223,557 | 2 bp→AG | coding (236-237/237 nt) | LLMG_RS13200→ | hypothetical protein | * | * |
| 1,318,548 | (A)7→6 | pseudogene (98/412 nt) | LLMG_RS06810→ | hypothetical protein | * | * |
| 1,612,853 | C→A | intergenic (-60/+63) | LLMG_RS08235←/←LLMG_RS08240 | ABC transporter ATP-binding protein/EamA family transporter | - | * |
| 1,614,130 | +T | coding (1149/1278 nt) | LLMG_RS08245← | citrate:sodium symporter | * | * |
| 1,660,056 | (T)7→8 | intergenic (-14/+100) | LLMG_RS08475←/←LLMG_RS08480 | hypothetical protein/DNA repair protein RecN | * | * |
| 1,826,465 | (T)6→7 | intergenic (-23/+300) | LLMG_RS09240←/←LLMG_RS09245 | metal-dependent hydrolase/cold-shock protein | * | * |
| 1,968,674 | G→A | S491F (TCT→TTT) | LLMG_RS09935← | DNA-directed RNA polymerase subunit beta | * | * |
| 2,112,674 | +T | pseudogene (396/1083 nt) | LLMG_RS10765← | hypothetical protein | * | * |
| 2,159,055 | G→A | R110 * (CGA→TGA) | LLMG_RS11020← | aspartate protease | - | * |
| 2,183,058 | (A)5→4 | coding (399/1161 nt) | LLMG_RS11130← | helix-turn-helix domain-containing protein | * | * |
| 2,191,553 | G→A | A94T (GCT→ACT) | LLMG_RS11180→ | DNA replication and repair protein RecF | * | * |
| 2,266,855 | C→T | R54W (CGG→TGG) | LLMG_RS11605→ | arginine repressor | * | * |
| 2,517,498 | T→C | K56R (AAA→AGA) | LLMG_RS12900← | 30S ribosomal protein S12 | * | * |
| Sample | Lcn972 (AU/mL) | Log CFU/mL | n 1 | Growth on Maltose 2 | Growth in Lcn972 (80 AU/mL) 2 | IS905 Insertion 2 | IS981 Insertion 2 |
|---|---|---|---|---|---|---|---|
| MA | 0 | 8.85 | 31 | 31 | 0 | 0 | 0 |
| MB | 0 | 8.91 | 31 | 31 | 0 | 0 | 0 |
| T4A | 80 | 8.93 | 31 | 31 | 28 | 0 | 31 |
| T4B | 80 | 8.30 | 31 | 31 | 31 | 0 | 31 |
| T7A | 1280 | 8.42 | 31 | 31 (31) | 31 (31) | 0 | 31 |
| T7B | 1280 | 7.86 | 31 | 31 (26) | 31 (26) | 0 (0) | 31 (26) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Escobedo, S.; Campelo, A.B.; Umu, Ö.C.O.; López-González, M.J.; Rodríguez, A.; Diep, D.B.; Martínez, B. Resistance to the Bacteriocin Lcn972 Deciphered by Genome Sequencing. Microorganisms 2023, 11, 501. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms11020501
Escobedo S, Campelo AB, Umu ÖCO, López-González MJ, Rodríguez A, Diep DB, Martínez B. Resistance to the Bacteriocin Lcn972 Deciphered by Genome Sequencing. Microorganisms. 2023; 11(2):501. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms11020501
Chicago/Turabian StyleEscobedo, Susana, Ana B. Campelo, Özgün C. O. Umu, María Jesús López-González, Ana Rodríguez, Dzung B. Diep, and Beatriz Martínez. 2023. "Resistance to the Bacteriocin Lcn972 Deciphered by Genome Sequencing" Microorganisms 11, no. 2: 501. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms11020501