
Antiviral Effect of Ephedrine Alkaloids-Free Ephedra Herb Extract against SARS-CoV-2 In Vitro
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.2. Preparation of EFE and EMCT
2.3. Virus Growth Inhibition Assay
2.4. TCID50 Assay
2.5. Cell Viability Assay
2.6. Statistical Analysis
3. Results
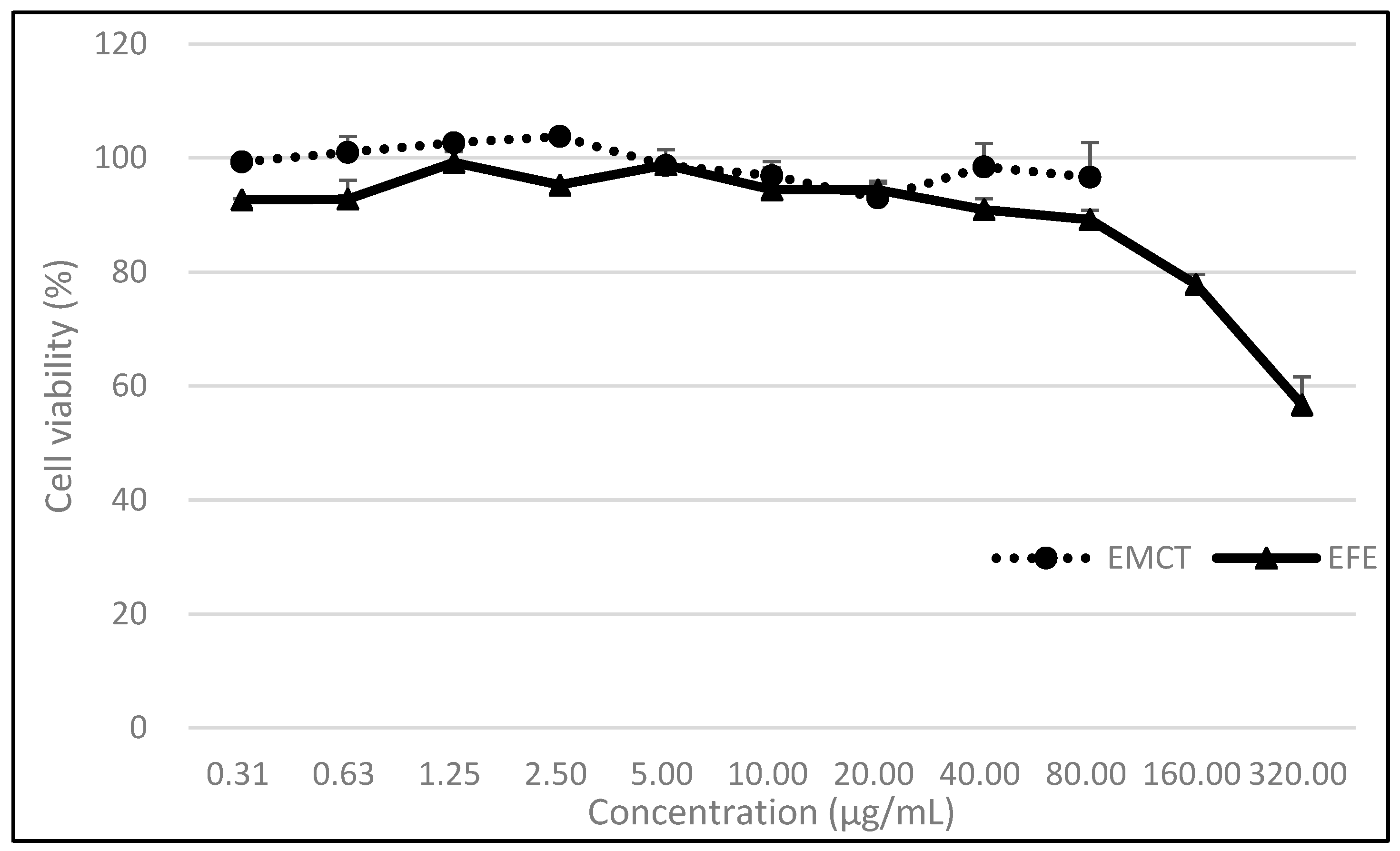
3.1. Cell Viability under EFE or EMCT Treatment
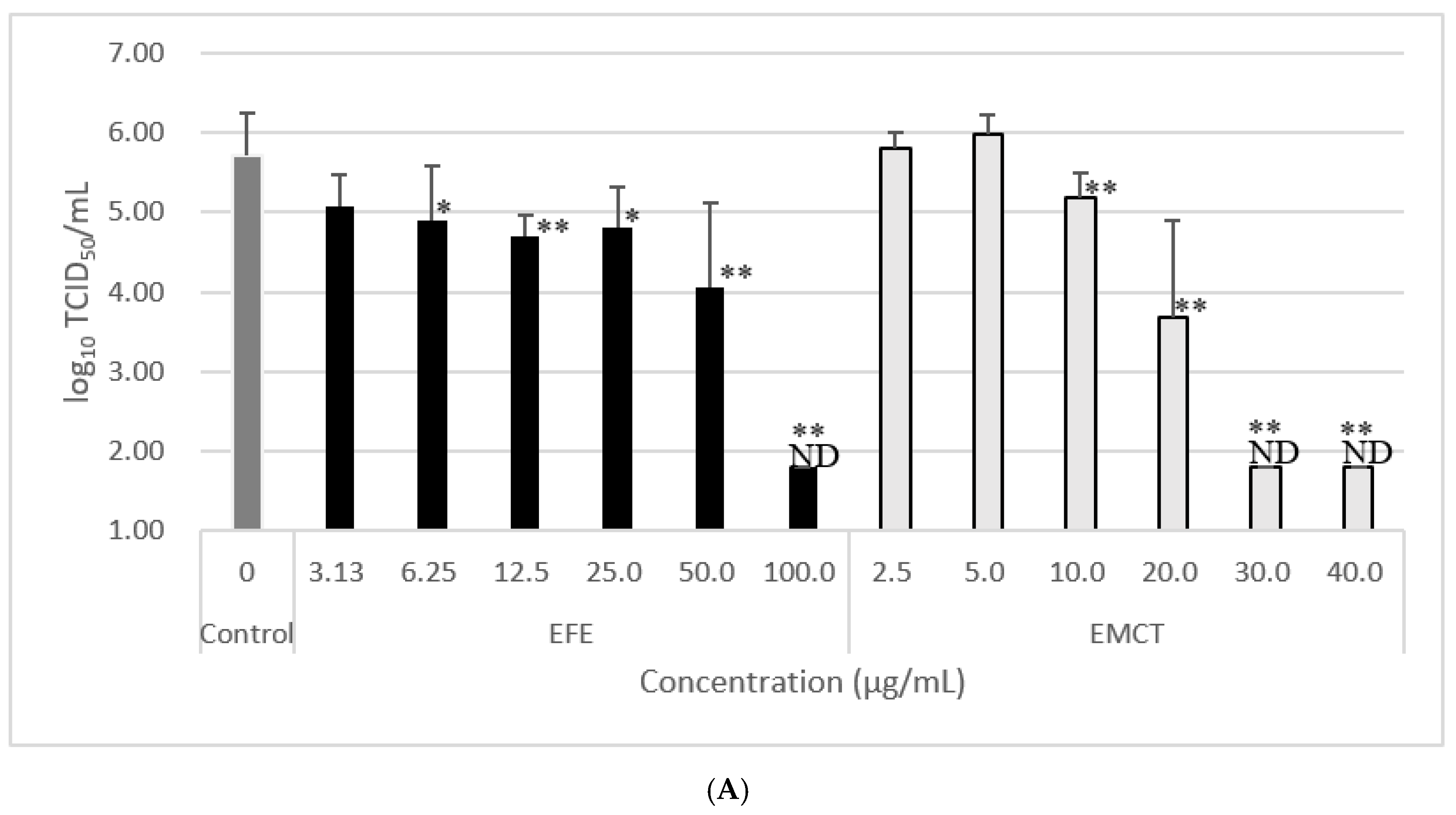
3.2. Virus Growth Inhibition of Pretreatment with EFE or EMCT
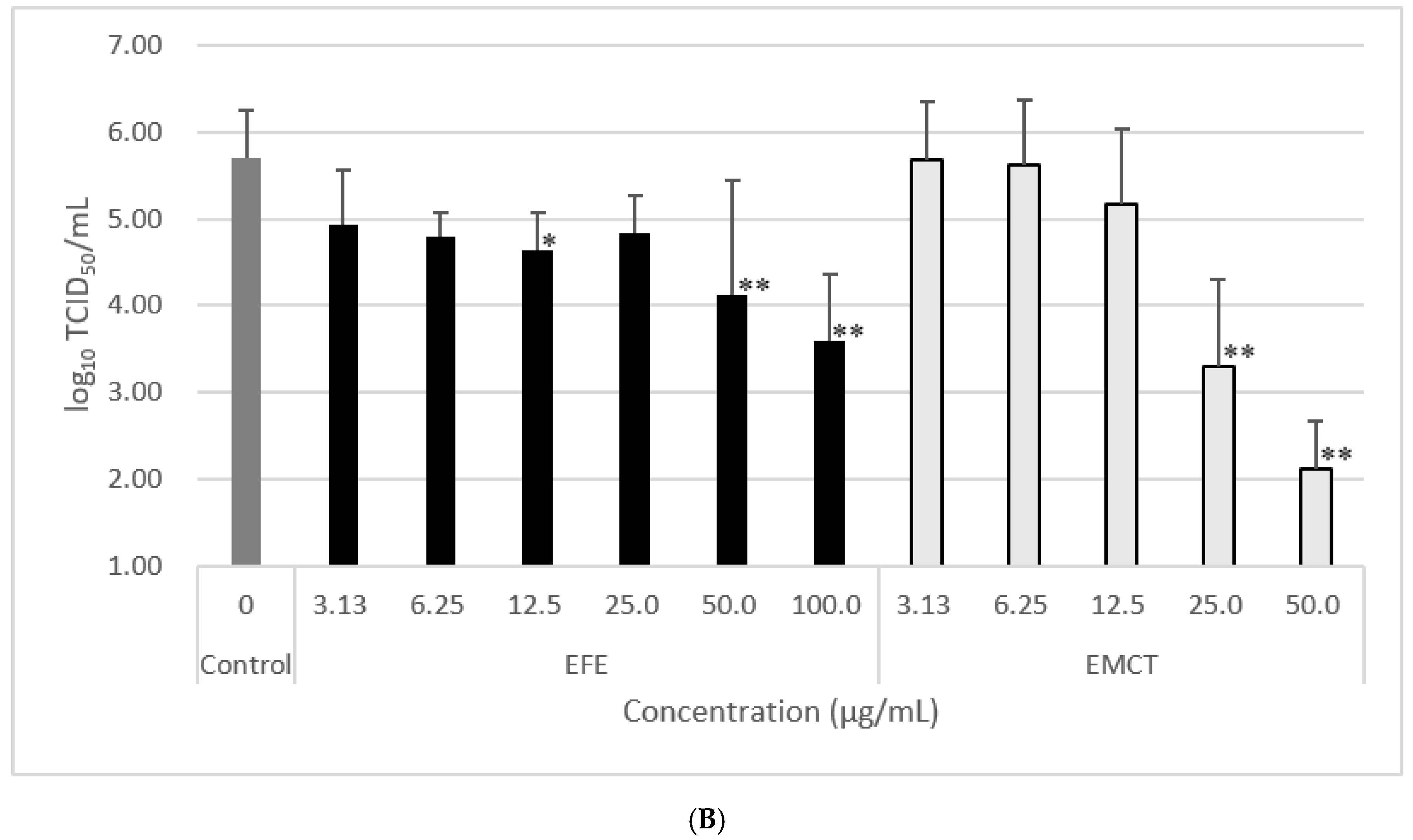
3.3. Virus Growth Inhibition of Post-Treatment with EFE or EMCT
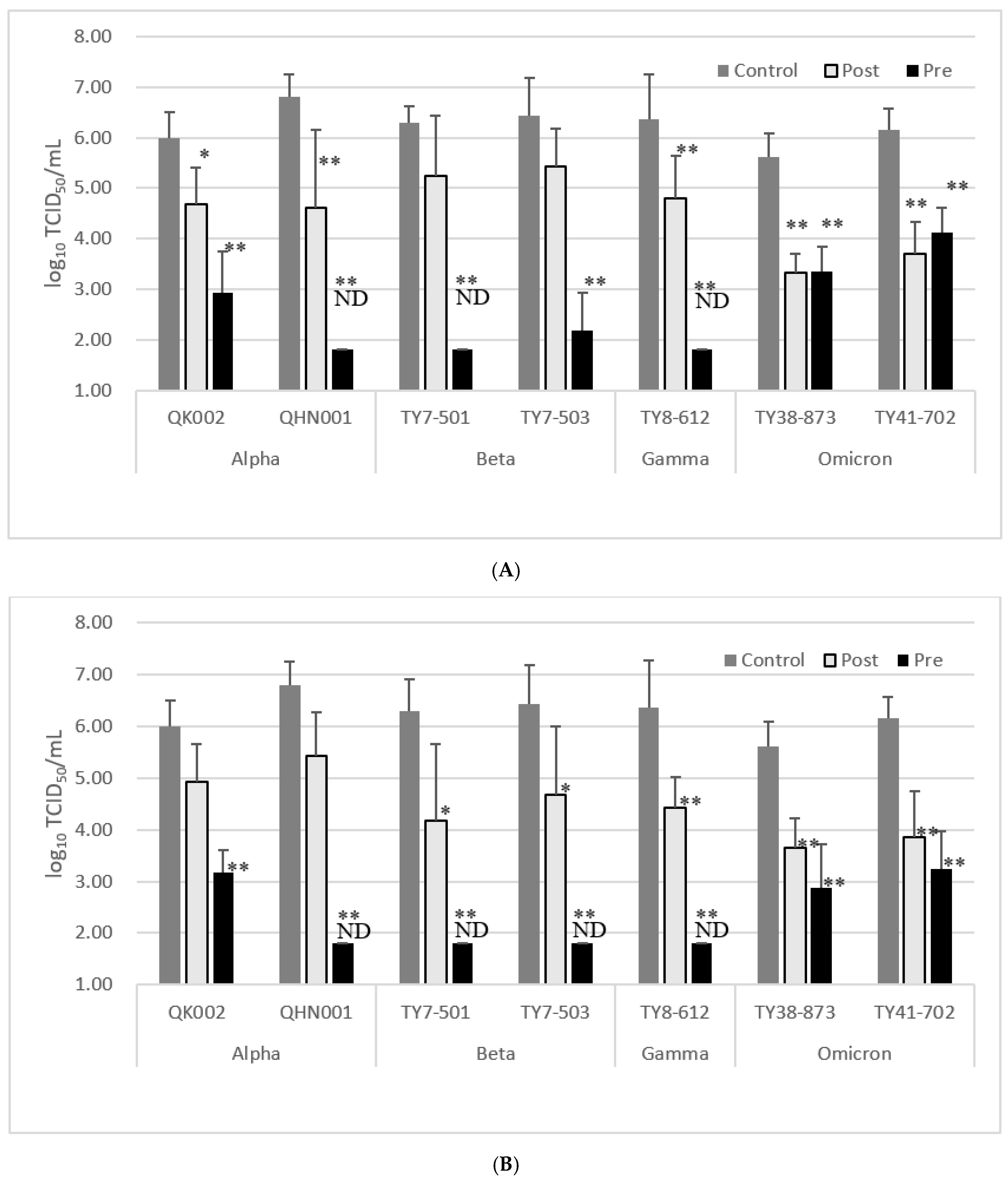
3.4. Inhibitory Effect of EFE or EMCT against Variant Strains
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. WHO Coronavirus (COVID-19) Dashboard. Available online: https://covid19.who.int/ (accessed on 30 December 2022).
- Ceramella, J.; Iacopetta, D.; Sinicropi, M.S.; Andreu, I.; Mariconda, A.; Saturnino, C.; Giuzio, F.; Longo, P.; Aquaro, S.; Catalano, A. Drugs for COVID-19: An Update. Molecules 2022, 27, 8562. [Google Scholar] [CrossRef] [PubMed]
- Anka, A.U.; Tahir, M.I.; Abubakar, S.D.; Alsabbagh, M.; Zian, Z.; Hamedifar, H.; Sabzevari, A.; Azizi, G. Coronavirus disease 2019 (COVID-19): An overview of the immunopathology, serological diagnosis and management. Scand. J. Immunol. 2020, 93, e12998. [Google Scholar] [CrossRef] [PubMed]
- Mantani, N.; Andoh, T.; Kawamata, H.; Terasawa, K.; Ochiai, H. Inhibitory effect of Ephedrae herba, an oriental traditional medicine, on the growth of influenza A/PR/8 virus in MDCK cells. Antivir. Res. 1999, 44, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Hyuga, S.; Hyuga, M.; Oshima, N.; Maruyama, T.; Kamakura, H.; Yamashita, T.; Yoshimura, M.; Amakura, Y.; Hakamatsuka, T.; Odaguchi, H.; et al. Ephedrine alkaloids-free Ephedra Herb extract: A safer alternative to ephedra with comparable analgesic, anticancer, and anti-influenza activities. J. Nat. Med. 2016, 70, 571–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takemoto, H.; Takahashi, J.; Hyuga, S.; Odaguchi, H.; Uchiyama, N.; Maruyama, T.; Yamashita, T.; Hyuga, M.; Oshima, N.; Amakura, Y.; et al. Ephedrine Alkaloids-Free Ephedra Herb Extract, EFE, Has No Adverse Effects Such as Excitation, Insomnia, and Arrhythmias. Biol. Pharm. Bull. 2018, 41, 247–253. [Google Scholar] [CrossRef] [Green Version]
- Odaguchi, H.; Sekine, M.; Hyuga, S.; Hanawa, T.; Hoshi, K.; Sasaki, Y.; Aso, M.; Yang, J.; Hyuga, M.; Kobayashi, Y.; et al. A Double-Blind, Randomized, Crossover Comparative Study for Evaluating the Clinical Safety of Ephedrine Alkaloids-Free Ephedra Herb Extract (EFE). Evid.-Based Complement. Altern. Med. 2018, 2018, 4625358. [Google Scholar] [CrossRef] [Green Version]
- Yoshimura, M.; Amakura, Y.; Hyuga, S.; Hyuga, M.; Nakamori, S.; Maruyama, T.; Oshima, N.; Uchiyama, N.; Yang, J.; Oka, H.; et al. Quality Evaluation and Characterization of Fractions with Biological Activity from Ephedra Herb Extract and Ephedrine Alkaloids-Free Ephedra Herb Extract. Chem. Pharm. Bull. 2020, 68, 140–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuyama, S.; Nao, N.; Shirato, K.; Kawase, M.; Saito, S.; Takayama, I.; Nagata, N.; Sekizuka, T.; Katoh, H.; Kato, F.; et al. Enhanced isolation of SARS-CoV-2 by TMPRSS2-expressing cells. Proc. Natl. Acad. Sci. USA 2020, 117, 7001–7003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oshima, N.; Yamashita, T.; Hyuga, S.; Hyuga, M.; Kamakura, H.; Yoshimura, M.; Maruyama, T.; Hakamatsuka, T.; Amakura, Y.; Hanawa, T.; et al. Efficiently prepared ephedrine alkaloids-free Ephedra Herb extract: A putative marker and antiproliferative effects. J. Nat. Med. 2016, 70, 554–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanda, Y. Investigation of the freely available easy-to-use software ‘EZR’ for medical statistics. Bone Marrow Transplant. 2013, 48, 452–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwata-Yoshikawa, N.; Kakizaki, M.; Shiwa-Sudo, N.; Okura, T.; Tahara, M.; Fukushi, S.; Maeda, K.; Kawase, M.; Asanuma, H.; Tomita, Y.; et al. Essential role of TMPRSS2 in SARS-CoV-2 infection in murine airways. Nat. Commun. 2022, 13, 6100. [Google Scholar] [CrossRef]
- Suzuki, R.; Yamasoba, D.; Kimura, I.; Wang, L.; Kishimoto, M.; Ito, J.; Morioka, Y.; Nao, N.; Nasser, H.; Uriu, K.; et al. Attenuated fusogenicity and pathogenicity of SARS-CoV-2 Omicron variant. Nature 2022, 603, 700–705. [Google Scholar] [CrossRef] [PubMed]
- Araf, Y.; Akter, F.; Tang, Y.; Fatemi, R.; Alam Parvez, S.; Zheng, C.; Hossain, G. Omicron variant of SARS-CoV-2: Genomics, transmissibility, and responses to current COVID-19 vaccines. J. Med. Virol. 2022, 94, 1825–1832. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, L.B.; Foster, C.; Rawlinson, W.; Tedla, N.; Bull, R.A. Evolution of the SARS-CoV-2 omicron variants BA.1 to BA.5: Implications for immune escape and transmission. Rev. Med. Virol. 2022, 32, e2381. [Google Scholar] [CrossRef] [PubMed]
- Boytz, R.; Słabicki, M.; Ramaswamy, S.; Patten, J.J.; Zou, C.; Meng, C.; Hurst, B.L.; Wang, J.; Nowak, R.P.; Yang, P.L.; et al. Anti-SARS-CoV-2 activity of targeted kinase inhibitors: Repurposing clinically available drugs for COVID-19 therapy. J. Med. Virol. 2022, 95, e28157. [Google Scholar] [CrossRef] [PubMed]
- Naik, R.R.; Shakya, A.K.; Aladwan, S.M.; El-Tanani, M. Kinase Inhibitors as Potential Therapeutic Agents in the Treatment of COVID-19. Front. Pharmacol. 2022, 13, 806568. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| EFE Concentration (µg/mL) | Log10TCID50/mL | Reduction of Log10tcid50.mL | EMCT Concentration (µg/mL) | Log10TCID50/mL | Reduction of Log10tcid50.mL | ||
|---|---|---|---|---|---|---|---|
| control (0) | 5.71 ± 0.55 | ||||||
| 100.00 | ND | >3.91 | ** 1 | 40 | ND | >3.91 | ** 1 |
| 50.00 | 4.05 ± 1.07 | 1.66 | ** 1 | 30 | ND | >3.91 | ** 1 |
| 25.00 | 4.80 ± 0.52 | 0.91 | * 1 | 20 | 3.68 ± 1.23 | 2.03 | ** 2 |
| 12.50 | 4.71 ± 0.27 | 1.00 | ** 1 | 10 | 5.18 ± 0.32 | 0.53 | ** 2 |
| 6.25 | 4.89 ± 0.68 | 0.82 | * 1 | 5 | 5.99 ± 0.24 | −0.28 | 3 |
| 3.13 | 5.08 ± 0.39 | 0.63 | 1 | 2.5 | 5.8 ± 0.20 | −0.09 | 3 |
| EFE Concentration (µg/mL) | Log10TCID50/mL | Reduction of LogTCID50 | EMCT Concentration (mg/mL) | Log10TCID50/mL | Reduction of LogTCID50 | ||
|---|---|---|---|---|---|---|---|
| control (0) | 5.71 ± 0.55 | ||||||
| 100.00 | 3.59 ± 0.77 | 2.12 | ** 1 | 50.00 | 2.11 ± 0.55 | 3.60 | ** 2 |
| 50.00 | 4.13 ± 1.32 | 1.58 | ** 1 | 25.00 | 3.30 ± 1.01 | 2.41 | ** 2 |
| 25.00 | 4.84 ± 0.43 | 0.87 | 1 | 12.50 | 5.18 ± 0.86 | 0.53 | 2 |
| 12.50 | 4.63 ± 0.44 | 1.08 | * 1 | 6.25 | 5.61 ± 0.76 | 0.10 | 2 |
| 6.25 | 4.80 ± 0.27 | 0.91 | 1 | 3.13 | 5.68 ± 0.68 | 0.03 | 2 |
| 3.13 | 4.93 ± 0.63 | 0.78 | 1 |
| Treatment before Infection | Treatment after Infection | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EFE (µg/mL) | EMCT (µg/mL) | EFE (µg/mL) | EMCT (µg/mL) | |||||||||||
| Variant | Control (0) | 100 | Reduction of Log TCID50 | 25 | Reduction of Log TCID50 | 100 | Reduction of Log TCID50 | 25 | Reduction of Log TCID50 | |||||
| QK002 | Alpha | 5.99 ± 0.52 | 2.93 ± 0.83 | 3.06 | ** 1 | 3.18 ± 0.43 | 2.81 | ** 1 | 4.68 ± 0.72 | 1.31 | * 1 | 4.93 ± 0.72 | 1.06 | 1 |
| QHN001 | Alpha | 6.8 ± 0.46 | ND | >5 | ** 1 | ND | >5 | ** 1 | 4.61 ± 1.53 | 2.19 | ** 1 | 5.43 ± 0.85 | 1.37 | 1 |
| TY7-501 | Beta | 6.3 ± 0.61 | ND | >4.5 | ** 1 | ND | >4.5 | ** 1 | 5.24 ± 1.20 | 1.06 | 1 | 4.18 ± 1.48 | 2.12 | * 1 |
| TY7-503 | Beta | 6.43 ± 0.75 | 2.18 ± 0.75 | 4.25 | ** 1 | ND | >4.6 | ** 1 | 5.43 ± 0.75 | 1 | 1 | 4.68 ± 1.33 | 1.75 | * 1 |
| TY8-612 | Gamma | 6.36 ± 0.90 | ND | >4.5 | ** 1 | ND | >4.5 | ** 1 | 4.8 ± 0.84 | 1.56 | ** 1 | 4.43 ± 0.60 | 1.93 | ** 1 |
| TY38-873 | Omicron BA.1 | 5.61 ± 0.48 | 3.36 ± 0.48 | 2.25 | ** 2 | 2.88 ± 0.84 | 2.73 | ** 2 | 3.34 ± 0.85 | 2.27 | ** 3 | 3.64 ± 0.58 | 1.97 | ** 3 |
| TY41-702 | Omicron BA.5 | 6.15 ± 0.42 | 4.11 ± 0.51 | 2.04 | ** 2 | 3.24 ± 0.73 | 2.91 | ** 2 | 3.69 ± 0.79 | 2.46 | ** 3 | 3.86 ± 0.88 | 2.29 | ** 3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Uema, M.; Hyuga, M.; Yonemitsu, K.; Hyuga, S.; Amakura, Y.; Uchiyama, N.; Mizoguchi, K.; Odaguchi, H.; Goda, Y. Antiviral Effect of Ephedrine Alkaloids-Free Ephedra Herb Extract against SARS-CoV-2 In Vitro. Microorganisms 2023, 11, 534. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms11020534
Uema M, Hyuga M, Yonemitsu K, Hyuga S, Amakura Y, Uchiyama N, Mizoguchi K, Odaguchi H, Goda Y. Antiviral Effect of Ephedrine Alkaloids-Free Ephedra Herb Extract against SARS-CoV-2 In Vitro. Microorganisms. 2023; 11(2):534. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms11020534
Chicago/Turabian StyleUema, Masashi, Masashi Hyuga, Kenzo Yonemitsu, Sumiko Hyuga, Yoshiaki Amakura, Nahoko Uchiyama, Kazushige Mizoguchi, Hiroshi Odaguchi, and Yukihiro Goda. 2023. "Antiviral Effect of Ephedrine Alkaloids-Free Ephedra Herb Extract against SARS-CoV-2 In Vitro" Microorganisms 11, no. 2: 534. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms11020534