

Surface Bacterioplankton Community Structure Crossing the Antarctic Circumpolar Current Fronts
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
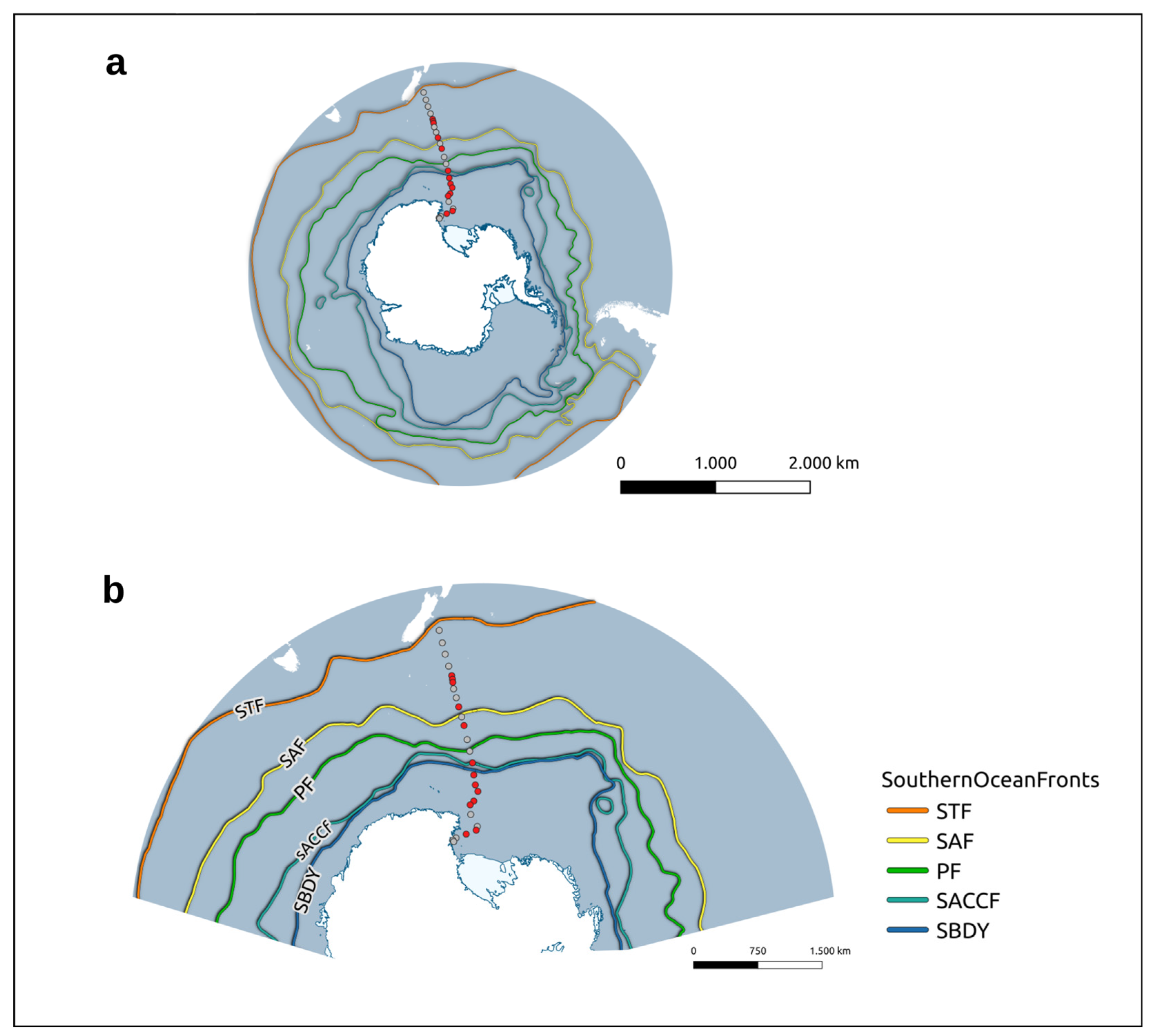
2.1. Sampling Procedure and Study Site
2.2. Environmental Parameters
2.3. Community DNA Extraction
2.4. 16S rRNA Gene Sequencing
2.5. Biochemical Measurements
2.6. Bioinformatics and Statistical Analyses
3. Results
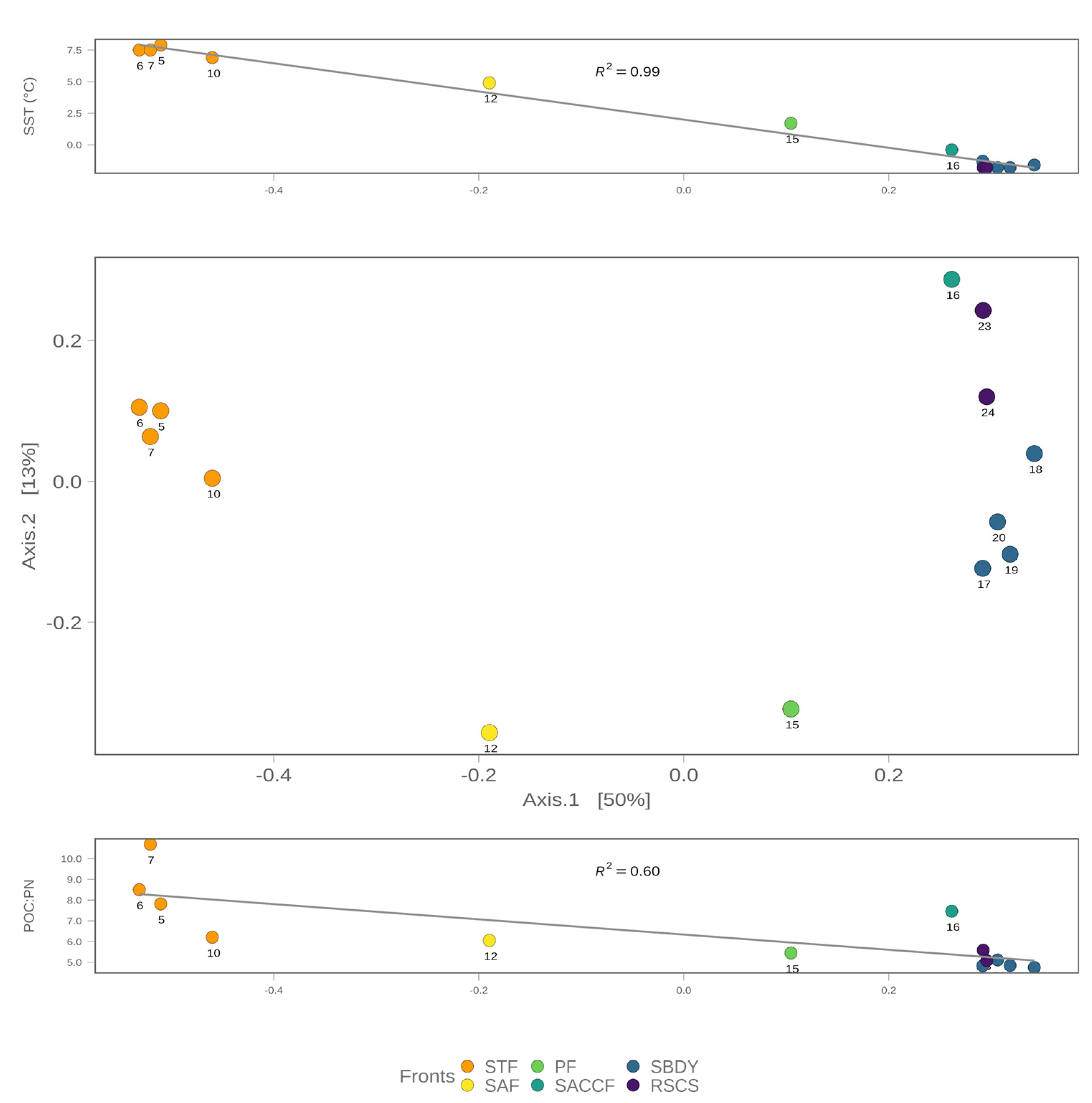
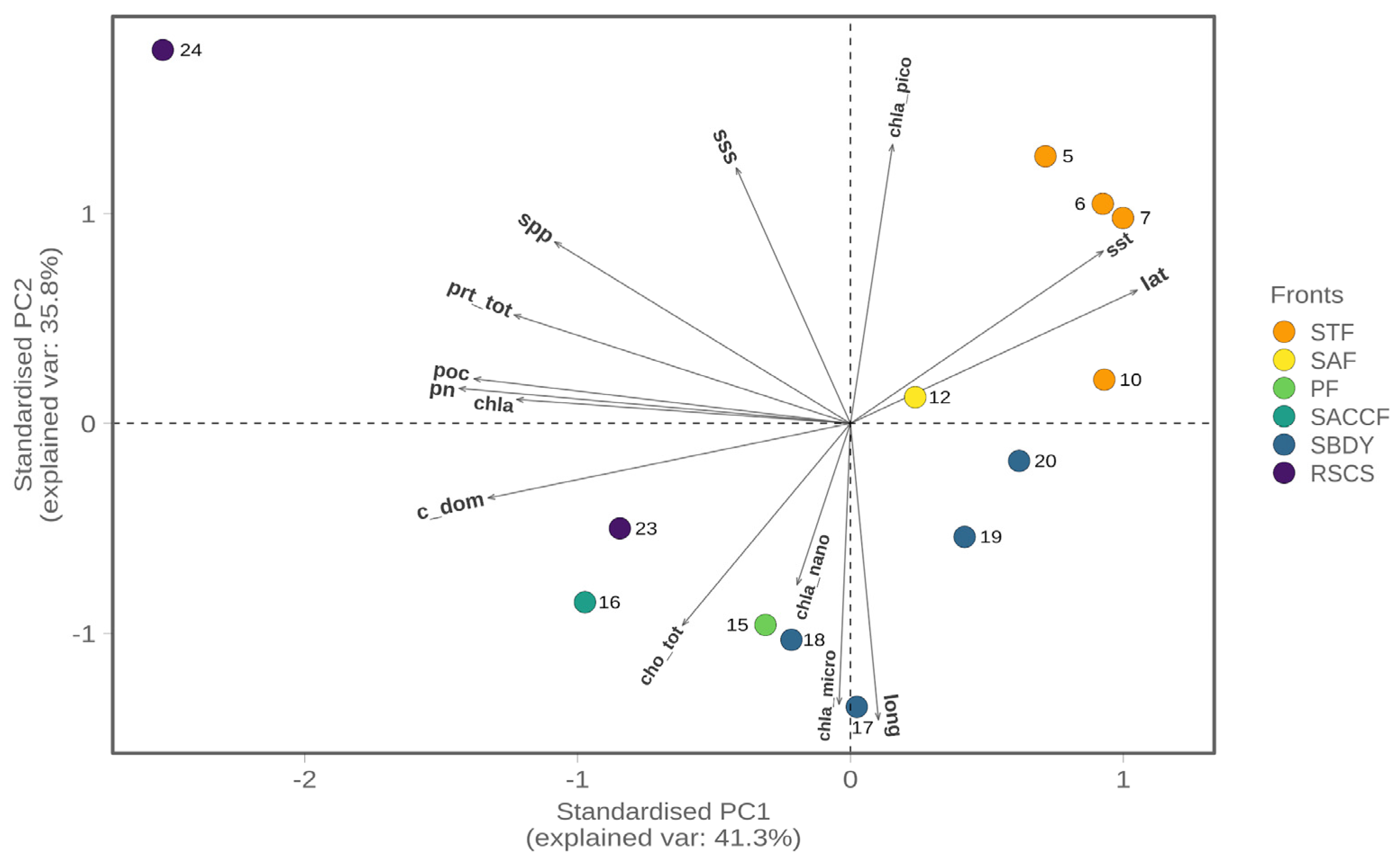
3.1. Environmental and Biogeochemical Parameters
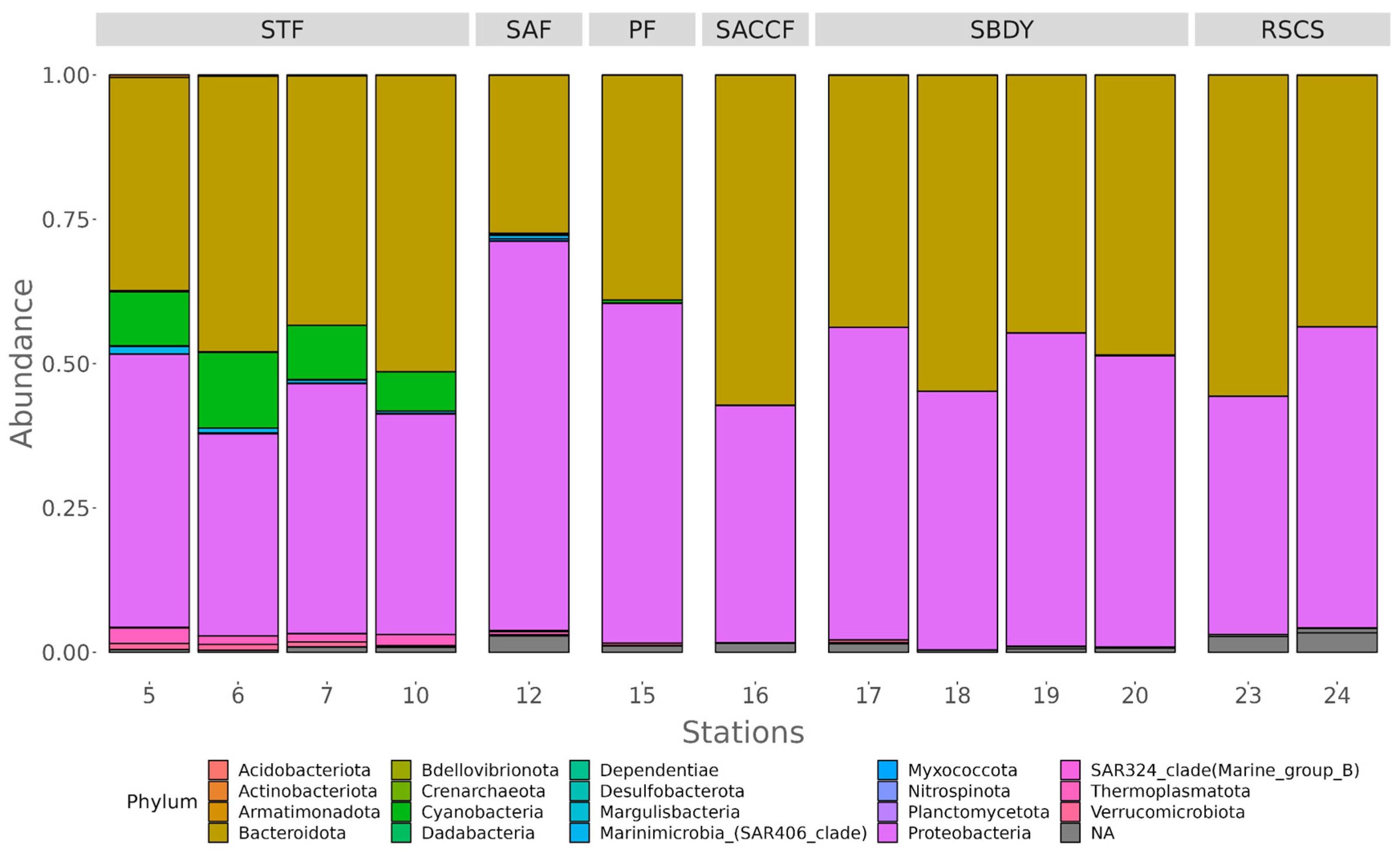
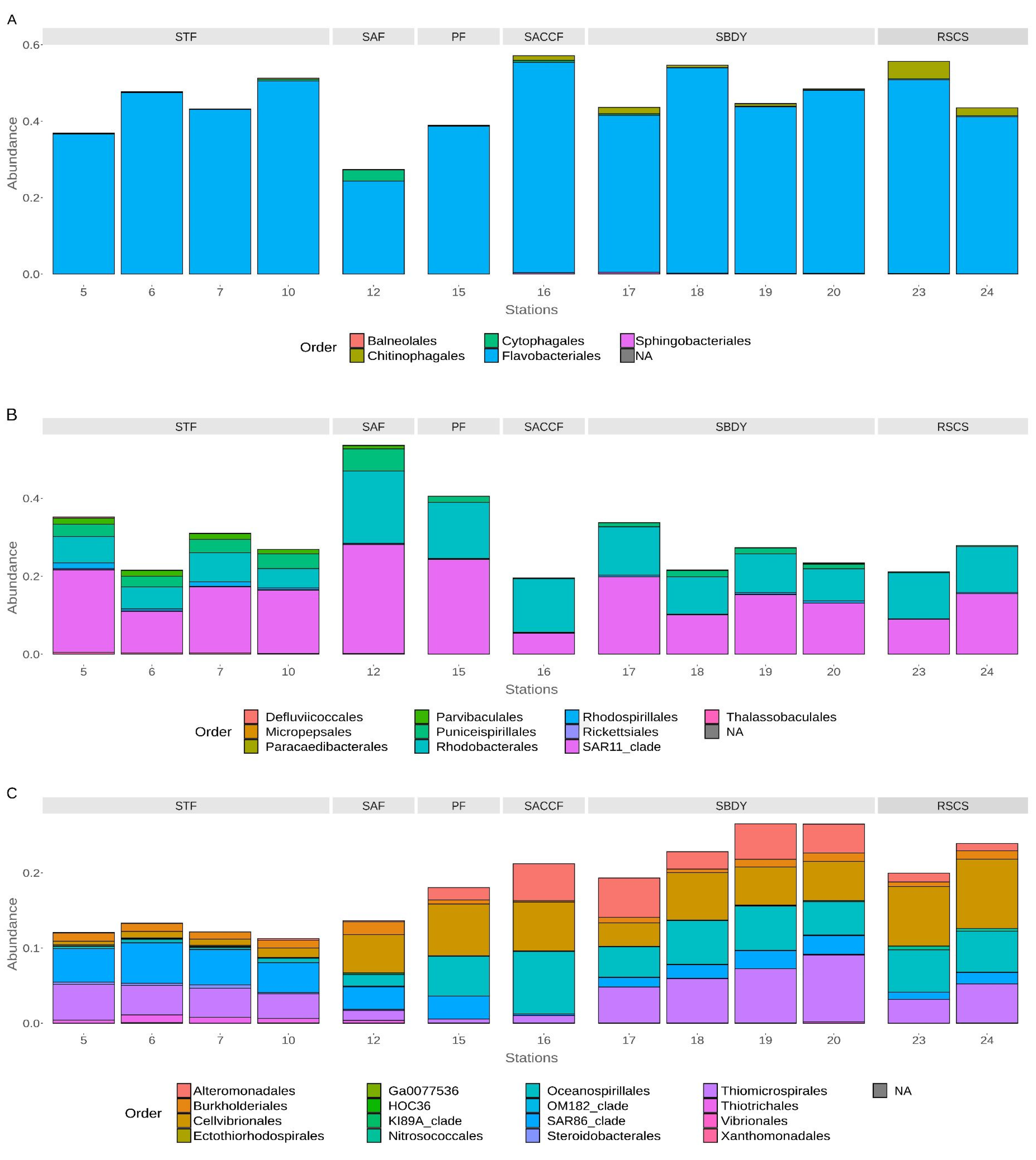
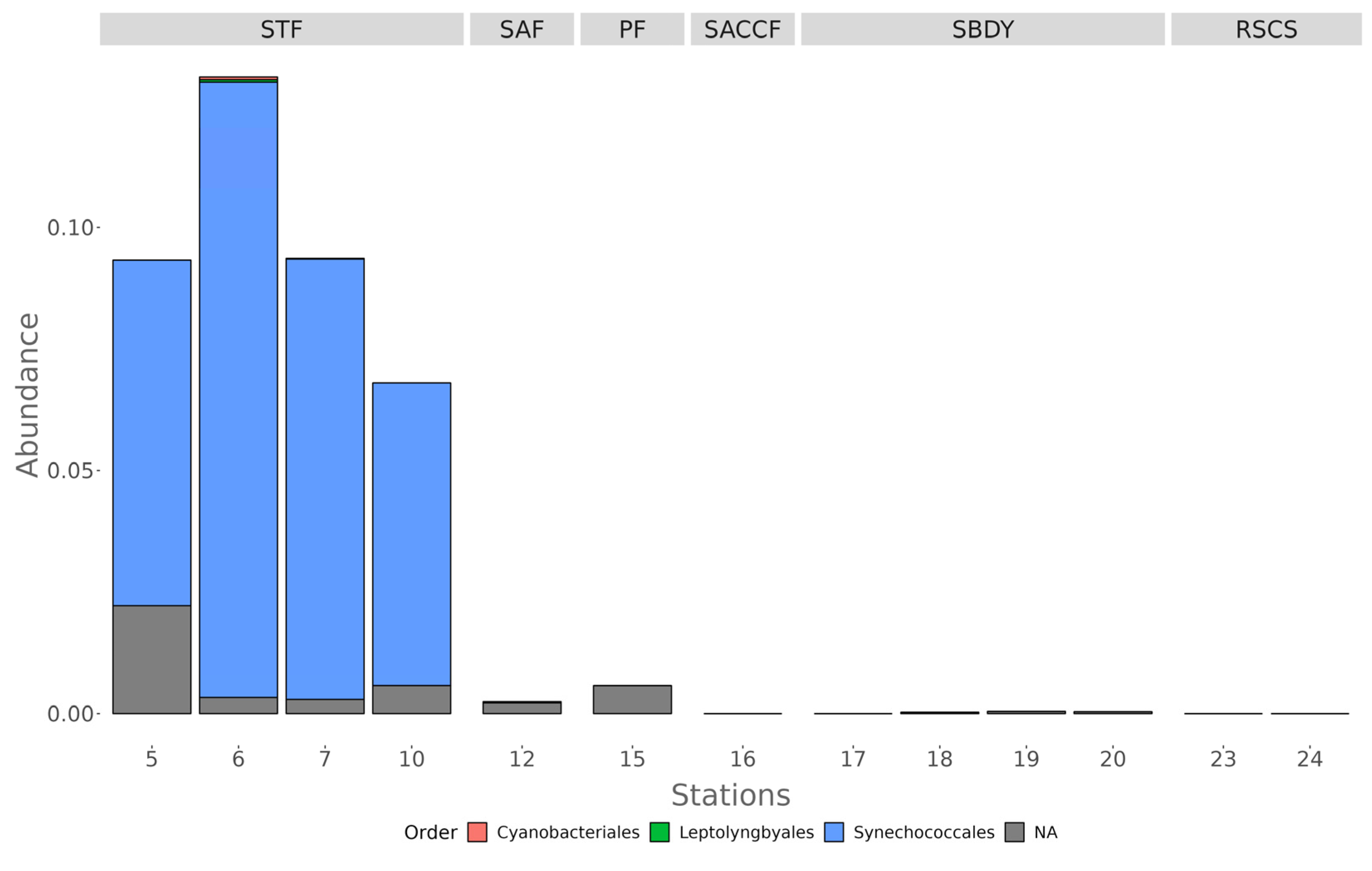
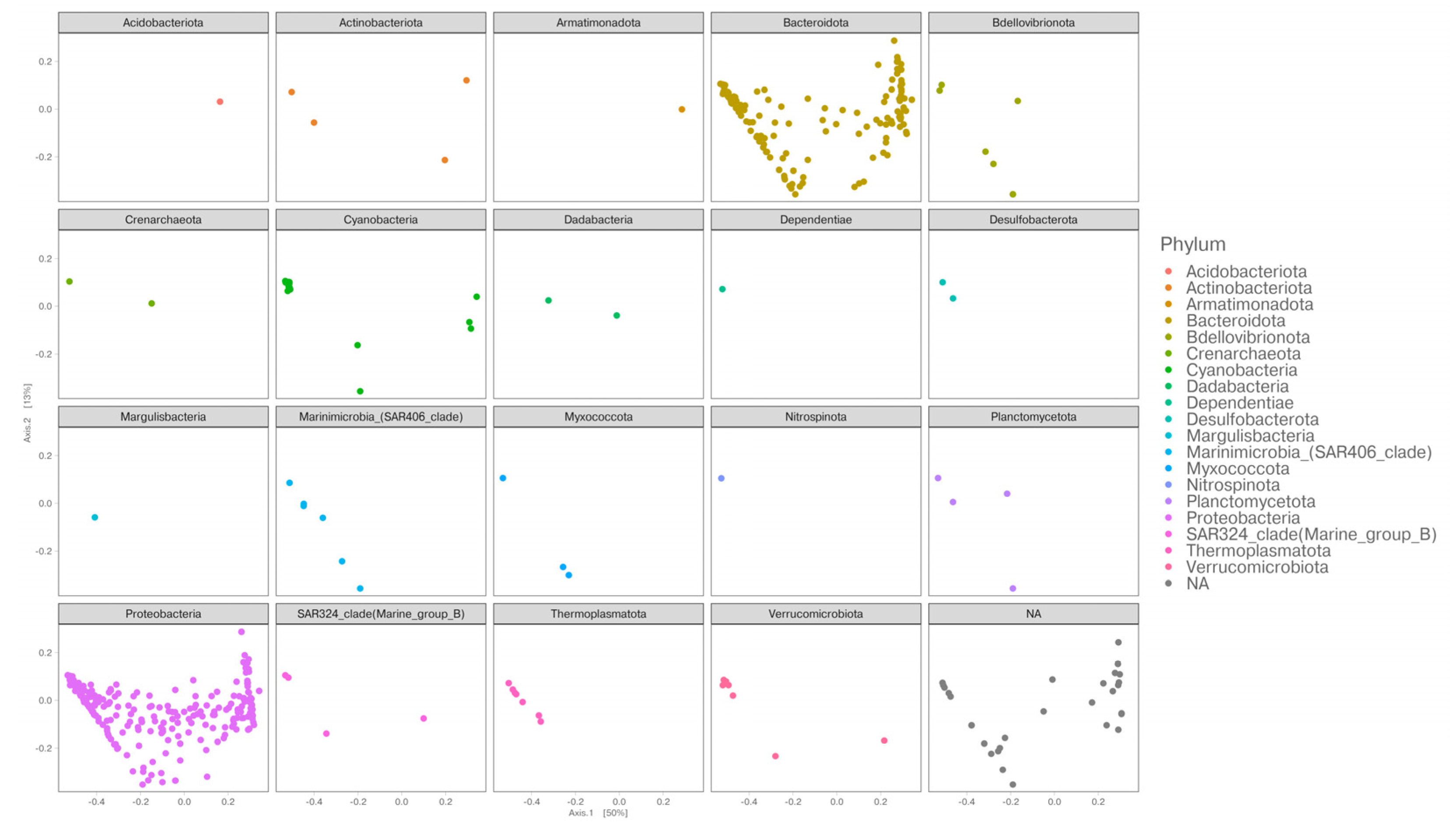
3.2. Diversity of Bacterial Communities
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Azam, F.; Fenchel, T.; Field, J.G.; Gray, J.S.; Meyer-Reil, L.A.; Thingstad, F. The Ecological Role of Water-Column Microbes in the Sea. Mar. Ecol. Prog. Ser. 1983, 10, 257–263. [Google Scholar] [CrossRef]
- Cho, B.; Azam, F. Biogeochemical significance of bacterial biomass in the ocean’s euphotic zone. Mar. Ecol. Prog. Ser. Oldendorf 1990, 63, 253–259. [Google Scholar] [CrossRef]
- Field, C.B.; Behrenfeld, M.J.; Randerson, J.T.; Falkowski, P. Primary Production of the Biosphere: Integrating Terrestrial and Oceanic Components. Science 1998, 281, 237–240. [Google Scholar] [CrossRef] [Green Version]
- Huntley, M.E.; Lopez, M.D.; Karl, D.M. Top Predators in the Southern Ocean: A Major Leak in the Biological Carbon Pump. Science 1991, 253, 64–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapman, C.C.; Lea, M.-A.; Meyer, A.; Sallée, J.-B.; Hindell, M. Defining Southern Ocean fronts and their influence on biological and physical processes in a changing climate. Nat. Clim. Chang. 2020, 10, 209–219. [Google Scholar] [CrossRef]
- Wilkins, D.; Lauro, F.M.; Williams, T.J.; DeMaere, M.; Brown, M.V.; Hoffman, J.M.; Andrews-Pfannkoch, C.; McQuaid, J.; Riddle, M.J.; Rintoul, S.R.; et al. Biogeographic partitioning of Southern Ocean microorganisms revealed by metagenomics. Environ. Microbiol. 2013, 15, 1318–1333. [Google Scholar] [CrossRef]
- Park, Y.; Park, T.; Kim, T.; Lee, S.; Hong, C.; Rio, M.; Pujol, M.; Ballarotta, M.; Durand, I.; Provost, C. Observations of the Antarctic Circumpolar Current over the Udintsev Fracture Zone, the Narrowest Choke Point in the Southern Ocean. J. Geophys. Res. Oceans 2019, 124, 4511–4528. [Google Scholar] [CrossRef] [Green Version]
- Orsi, A.H.; Whitworth, T., III; Nowlin, W.D., Jr. On the Meridional Extent and Fronts of the Antarctic Circumpolar Current. Deep Sea Res. Part Oceanogr. Res. Pap. 1995, 42, 641–673. [Google Scholar] [CrossRef]
- Orsi, A.H.; Wiederwohl, C.L. A recount of Ross Sea waters. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2009, 56, 778–795. [Google Scholar] [CrossRef]
- Graham, R.M.; De Boer, A.M. The Dynamical Subtropical Front. J. Geophys. Res. Oceans 2013, 118, 5676–5685. [Google Scholar] [CrossRef] [Green Version]
- Jacques, G. Dynamics of nutrients and phytoplankton, and fluxes of carbon, nitrogen and silicon in the Antarctic Ocean. Polar Biol. 1992, 12, 149–162. [Google Scholar] [CrossRef]
- Boyd, P.; Robinson, C.; Savidge, G.; Williams, P. Water column and sea-ice primary production during Austral spring in the Bellingshausen Sea. Deep. Sea Res. Part II Top. Stud. Oceanogr. 1995, 42, 1177–1200. [Google Scholar] [CrossRef]
- Van Leeuwe, M.; Scharek, R.; De Baar, H.; De Jong, J.; Goeyens, L. Iron enrichment experiments in the Southern Ocean: Physiological responses of plankton communities. Deep. Sea Res. Part II Top. Stud. Oceanogr. 1997, 44, 189–207. [Google Scholar] [CrossRef]
- van Leeuwe, M.A.; de Baar, H.J.W.; Veldhuis, M.J.W. Pigment distribution in the Pacific region of the Southern Ocean (autumn 1995). Polar Biol. 1998, 19, 348–353. [Google Scholar] [CrossRef]
- De Baar, H.J.W.; de Jong, J.; Bakker, D.; Löscher, B.M.; Veth, C.; Bathmann, U.; Smetacek, V. Importance of iron for plankton blooms and carbon dioxide drawdown in the Southern Ocean. Nature 1995, 373, 412–415. [Google Scholar] [CrossRef]
- Abraham, E.R.; Law, C.S.; Boyd, P.W.; Lavender, S.J.; Maldonado, M.T.; Bowie, A.R. Importance of stirring in the development of an iron-fertilized phytoplankton bloom. Nature 2000, 407, 727–730. [Google Scholar] [CrossRef] [PubMed]
- Ward, P.; Whitehouse, M.; Meredith, M.; Murphy, E.; Shreeve, R.; Korb, R.; Watkins, J.; Thorpe, S.; Woodd-Walker, R.; Brierley, A.; et al. The Southern Antarctic Circumpolar Current Front: Physical and biological coupling at South Georgia. Deep. Sea Res. Part I Oceanogr. Res. Pap. 2002, 49, 2183–2202. [Google Scholar] [CrossRef]
- Green, J.; Bohannan, B.J. Spatial scaling of microbial biodiversity. Trends Ecol. Evol. 2006, 21, 501–507. [Google Scholar] [CrossRef]
- Palumbi, S.R. Genetic Divergence, Reproductive Isolation, and Marine Speciation. Annu. Rev. Ecol. Syst. 1994, 25, 547–572. [Google Scholar] [CrossRef]
- Belkin, I.M.; Gordon, A.L. Southern Ocean fronts from the Greenwich meridian to Tasmania. J. Geophys. Res. Oceans 1996, 101, 3675–3696. [Google Scholar] [CrossRef]
- Ikeda, Y.; Siedler, G.; Zwierz, M. On the variability of Southern Ocean front locations between southern Brazil and the Antarctic Peninsula. J. Geophys. Res. Ocean. 1989, 94, 4757. [Google Scholar] [CrossRef] [Green Version]
- Thornhill, D.J.; Mahon, A.R.; Norenburg, J.L.; Halanych, K.M. Open-ocean barriers to dispersal: A test case with the Antarctic Polar Front and the ribbon worm Parborlasia corrugatus (Nemertea: Lineidae). Mol. Ecol. 2008, 17, 5104–5117. [Google Scholar] [CrossRef] [PubMed]
- Baltar, F.; Currie, K.; Stuck, E.; Roosa, S.; Morales, S.E. Oceanic fronts: Transition zones for bacterioplankton community composition. Environ. Microbiol. Rep. 2016, 8, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Hunter, R.L.; Halanych, K.M. Evaluating Connectivity in the Brooding Brittle Star Astrotoma agassizii across the Drake Passage in the Southern Ocean. J. Hered. 2008, 99, 137–148. [Google Scholar] [CrossRef] [Green Version]
- Shaw, P.; Arkhipkin, A.I.; Al-Khairulla, H. Genetic structuring of Patagonian toothfish populations in the Southwest Atlantic Ocean: The effect of the Antarctic Polar Front and deep-water troughs as barriers to genetic exchange. Mol. Ecol. 2004, 13, 3293–3303. [Google Scholar] [CrossRef]
- Milici, M.; Vital, M.; Tomasch, J.; Badewien, T.H.; Giebel, H.-A.; Plumeier, I.; Wang, H.; Pieper, D.H.; Wagner-Döbler, I.; Simon, M. Diversity and Community Composition of Particle-associated and Free-living Bacteria in Mesopelagic and Bathypelagic Southern Ocean Water Masses: Evidence of Dispersal Limitation in the Bransfield Strait. Limnol. Oceanogr. 2017, 62, 1080–1095. [Google Scholar] [CrossRef]
- Mendes, C.R.B.; Kerr, R.; Tavano, V.M.; Cavalheiro, F.A.; Garcia, C.A.E.; Dessai, D.R.G.; Anilkumar, N. Cross-front phytoplankton pigments and chemotaxonomic groups in the Indian sector of the Southern Ocean. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2015, 118, 221–232. [Google Scholar] [CrossRef]
- Studying the Biodiversity of Marine Plankton|Tara Oceans. Available online: https://fondationtaraocean.org/en/expedition/tara-oceans/ (accessed on 20 February 2023).
- Sunagawa, S.; Coelho, L.P.; Chaffron, S.; Kultima, J.R.; Labadie, K.; Salazar, G.; Djahanschiri, B.; Zeller, G.; Mende, D.R.; Alberti, A.; et al. Structure and function of the global ocean microbiome. Science 2015, 348, 1261359. [Google Scholar] [CrossRef] [Green Version]
- Ocean Productivity: Online Vgpm Data. Available online: http://orca.science.oregonstate.edu/1080.by.2160.monthly.hdf.vgpm.s.chl.a.sst.php (accessed on 26 January 2023).
- Behrenfeld, M.J.; Falkowski, P.G. Photosynthetic rates derived from satellite-based chlorophyll concentration. Limnol. Oceanogr. 1997, 42, 1–20. [Google Scholar] [CrossRef]
- AVHRR Pathfinder SST. Available online: https://www.ncei.noaa.gov/products/avhrr-pathfinder-sst (accessed on 26 January 2023).
- OSCAR—Earth and Space Research. Available online: https://www.esr.org/research/oscar/ (accessed on 26 January 2023).
- RSS SMAP Level 3 Sea Surface Salinity Standard Mapped Image Monthly V4.0 Validated Dataset. Available online: https://podaac.jpl.nasa.gov/dataset/SMAP_RSS_L3_SSS_SMI_MONTHLY_V4 (accessed on 26 January 2023).
- Cordone, A.; Lucchini, S.; De Felice, M.; Ricca, E. Direct and indirect control of Lrp on LEE pathogenicity genes of Citrobacter rodentium. FEMS Microbiol. Lett. 2011, 325, 64–70. [Google Scholar] [CrossRef] [Green Version]
- Zanfardino, A.; Migliardi, A.; D’Alonzo, D.; Lombardi, A.; Varcamonti, M.; Cordone, A. Inactivation of MSMEG_0412 gene drastically affects surface related properties of Mycobacterium smegmatis. BMC Microbiol. 2016, 16, 267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cordone, A.; Mauriello, E.M.F.; Pickard, D.J.; Dougan, G.; De Felice, M.; Ricca, E. The lrp Gene and Its Role in Type I Fimbriation in Citrobacter rodentium. J. Bacteriol. 2005, 187, 7009–7017. [Google Scholar] [CrossRef] [Green Version]
- Cordone, A.; D’Errico, G.; Magliulo, M.; Bolinesi, F.; Selci, M.; Basili, M.; de Marco, R.; Saggiomo, M.; Rivaro, P.; Giovannelli, D.; et al. Bacterioplankton Diversity and Distribution in Relation to Phytoplankton Community Structure in the Ross Sea Surface Waters. Front. Microbiol. 2022, 13, 39. [Google Scholar] [CrossRef]
- Buglione, M.; Ricca, E.; Petrelli, S.; Baccigalupi, L.; Troiano, C.; Saggese, A.; Rivieccio, E.; Fulgione, D. Gut microbiota plasticity in insular lizards under reversed island syndrome. Sci. Rep. 2022, 12, 12682. [Google Scholar] [CrossRef]
- Integrated Microbiome Resource (IMR). Available online: https://imr.bio/ (accessed on 30 January 2023).
- Hedges, J.I.; Stern, J.H. Carbon and nitrogen determinations of carbonate-containing solids1. Limnol. Oceanogr. 1984, 29, 657–663. [Google Scholar] [CrossRef]
- Mangoni, O.; Saggiomo, V.; Bolinesi, F.; Margiotta, F.; Budillon, G.; Cotroneo, Y.; Misic, C.; Rivaro, P.; Saggiomo, M. Phytoplankton blooms during austral summer in the Ross Sea, Antarctica: Driving factors and trophic implications. PLoS ONE 2017, 12, e0176033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holm-Hansen, O.; Lorenzen, C.J.; Holmes, R.W.; Strickland, J.D.H. Fluorometric Determination of Chlorophyll. ICES J. Mar. Sci. 1965, 30, 3–15. [Google Scholar] [CrossRef]
- Manini, E.; Baldrighi, E.; Ricci, F.; Grilli, F.; Giovannelli, D.; Intoccia, M.; Casabianca, S.; Capellacci, S.; Marinchel, N.; Penna, P.; et al. Assessment of Spatio-Temporal Variability of Faecal Pollution along Coastal Waters during and after Rainfall Events. Water 2022, 14, 502. [Google Scholar] [CrossRef]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Hartree, E. Determination of protein: A modification of the lowry method that gives a linear photometric response. Anal. Biochem. 1972, 48, 422–427. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Holmes, S.P. Exact sequence variants should replace operational taxonomic units in marker-gene data analysis. ISME J. 2017, 11, 2639–2643. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [Green Version]
- Sheik, C.S.; Reese, B.K.; Twing, K.I.; Sylvan, J.B.; Grim, S.L.; Schrenk, M.O.; Sogin, M.L.; Colwell, F.S. Identification and Removal of Contaminant Sequences from Ribosomal Gene Databases: Lessons From the Census of Deep Life. Front. Microbiol. 2018, 9, 840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Oksanen, J.; Simpson, G.; Blanchet, F.; Kindt, R.; Legendre, P.; Minchin, P.; O’Hara, R.; Solymos, P.; Stevens, M.; Szoecs, E.; et al. Vegan: Community Ecology Package 2022. Available online: https://github.com/vegandevs/vegan (accessed on 12 October 2022).
- Wickham, H. Ggplot2. WIREs Comput. Stat. 2011, 3, 180–185. [Google Scholar] [CrossRef]
- Morton, J.T.; Toran, L.; Edlund, A.; Metcalf, J.L.; Lauber, C.; Knight, R. Uncovering the Horseshoe Effect in Microbial Analyses. Msystems 2017, 2, e00166-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danis, B.; Van De Putte, A.; Convey, P.; Griffiths, H.; Linse, K.; Murray, A.E. Editorial: Antarctic Biology: Scale Matters. Front. Ecol. Evol. 2020, 8, 91. [Google Scholar] [CrossRef] [Green Version]
- Lim, Y.; Kang, I.; Cho, J.-C. Genome characteristics of Kordia antarctica IMCC3317T and comparative genome analysis of the genus Kordia. Sci. Rep. 2020, 10, 14715. [Google Scholar] [CrossRef]
- Sadaiappan, B.; Kannan, S.; Palaniappan, S.; Manikkam, R.; Ramasamy, B.; Anilkumar, N.; Subramanian, M. Metagenomic 16S rDNA amplicon data of microbial diversity and its predicted metabolic functions in the Southern Ocean (Antarctic). Data Brief 2020, 28, 104876. [Google Scholar] [CrossRef]
- Giebel, H.-A.; Brinkhoff, T.; Zwisler, W.; Selje, N.; Simon, M. Distribution of Roseobacter RCA and SAR11 lineages and distinct bacterial communities from the subtropics to the Southern Ocean. Environ. Microbiol. 2009, 11, 2164–2178. [Google Scholar] [CrossRef]
- Obernosterer, I.; Catala, P.; Lebaron, P.; West, N.J. Distinct bacterial groups contribute to carbon cycling during a naturally iron fertilized phytoplankton bloom in the Southern Ocean. Limnol. Oceanogr. 2011, 56, 2391–2401. [Google Scholar] [CrossRef]
- Kong, L.-F.; He, Y.-B.; Xie, Z.-X.; Luo, X.; Zhang, H.; Yi, S.-H.; Lin, Z.-L.; Zhang, S.-F.; Yan, K.-Q.; Xu, H.-K.; et al. Illuminating Key Microbial Players and Metabolic Processes Involved in the Remineralization of Particulate Organic Carbon in the Ocean’s Twilight Zone by Metaproteomics. Environ. Microbiol. 2021, 87, e00986-21. [Google Scholar] [CrossRef] [PubMed]
- Kirchman, D.L. The Ecology of Cytophaga–Flavobacteria in aquatic environments. FEMS Microbiol. Ecol. 2002, 39, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Pinhassi, J.; Sala, M.M.; Havskum, H.; Peters, F.; Guadayol, O.; Malits, A.; Marrase, C. Changes in Bacterioplankton Composition under Different Phytoplankton Regimens. Appl. Environ. Microbiol. 2004, 70, 6753–6766. [Google Scholar] [CrossRef] [Green Version]
- Abell, G.C.; Bowman, J.P. Ecological and biogeographic relationships of class Flavobacteria in the Southern Ocean. FEMS Microbiol. Ecol. 2005, 51, 265–277. [Google Scholar] [CrossRef] [Green Version]
- Abell, G.; Bowman, J.P. Colonization and community dynamics of class Flavobacteria on diatom detritus in experimental mesocosms based on Southern Ocean Seawater. FEMS Microbiol. Ecol. 2005, 53, 379–391. [Google Scholar] [CrossRef]
- Williams, T.J.; Long, E.; Evans, F.; DeMaere, M.; Lauro, F.M.; Raftery, M.J.; Ducklow, H.; Grzymski, J.J.; Murray, A.E.; Cavicchioli, R. A metaproteomic assessment of winter and summer bacterioplankton from Antarctic Peninsula coastal surface waters. ISME J. 2012, 6, 1883–1900. [Google Scholar] [CrossRef] [Green Version]
- West, N.J.; Obernosterer, I.; Zemb, O.; Lebaron, P. Major differences of bacterial diversity and activity inside and outside of a natural iron-fertilized phytoplankton bloom in the Southern Ocean. Environ. Microbiol. 2008, 10, 738–756. [Google Scholar] [CrossRef]
- Williams, T.J.; Wilkins, D.; Long, E.; Evans, F.; DeMaere, M.Z.; Raftery, M.J.; Cavicchioli, R. The role of planktonic Flavobacteria in processing algal organic matter in coastal East Antarctica revealed using metagenomics and metaproteomics. Environ. Microbiol. 2013, 15, 1302–1317. [Google Scholar] [CrossRef] [PubMed]
- Berner, C.; Bertos-Fortis, M.; Pinhassi, J.; Legrand, C. Response of Microbial Communities to Changing Climate Conditions During Summer Cyanobacterial Blooms in the Baltic Sea. Front. Microbiol. 2018, 9, 1562. [Google Scholar] [CrossRef]
- Auger, M.; Morrow, R.; Kestenare, E.; Sallée, J.-B.; Cowley, R. Southern Ocean in-situ temperature trends over 25 years emerge from interannual variability. Nat. Commun. 2021, 12, 514. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Station | Latitude [°N] | Longitude [°E] | SST [°C] | SSTA [°C] | SSS [PSU] | cs [m/s] | ws [km/h] | Antarctic Current | Oceanic Front |
|---|---|---|---|---|---|---|---|---|---|
| 5 | −51.8092 | 175.0061 | 7.9 | −0.8 | 34.4 | 0.32 | 62 | NSAF | STF |
| 6 | −52.3100 | 175.0749 | 7.5 | −0.7 | 34.3 | 0.29 | 14 | NSAF | STF |
| 7 | −52.7343 | 175.0708 | 7.5 | −0.7 | 34.3 | 0.24 | 15 | NSAF | STF |
| 10 | −56.1723 | 175.8037 | 6.9 | 0 | 33.9 | 0.1 | 26 | SSAF | STF |
| 12 | −58.8349 | 176.6923 | 4.9 | −0.3 | 34 | 0.06 | 37 | SSAF | SAF |
| 15 | −64.2124 | 178.3410 | 1.7 | 0.6 | 33.8 | 0.24 | 39 | PF | PF |
| 16 | −65.9702 | 178.5408 | −0.4 | 0 | 33.6 | 0.01 | 30 | SACCF | SACCF |
| 17 | −67.4229 | 178.9109 | −1.3 | −0.3 | 33 | 0.01 | 13 | SACCF | sBdy |
| 18 | −68.3780 | 179.7789 | −1.6 | −0.5 | 33.2 | 0.02 | 16 | SACCF | sBdy |
| 19 | −69.7571 | 177.9027 | −1.8 | −0.6 | 33.8 | 0.03 | 17 | SACCF | sBdy |
| 20 | −70.2680 | 176.2240 | −1.8 | −0.7 | 34.3 | 0.03 | 7 | SACCF | sBdy |
| 23 | −74.0641 | 178.1007 | −1.8 | −1.5 | 34.5 | 0.05 | 17 | RSCS | RSCS |
| 24 | −74.4742 | 172.4104 | −1.8 | −1.7 | 35.2 | 0.03 | 23 | RSCS | RSCS |
| Station | ssp [mg C/m2 Day] | prt_tot [µg/L] | cho_tot [µg/L] | PN [µg/L] | POC [µg/L] | C-DOM [µg/L] | Chl-a [µg/L] | Micro- Chla [%] | Nano- Chla [%] | Pico-Chla [%] |
|---|---|---|---|---|---|---|---|---|---|---|
| 5 | 398.14 | 66.84 | 41.60 | 9.00 | 69.77 | 11.11 | 0.32 | 12.98 | 30.81 | 56.20 |
| 6 | 323.15 | 56.58 | 35.98 | 6.71 | 57.22 | 8.12 | 0.41 | 1.63 | 54.14 | 44.23 |
| 7 | 290.04 | 48.64 | 33.71 | 4.91 | 52.76 | 8.70 | 0.58 | 12.66 | 40.06 | 47.28 |
| 10 | 282.89 | 46.76 | 18.05 | 7.38 | 46.41 | 8.49 | 0.69 | 14.22 | 64.64 | 21.14 |
| 12 | 265.15 | 56.56 | 34.37 | 14.87 | 90.08 | 11.72 | 1.44 | 15.28 | 56.38 | 28.33 |
| 15 | 211.15 | 56.81 | 58.67 | 17.94 | 97.71 | 16.74 | 1.56 | 23.13 | 62.66 | 14.21 |
| 16 | 275.82 | 98.02 | 63.21 | 28.90 | 216.08 | 17.41 | 1.06 | 24.51 | 56.68 | 18.81 |
| 17 | 285.45 | 49.86 | 59.06 | 16.59 | 79.81 | 14.90 | 0.55 | 31.24 | 51.44 | 17.32 |
| 18 | 249.01 | 53.63 | 54.32 | 22.27 | 106.55 | 12.12 | 1.44 | 21.42 | 55.29 | 23.29 |
| 19 | 244.42 | 36.57 | 43.70 | 10.49 | 50.87 | 10.08 | 0.63 | 19.80 | 51.32 | 28.88 |
| 20 | 103.30 | 23.87 | 31.64 | 8.92 | 45.32 | 10.25 | 0.55 | 14.85 | 54.89 | 30.26 |
| 23 | 475.60 | 71.14 | 64.82 | 20.80 | 116.38 | 18.08 | 0.99 | 20.91 | 53.31 | 25.78 |
| 24 | 943.99 | 124.22 | 34.73 | 54.01 | 274.66 | 19.13 | 2.47 | 3.15 | 49.46 | 47.39 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cordone, A.; Selci, M.; Barosa, B.; Bastianoni, A.; Bastoni, D.; Bolinesi, F.; Capuozzo, R.; Cascone, M.; Correggia, M.; Corso, D.; et al. Surface Bacterioplankton Community Structure Crossing the Antarctic Circumpolar Current Fronts. Microorganisms 2023, 11, 702. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms11030702
Cordone A, Selci M, Barosa B, Bastianoni A, Bastoni D, Bolinesi F, Capuozzo R, Cascone M, Correggia M, Corso D, et al. Surface Bacterioplankton Community Structure Crossing the Antarctic Circumpolar Current Fronts. Microorganisms. 2023; 11(3):702. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms11030702
Chicago/Turabian StyleCordone, Angelina, Matteo Selci, Bernardo Barosa, Alessia Bastianoni, Deborah Bastoni, Francesco Bolinesi, Rosaria Capuozzo, Martina Cascone, Monica Correggia, Davide Corso, and et al. 2023. "Surface Bacterioplankton Community Structure Crossing the Antarctic Circumpolar Current Fronts" Microorganisms 11, no. 3: 702. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms11030702