
Biogenic Silver Nanoparticles Produced by Soil Rare Actinomycetes and Their Significant Effect on Aspergillus-derived mycotoxins

 ,
,  , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Actinobacteria
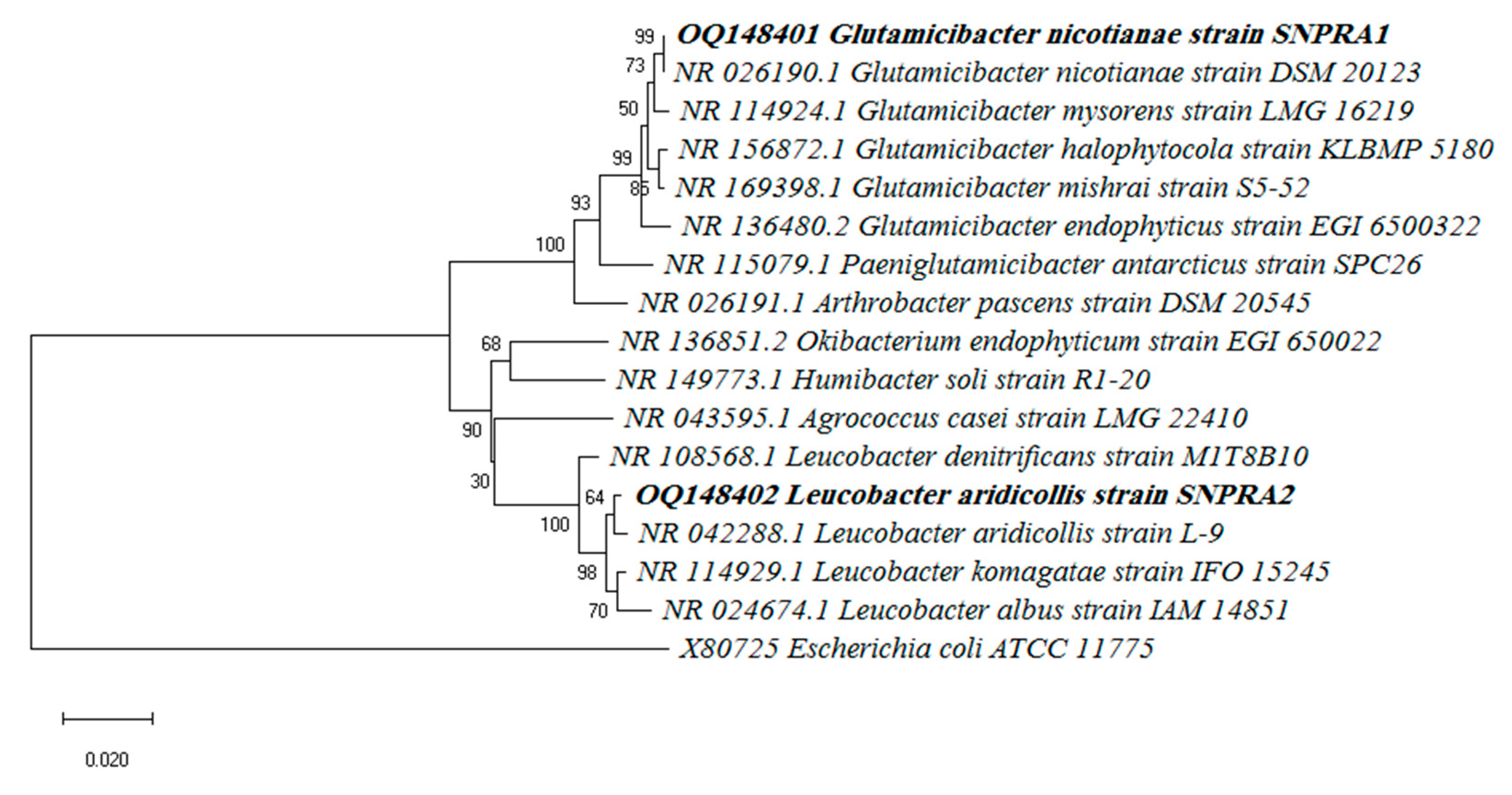
2.2. Identification and Phylogenetic Analysis
2.3. Biosynthesis of AgNPs
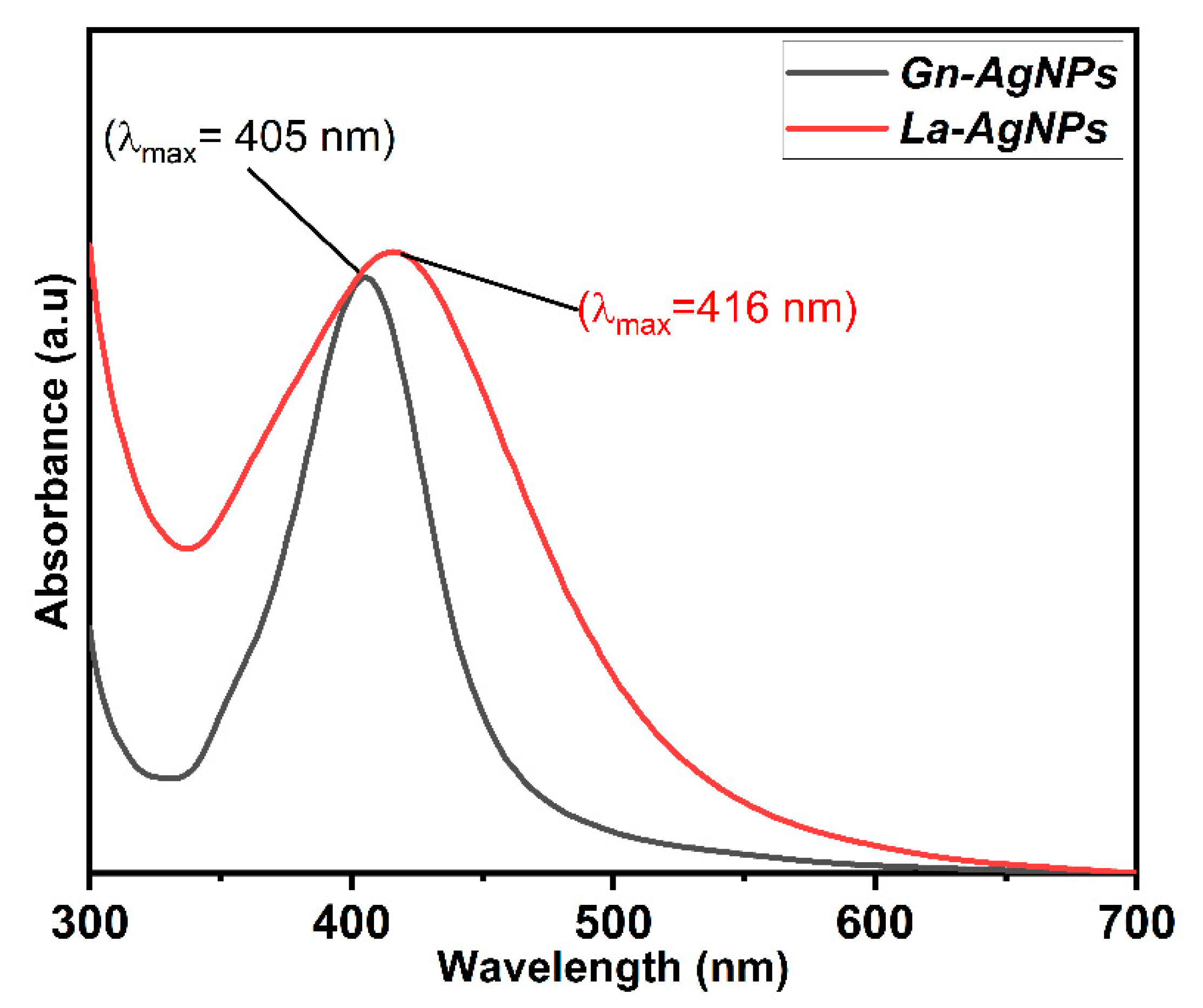
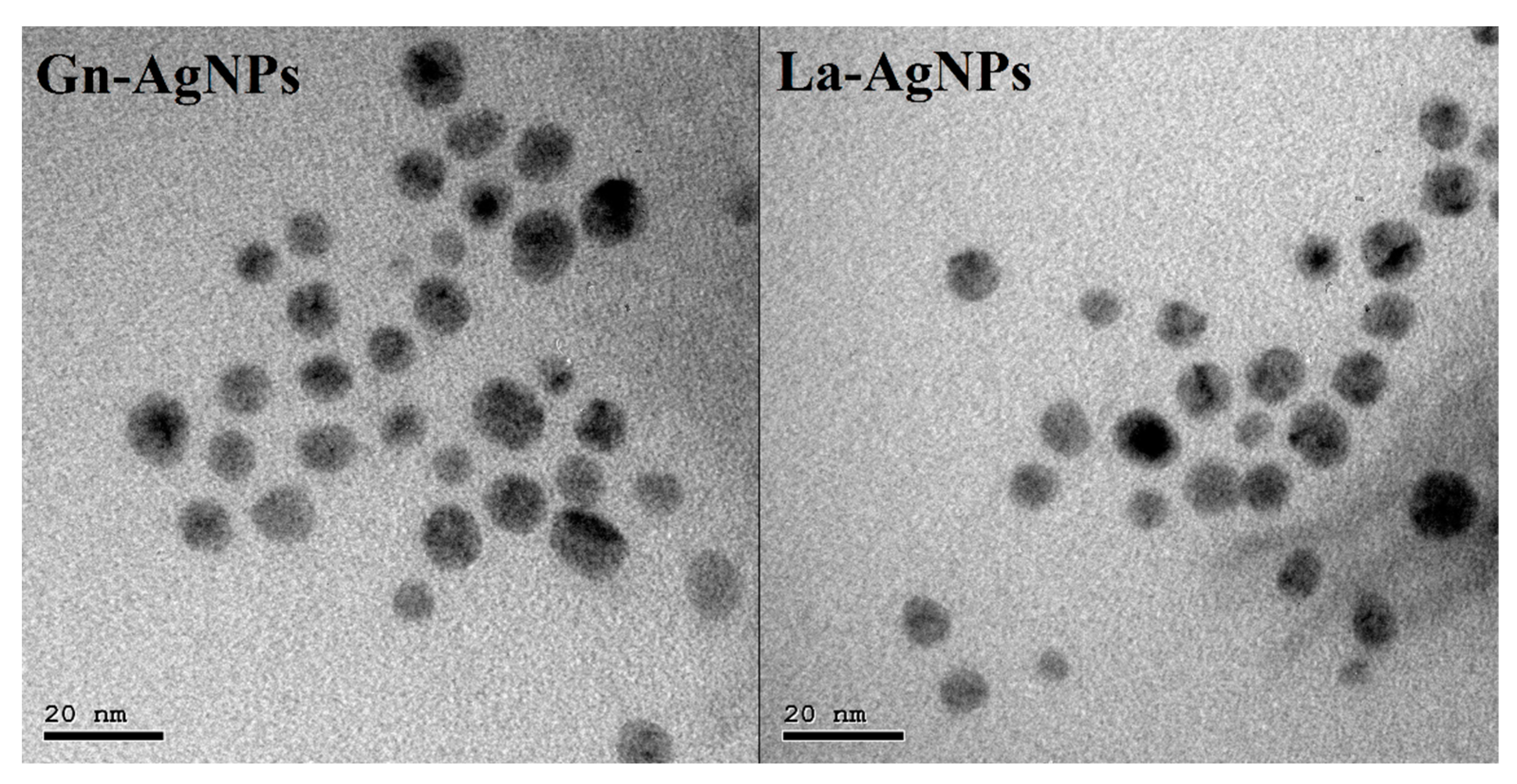
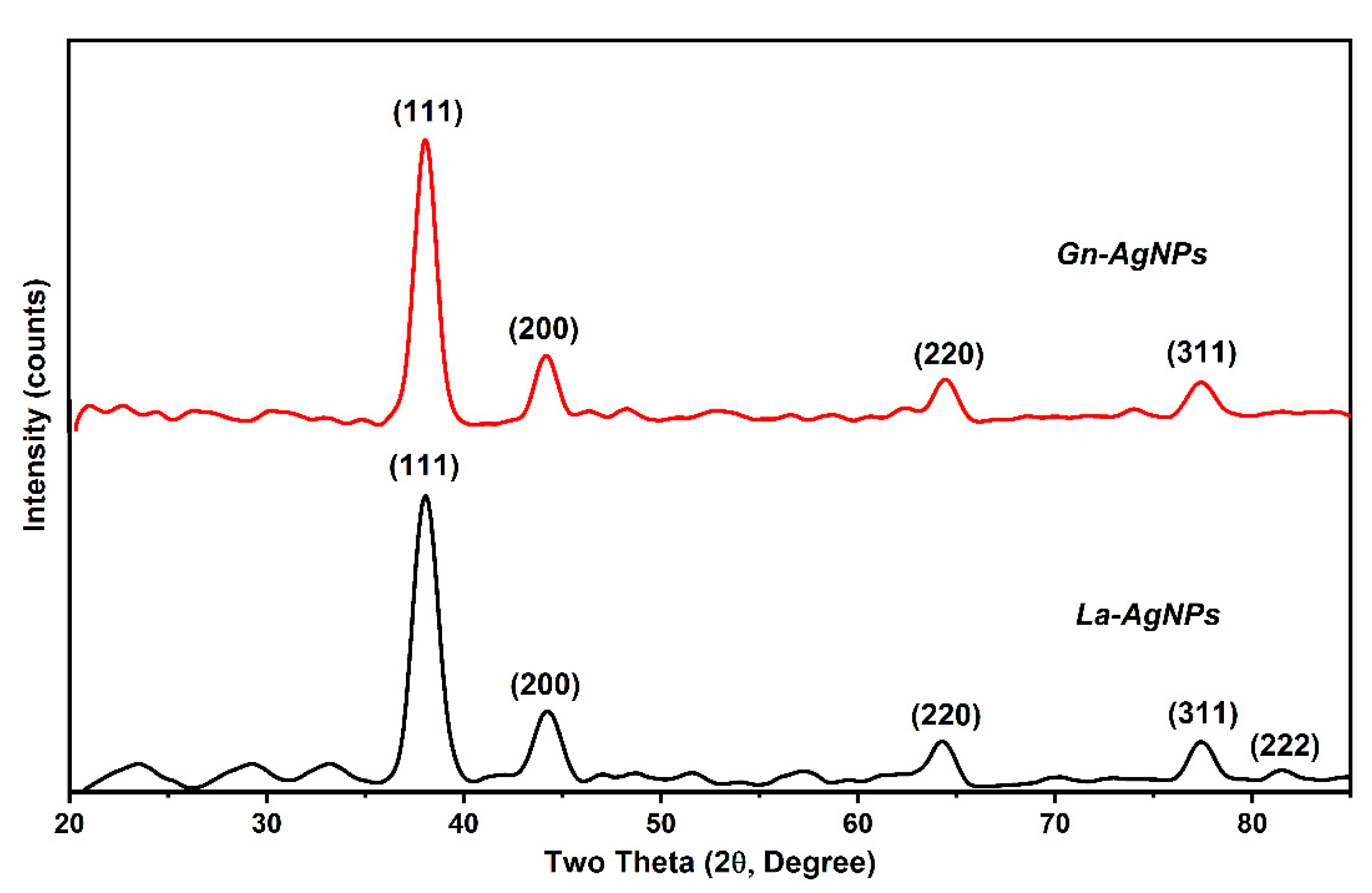
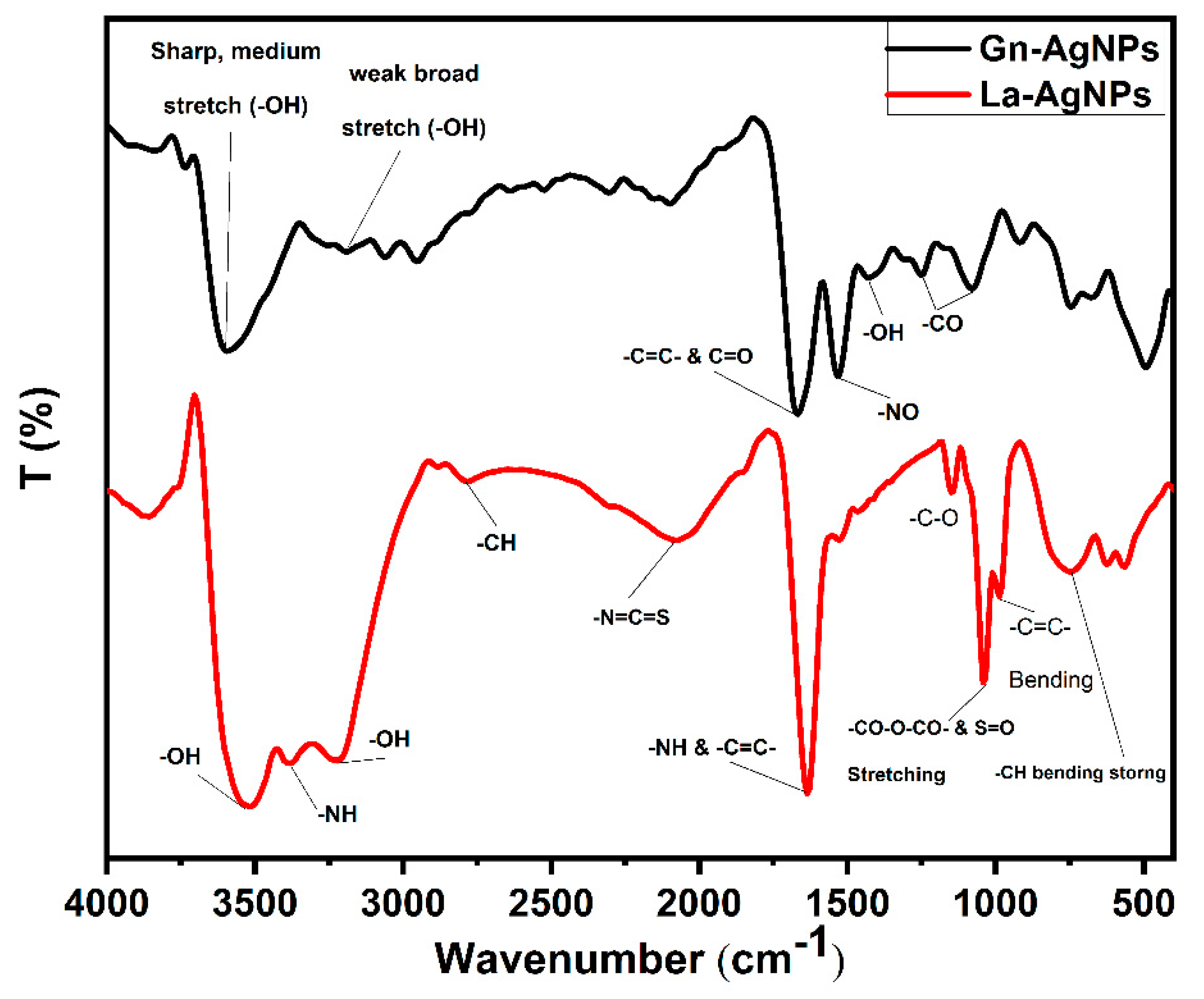
2.4. Characterization of Biogenic AgNPs
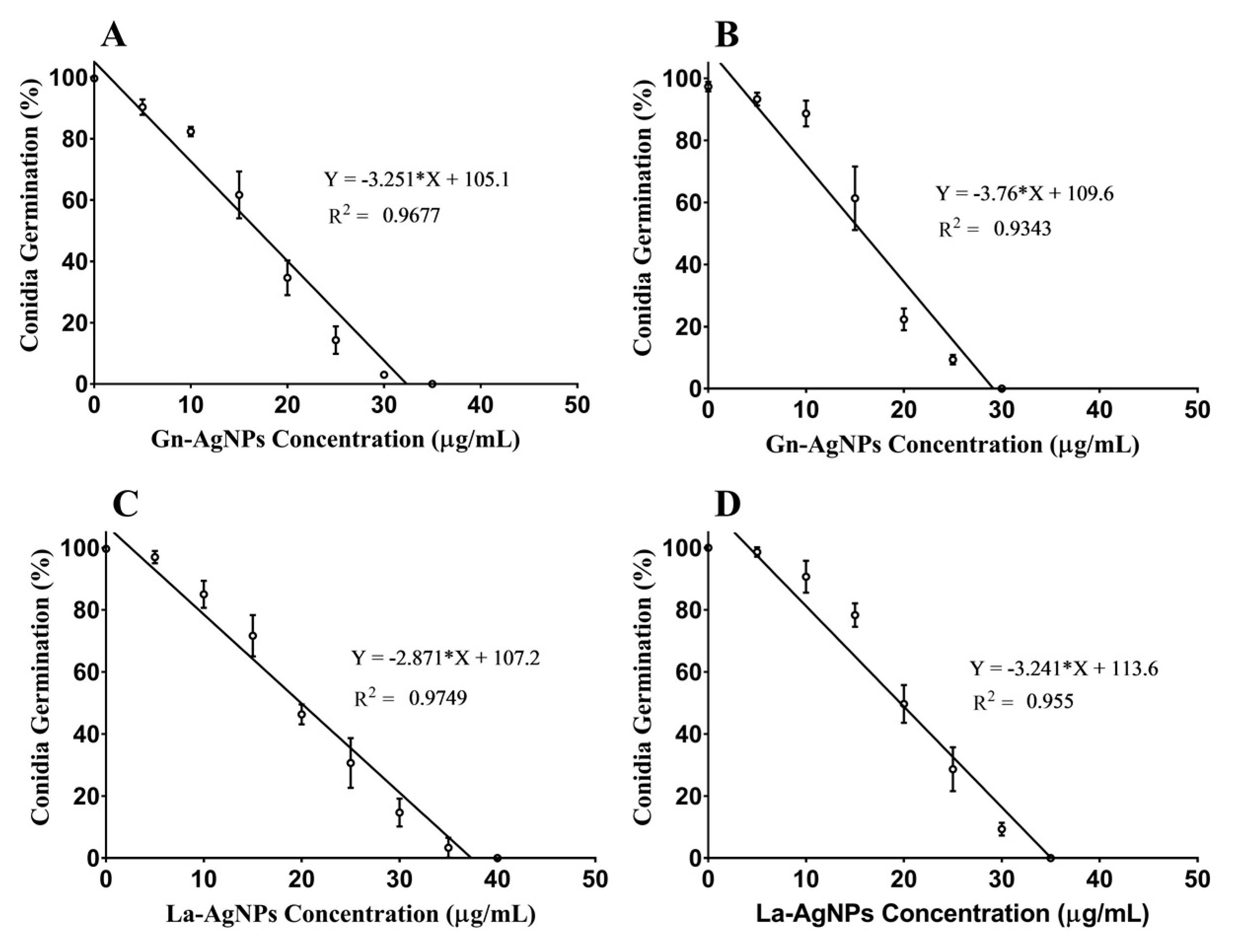
2.5. Antifungal Activity
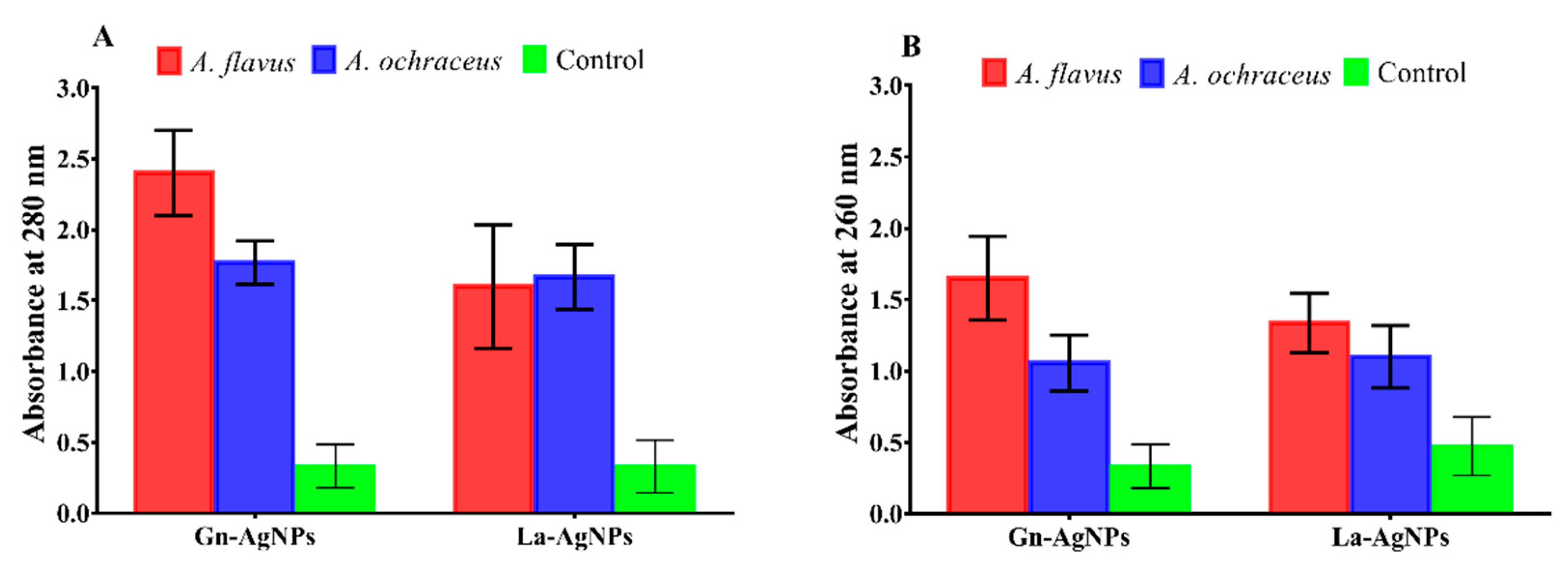
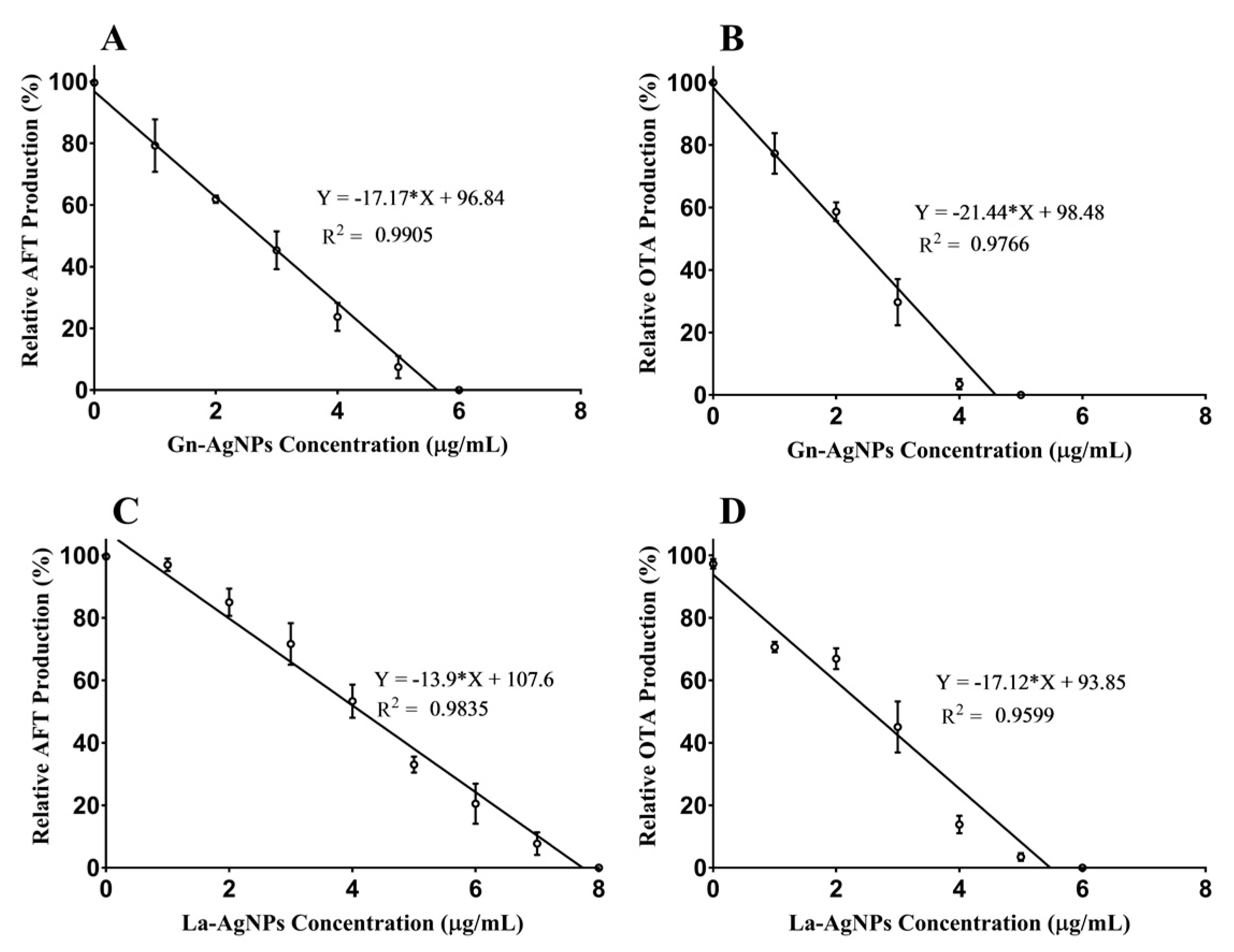
2.6. Effect of Biogenic AgNPs on Mycotoxin Production
2.7. Cytotoxicity Assay
2.8. Statistical Analysis
3. Results
3.1. Isolation of Actinobacteria
3.2. Identification and Phylogenetic Analysis
3.3. Biosynthesis of AgNPs
3.4. Characterization of Biogenic AgNPs
3.5. Antifungal Activity
3.6. Effect of Biogenic AgNPs on Mycotoxin Production
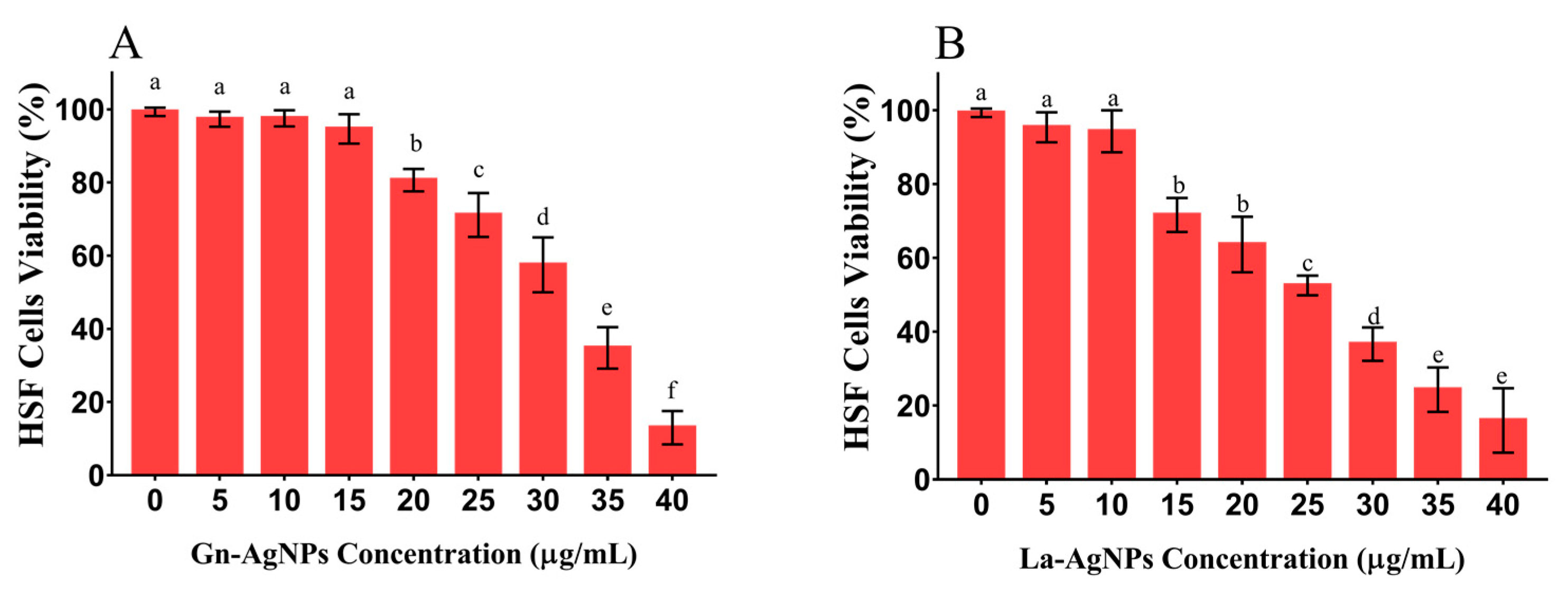
3.7. Cytotoxicity Assay
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Khan, Y.; Sadia, H.; Ali Shah, S.Z.; Khan, M.N.; Shah, A.A.; Ullah, N.; Ullah, M.F.; Bibi, H.; Bafakeeh, O.T.; Khedher, N.B.; et al. Classification, Synthetic, and Characterization Approaches to Nanoparticles, and Their Applications in Various Fields of Nanotechnology: A Review. Catalysts 2022, 12, 1386. [Google Scholar] [CrossRef]
- Chandrakala, V.; Aruna, V.; Angajala, G. Review on Metal Nanoparticles as Nanocarriers: Current Challenges and Perspectives in Drug Delivery Systems. Emergent Mater. 2022, 5, 1593–1615. [Google Scholar] [CrossRef]
- Saravanan, A.; Kumar, P.S.; Karishma, S.; Vo, D.V.N.; Jeevanantham, S.; Yaashikaa, P.R.; George, C.S. A Review on Biosynthesis of Metal Nanoparticles and Its Environmental Applications. Chemosphere 2021, 264, 128580. [Google Scholar] [CrossRef] [PubMed]
- Couto, C.; Almeida, A. Metallic Nanoparticles in the Food Sector: A Mini-Review. Foods 2022, 11, 402. [Google Scholar] [CrossRef]
- Dua, T.K.; Giri, S.; Nandi, G.; Sahu, R.; Shaw, T.K.; Paul, P. Green Synthesis of Silver Nanoparticles Using Eupatorium Adenophorum Leaf Extract: Characterizations, Antioxidant, Antibacterial, and Photocatalytic Activities. Food Chem. Adv. 2023, 2, 100205. [Google Scholar] [CrossRef]
- Thomas, T.; Thalla, A.K. Synthesis of Silver Nanoparticles Using Myristica Fragrans Seed Shell: Assessment of Antibacterial, Antioxidant Properties and Photocatalytic Degradation of Dyes. J. Environ. Chem. Eng. 2023, 109585, 109585. [Google Scholar] [CrossRef]
- Shyamalagowri, S.; Charles, P.; Manjunathan, J.; Kamaraj, M.; Anitha, R.; Pugazhendhi, A. In Vitro Anticancer Activity of Silver Nanoparticles Phyto-Fabricated by Hylocereus Undatus Peel Extracts on Human Liver Carcinoma (HepG2) Cell Lines. Process Biochem. 2022, 116, 17–25. [Google Scholar] [CrossRef]
- Saxena, J.; Ayushi, K.M. Evaluation of Sclerotinia Sclerotiorum MTCC 8785 as a Biological Agent for the Synthesis of Silver Nanoparticles and Assessment of Their Antifungal Potential against Trichoderma Harzianum MTCC 801. Environ. Res. 2023, 216, 114752. [Google Scholar] [CrossRef] [PubMed]
- Shantkriti, S.; Pradeep, M.; Unish, K.K.; Das, V.; Nidhin, S.; Gugan, K.; Murugan, A. Bioynthesis of Silver Nanoparticles Using Dunaliella Salina and Its Antibacterial Applications. Appl. Surf. Sci. Adv. 2023, 13, 100377. [Google Scholar] [CrossRef]
- Mourad, R.; Helaly, F.; Darwesh, O.; Sawy, S. El Antimicrobial and Physicomechanical Natures of Silver Nanoparticles Incorporated into Silicone-Hydrogel Films. Contact Lens Anterior Eye 2019, 42, 325–333. [Google Scholar] [CrossRef]
- Mamdouh, S.; Mahmoud, A.; Samir, A.; Mobarak, M.; Mohamed, T. Using Femtosecond Laser Pulses to Investigate the Nonlinear Optical Properties of Silver Nanoparticles Colloids in Distilled Water Synthesized by Laser Ablation. Phys. B Condens. Matter 2022, 631, 413727. [Google Scholar] [CrossRef]
- Sportelli, M.C.; Clemente, M.; Izzi, M.; Volpe, A.; Ancona, A.; Picca, R.A.; Palazzo, G.; Cioffi, N. Exceptionally Stable Silver Nanoparticles Synthesized by Laser Ablation in Alcoholic Organic Solvent. Colloids Surf. A Physicochem. Eng. Asp. 2018, 559, 148–158. [Google Scholar] [CrossRef]
- Jeevanandam, J.; Krishnan, S.; Hii, Y.S.; Pan, S.; Chan, Y.S.; Acquah, C.; Danquah, M.K.; Rodrigues, J. Synthesis Approach-Dependent Antiviral Properties of Silver Nanoparticles and Nanocomposites. J. Nanostructure Chem. 2022, 12, 809–831. [Google Scholar] [CrossRef]
- Vigneswari, S.; Amelia, T.S.M.; Hazwan, M.H.; Mouriya, G.K.; Bhubalan, K.; Amirul, A.A.A.; Ramakrishna, S. Transformation of Biowaste for Medical Applications: Incorporation of Biologically Derived Silver Nanoparticles as Antimicrobial Coating. Antibiotics 2021, 10, 229. [Google Scholar] [CrossRef]
- Gahlawat, G.; Choudhury, A.R. A Review on the Biosynthesis of Metal and Metal Salt Nanoparticles by Microbes. RSC Adv. 2019, 9, 12944–12967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mondal, A.H.; Yadav, D.; Mitra, S.; Mukhopadhyay, K. Biosynthesis of Silver Nanoparticles Using Culture Supernatant of Shewanella Sp. ARY1 and Their Antibacterial Activity. Int. J. Nanomed. 2020, 15, 8295–8310. [Google Scholar] [CrossRef]
- Alfryyan, N.; Kordy, M.G.M.; Abdel-Gabbar, M.; Soliman, H.A.; Shaban, M. Characterization of the Biosynthesized Intracellular and Extracellular Plasmonic Silver Nanoparticles Using Bacillus Cereus and Their Catalytic Reduction of Methylene Blue. Sci. Rep. 2022, 12, 12495. [Google Scholar] [CrossRef] [PubMed]
- Hanna, A.L.; Hamouda, H.M.; Goda, H.A.; Sadik, M.W.; Moghanm, F.S.; Ghoneim, A.M.; Alenezi, M.A.; Alnomasy, S.F.; Alam, P.; Elsayed, T.R. Biosynthesis and Characterization of Silver Nanoparticles Produced by Phormidium Ambiguum and Desertifilum Tharense Cyanobacteria. Bioinorg. Chem. Appl. 2022, 2022, 9072508. [Google Scholar] [CrossRef] [PubMed]
- Cepoi, L.; Rudi, L.; Zinicovscaia, I.; Chiriac, T.; Miscu, V.; Rudic, V. Biochemical Changes in Microalga Porphyridium Cruentum Associated with Silver Nanoparticles Biosynthesis. Arch. Microbiol. 2021, 203, 1547–1554. [Google Scholar] [CrossRef]
- Silva, L.P.C.; Oliveira, J.P.; Keijok, W.J.; da Silva, A.R.; Aguiar, A.R.; Guimarães, M.C.C.; Ferraz, C.M.; Araújo, J.V.; Tobias, F.L.; Braga, F.R. Extracellular Biosynthesis of Silver Nanoparticles Using the Cell-Free Filtrate of Nematophagous Fungus Duddingtonia Flagrans. Int. J. Nanomed. 2017, 12, 6373–6381. [Google Scholar] [CrossRef] [Green Version]
- Saad, A.M.; El-Saadony, M.T.; El-Tahan, A.M.; Sayed, S.; Moustafa, M.A.M.; Taha, A.E.; Taha, T.F.; Ramadan, M.M. Polyphenolic Extracts from Pomegranate and Watermelon Wastes as Substrate to Fabricate Sustainable Silver Nanoparticles with Larvicidal Effect against Spodoptera Littoralis. Saudi J. Biol. Sci. 2021, 28, 5674–5683. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.; Liao, X.; Liu, X.; Li, W.; Huang, R.; Tang, J.; Xu, Q.; Li, X.; Yu, J. Characterization and Antimicrobial Activity of Silver Nanoparticles Synthesized with the Peel Extract of Mango. Materials 2021, 14, 5878. [Google Scholar] [CrossRef]
- Devi, N.; Rani, K.; Kharb, P.; Kaushik, P. Bio-Fabrication of Euryale Ferox (Makhana) Leaf Silver Nanoparticles and Their Antibacterial, Antioxidant and Cytotoxic Potential. Plants 2022, 11, 2766. [Google Scholar] [CrossRef]
- Navale, V.; Vamkudoth, K.R.; Ajmera, S.; Dhuri, V. Aspergillus Derived Mycotoxins in Food and the Environment: Prevalence, Detection, and Toxicity. Toxicol Rep. 2021, 8, 1008–1030. [Google Scholar] [CrossRef] [PubMed]
- Chuaysrinule, C.; Maneeboon, T.; Roopkham, C.; Mahakarnchanakul, W. Occurrence of Aflatoxin- and Ochratoxin A-Producing Aspergillus Species in Thai Dried Chilli. J. Agric. Food Res. 2020, 2, 100054. [Google Scholar] [CrossRef]
- Massomo, S.M.S. Aspergillus Flavus and Aflatoxin Contamination in the Maize Value Chain and What Needs to Be Done in Tanzania. Sci. Afr. 2020, 10, e00606. [Google Scholar] [CrossRef]
- Kumar, P.; Mahato, D.K.; Sharma, B.; Borah, R.; Haque, S.; Mahmud, M.M.C.; Shah, A.K.; Rawal, D.; Bora, H.; Bui, S. Ochratoxins in Food and Feed: Occurrence and Its Impact on Human Health and Management Strategies. Toxicon 2020, 187, 151–162. [Google Scholar] [CrossRef]
- Ma, L.; Liu, J.; Su, W.; Zeng, X.; Liu, X.; Li, W.; Deng, J.; Tang, J. Extracellular Biosynthesis, Characterization and Cytotoxic Effect of Silver Nanoparticles by Streptomyces Coelicoflavus KS-3. J. Nanosci. Nanotechnol. 2018, 18, 8133–8141. [Google Scholar] [CrossRef]
- Bakhtiari-Sardari, A.; Mashreghi, M.; Eshghi, H.; Behnam-Rasouli, F.; Lashani, E.; Shahnavaz, B. Comparative Evaluation of Silver Nanoparticles Biosynthesis by Two Cold-Tolerant Streptomyces Strains and Their Biological Activities. Biotechnol. Lett. 2020, 42, 1985–1999. [Google Scholar] [CrossRef]
- Al-Dhabi, N.A.; Ghilan, A.K.M.; Arasu, M.V.; Duraipandiyan, V. Green Biosynthesis of Silver Nanoparticles Produced from Marine Streptomyces Sp. Al-Dhabi-89 and Their Potential Applications against Wound Infection and Drug Resistant Clinical Pathogens. J. Photochem. Photobiol. B 2018, 189, 176–184. [Google Scholar] [CrossRef]
- Elsilk, S.E.; Khalil, M.A.; Aboshady, T.A.; Alsalmi, F.A.; Ali, S.S. Streptomyces Rochei MS-37 as a Novel Marine Actinobacterium for Green Biosynthesis of Silver Nanoparticles and Their Biomedical Applications. Molecules 2022, 27, 7296. [Google Scholar] [CrossRef] [PubMed]
- Abd-Elhady, H.M.; Ashor, M.A.; Hazem, A.; Saleh, F.M.; Selim, S.; el Nahhas, N.; Abdel-Hafez, S.H.; Sayed, S.; Hassan, E.A. Biosynthesis and Characterization of Extracellular Silver Nanoparticles from Streptomyces Aizuneusis: Antimicrobial, Anti Larval, and Anticancer Activities. Molecules 2021, 27, 212. [Google Scholar] [CrossRef] [PubMed]
- Zwar, I.P.; Trotta, C.D.V.; Ziotti, A.B.S.; Lima Neto, M.; Araújo, W.L.; de Melo, I.S.; Ottoni, C.A.; de Souza, A.O. Biosynthesis of Silver Nanoparticles Using Actinomycetes, Phytotoxicity on Rice Seeds, and Potential Application in the Biocontrol of Phytopathogens. J. Basic Microbiol. 2023, 63, 64–74. [Google Scholar] [CrossRef]
- Elnady, A.; Sorour, N.M.; Abbas, R.N. Characterization, Cytotoxicity, and Genotoxicity Properties of Novel Biomediated Nanosized-Silver by Egyptian Streptomyces Roseolus for Safe Antimicrobial Applications. World J. Microbiol. Biotechnol. 2022, 38, 47. [Google Scholar] [CrossRef]
- Mechouche, M.S.; Merouane, F.; Messaad, C.E.H.; Golzadeh, N.; Vasseghian, Y.; Berkani, M. Biosynthesis, Characterization, and Evaluation of Antibacterial and Photocatalytic Methylene Blue Dye Degradation Activities of Silver Nanoparticles from Streptomyces Tuirus Strain. Environ. Res. 2022, 204, 112360. [Google Scholar] [CrossRef]
- Hayakawa, M.; Nonomura, H. Humic Acid-Vitamin Agar, a New Medium for the Selective Isolation of Soil Actinomycetes. J. Ferment. Technol. 1987, 65, 501–509. [Google Scholar] [CrossRef]
- Kamel, Z.; Mohamed, N.M.; Farahat, M.G. Optimization of Culture Conditions for Production of B-Galactosidase by Bacillus Megaterium NM56 Isolated from Raw Milk. Res. J. Pharm. Biol. Chem. Sci. 2016, 7, 366–376. [Google Scholar]
- Yoon, S.H.; Ha, S.M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A Taxonomically United Database of 16S RRNA Gene Sequences and Whole-Genome Assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef] [PubMed]
- Khalil, N.M.; Abd El-Ghany, M.N.; Rodríguez-Couto, S. Antifungal and Anti-Mycotoxin Efficacy of Biogenic Silver Nanoparticles Produced by Fusarium Chlamydosporum and Penicillium Chrysogenum at Non-Cytotoxic Doses. Chemosphere 2019, 218, 477–486. [Google Scholar] [CrossRef]
- Davis, N.D.; Diener, U.L.; Eldridge, D.W. Production of Aflatoxins B1 and G1 by Aspergillus Flavus in a Semisynthetic Medium. Appl. Microbiol. 1966, 14, 378–380. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Ghany, M.N.; Hamdi, S.A.; Korany, S.M.; Elbaz, R.M.; Farahat, M.G. Biosynthesis of Novel Tellurium Nanorods by Gayadomonas Sp. TNPM15 Isolated from Mangrove Sediments and Assessment of Their Impact on Spore Germination and Ultrastructure of Phytopathogenic Fungi. Microorganisms 2023, 11, 558. [Google Scholar] [CrossRef] [PubMed]
- Qin, S.; Li, J.; Chen, H.H.; Zhao, G.Z.; Zhu, W.Y.; Jiang, C.L.; Xu, L.H.; Li, W.J. Isolation, Diversity, and Antimicrobial Activity of Rare Actinobacteria from Medicinal Plants of Tropical Rain Forests in Xishuangbanna China. Appl. Environ. Microbiol. 2009, 75, 6176–6186. [Google Scholar] [CrossRef] [Green Version]
- Subramani, R.; Aalbersberg, W. Culturable Rare Actinomycetes: Diversity, Isolation and Marine Natural Product Discovery. Appl. Microbiol. Biotechnol. 2013, 97, 9291–9321. [Google Scholar] [CrossRef]
- Singh, V.; Haque, S.; Singh, H.; Verma, J.; Vibha, K.; Singh, R.; Jawed, A.; Tripathi, C.K.M. Isolation, Screening, and Identification of Novel Isolates of Actinomycetes from India for Antimicrobial Applications. Front. Microbiol. 2016, 7, 1921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niyomvong, N.; Pathom-Aree, W.; Thamchaipenet, A.; Duangmal, K. Actinomycetes from Tropical Limestone Caves. Chiang Mai J. Sci. 2012, 39, 373–388. [Google Scholar]
- González Henao, S.; Ghneim-Herrera, T. Heavy Metals in Soils and the Remediation Potential of Bacteria Associated with the Plant Microbiome. Front. Environ. Sci. 2021, 9, 15. [Google Scholar] [CrossRef]
- Hao, X.; Zhu, J.; Rensing, C.; Liu, Y.; Gao, S.; Chen, W.; Huang, Q.; Liu, Y.R. Recent Advances in Exploring the Heavy Metal(Loid) Resistant Microbiome. Comput. Struct. Biotechnol. J. 2021, 19, 94–109. [Google Scholar] [CrossRef]
- Fadl, M.G.; Farahat, M.G.; Mohamed, Z.K. Optimum Biosorption and Resistance of Uranium by Metal-Resistant Bacteria Isolated from Rock Ore. Geomicrobiol. J. 2022, 39, 689–696. [Google Scholar] [CrossRef]
- Duraisamy, P.; Sekar, J.; Arunkumar, A.D.; Ramalingam, P.V. Kinetics of Phenol Biodegradation by Heavy Metal Tolerant Rhizobacteria Glutamicibacter Nicotianae MSSRFPD35 from Distillery Effluent Contaminated Soils. Front. Microbiol. 2020, 11, 1573. [Google Scholar] [CrossRef]
- Sturm, G.; Brunner, S.; Suvorova, E.; Dempwolff, F.; Reiner, J.; Graumann, P.; Bernier-Latmani, R.; Majzlan, J.; Gescher, J. Chromate Resistance Mechanisms in Leucobacter Chromiiresistens. Appl. Environ. Microbiol. 2018, 84, 02208–02218. [Google Scholar] [CrossRef] [Green Version]
- Tahri Joutey, N.; Bahafid, W.; Sayel, H.; Nassef, S.; El Ghachtouli, N. Leucobacter Chromiireducens CRB2, a New Strain with High Cr(VI) Reduction Potential Isolated from Tannery-Contaminated Soil (Fez, Morocco). Ann. Microbiol. 2016, 66, 425–436. [Google Scholar] [CrossRef]
- Hyun, D.W.; Sung, H.; Kim, P.S.; Yun, J.H.; Bae, J.W. Leucobacter Coleopterorum Sp. Nov., Leucobacter Insecticola Sp. Nov., and Leucobacter Viscericola Sp. Nov., Isolated from the Intestine of the Diving Beetles, Cybister Brevis and Cybister Lewisianus, and Emended Description of the Genus Leucobacter. J. Microbiol. 2021, 59, 360–368. [Google Scholar] [CrossRef]
- Khalil, M.A.; El-Shanshoury, A.E.R.R.; Alghamdi, M.A.; Alsalmi, F.A.; Mohamed, S.F.; Sun, J.; Ali, S.S. Biosynthesis of Silver Nanoparticles by Marine Actinobacterium Nocardiopsis Dassonvillei and Exploring Their Therapeutic Potentials. Front. Microbiol. 2022, 12, 4117. [Google Scholar] [CrossRef] [PubMed]
- Bahrulolum, H.; Nooraei, S.; Javanshir, N.; Tarrahimofrad, H.; Mirbagheri, V.S.; Easton, A.J.; Ahmadian, G. Green Synthesis of Metal Nanoparticles Using Microorganisms and Their Application in the Agrifood Sector. J. Nanobiotechnol. 2021, 19, 1–26. [Google Scholar] [CrossRef]
- Roy, A.; Bulut, O.; Some, S.; Mandal, A.K.; Yilmaz, M.D. Green Synthesis of Silver Nanoparticles: Biomolecule-Nanoparticle Organizations Targeting Antimicrobial Activity. RSC Adv. 2019, 9, 2673–2702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hietzschold, S.; Walter, A.; Davis, C.; Taylor, A.A.; Sepunaru, L. Does Nitrate Reductase Play a Role in Silver Nanoparticle Synthesis? Evidence for NADPH as the Sole Reducing Agent. ACS Sustain. Chem. Eng. 2019, 7, 8070–8076. [Google Scholar] [CrossRef]
- Tag, H.M.; Saddiq, A.A.; Alkinani, M.; Hagagy, N. Biosynthesis of Silver Nanoparticles Using Haloferax Sp. NRS1: Image Analysis, Characterization, In Vitro Thrombolysis and Cytotoxicity. AMB Express 2021, 11, 75. [Google Scholar] [CrossRef]
- Alam, A.; Tanveer, F.; Khalil, A.T.; Zohra, T.; Khamlich, S.; Alam, M.M.; Salman, M.; Ali, M.; Ikram, A.; Shinwari, Z.K.; et al. Silver Nanoparticles Biosynthesized from Secondary Metabolite Producing Marine Actinobacteria and Evaluation of Their Biomedical Potential. Antonie Van Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2021, 114, 1497–1516. [Google Scholar] [CrossRef]
- El-Bendary, M.A.; Afifi, S.S.; Moharam, M.E.; Abo El-Ola, S.M.; Salama, A.; Omara, E.A.; Shaheen, M.N.F.; Hamed, A.A.; Gawdat, N.A. Biosynthesis of Silver Nanoparticles Using Isolated Bacillus Subtilis: Characterization, Antimicrobial Activity, Cytotoxicity, and Their Performance as Antimicrobial Agent for Textile Materials. Prep. Biochem. Biotechnol. 2020, 51, 54–68. [Google Scholar] [CrossRef]
- Huq, M.A.; Akter, S. Biosynthesis, Characterization and Antibacterial Application of Novel Silver Nanoparticles against Drug Resistant Pathogenic Klebsiella Pneumoniae and Salmonella Enteritidis. Molecules 2021, 26, 5996. [Google Scholar] [CrossRef]
- Talabani, R.F.; Hamad, S.M.; Barzinjy, A.A.; Demir, U. Biosynthesis of Silver Nanoparticles and Their Applications in Harvesting Sunlight for Solar Thermal Generation. Nanomaterials 2021, 11, 2421. [Google Scholar] [CrossRef]
- Wang, X.; Lee, S.Y.; Akter, S.; Huq, M.A. Probiotic-Mediated Biosynthesis of Silver Nanoparticles and Their Antibacterial Applications against Pathogenic Strains of Escherichia Coli O157:H7. Polymers 2022, 14, 1834. [Google Scholar] [CrossRef] [PubMed]
- Nagaraja, S.K.; Niazi, S.K.; Bepari, A.; Assiri, R.A.; Nayaka, S. Leonotis Nepetifolia Flower Bud Extract Mediated Green Synthesis of Silver Nanoparticles, Their Characterization, and In Vitro Evaluation of Biological Applications. Materials 2022, 15, 8990. [Google Scholar] [CrossRef] [PubMed]
- Malik, M.; Iqbal, M.A.; Malik, M.; Raza, M.A.; Shahid, W.; Choi, J.R.; Pham, P.V. Biosynthesis and Characterizations of Silver Nanoparticles from Annona Squamosa Leaf and Fruit Extracts for Size-Dependent Biomedical Applications. Nanomaterials 2022, 12, 616. [Google Scholar] [CrossRef] [PubMed]
- Mahmoudi, F.; Mahmoudi, F.; Gollo, K.H.; Amini, M.M. Biosynthesis of Novel Silver Nanoparticles Using Eryngium Thyrsoideum Boiss Extract and Comparison of Their Antidiabetic Activity with Chemical Synthesized Silver Nanoparticles in Diabetic Rats. Biol. Trace Elem. Res. 2021, 199, 1967–1978. [Google Scholar] [CrossRef]
- Bold, B.E.; Urnukhsaikhan, E.; Mishig-Ochir, T. Biosynthesis of Silver Nanoparticles with Antibacterial, Antioxidant, Anti-Inflammatory Properties and Their Burn Wound Healing Efficacy. Front. Chem. 2022, 10, 1022. [Google Scholar] [CrossRef]
- Weng, X.; Yang, K.; Owens, G.; Chen, Z. Biosynthesis of Silver Nanoparticles Using Three Different Fruit Extracts: Characterization, Formation Mechanism and Estrogen Removal. J. Environ. Manag. 2022, 316, 115224. [Google Scholar] [CrossRef]
- Alhomaidi, E.; Jasim, S.A.; Amin, H.I.M.; Lima Nobre, M.A.; Khatami, M.; Jalil, A.T.; Hussain Dilfy, S. Biosynthesis of Silver Nanoparticles Using Lawsonia Inermis and Their Biomedical Application. IET Nanobiotechnol. 2022, 16, 284–294. [Google Scholar] [CrossRef]
- Ogunleye, G.E.; Adebayo-Tayo, B.C.; Oyinlola, K.A. Biological Evaluation of Extracellular Mycosynthesized Silver Nanoparticles by Trichoderma Asperellum. BioMetals 2022, 36, 97–109. [Google Scholar] [CrossRef]
- Bhadani, R.V.; Gajera, H.P.; Hirpara, D.G.; Savaliya, D.D.; Anuj, S.A. Biosynthesis and Characterization of Extracellular Metabolites-Based Nanoparticles to Control the Whitefly. Arch. Microbiol. 2022, 204, 311. [Google Scholar] [CrossRef]
- Li, P.-J.; Pan, J.-J.; Tao, L.-J.; Li, X.; Su, D.-L.; Shan, Y.; Li, H.-Y.; Li, P.-J.; Pan, J.-J.; Tao, L.-J.; et al. Green Synthesis of Silver Nanoparticles by Extracellular Extracts from Aspergillus Japonicus PJ01. Molecules 2021, 26, 4479. [Google Scholar] [CrossRef]
- Abdelmoneim, H.M.; Taha, T.H.; Elnouby, M.S.; AbuShady, H.M. Extracellular Biosynthesis, OVAT/Statistical Optimization, and Characterization of Silver Nanoparticles (AgNPs) Using Leclercia Adecarboxylata THHM and Its Antimicrobial Activity. Microb. Cell Factories 2022, 21, 277. [Google Scholar] [CrossRef] [PubMed]
- Murugaiah, H.; Teh, C.L.; Loh, K.C.; Mohamad Yahya, A.R.; Md Noh, N.A.; Abu Bakar, N.H.H.; Kernain, D.; Hashim, R.; Bustami, Y. Study of Antibacterial and Anticancer Properties of BioAgNPs Synthesized Using Streptomyces Sp. PBD-311B and the Application of BioAgNP-CNC/Alg as an Antibacterial Hydrogel Film against P. Aeruginosa USM-AR2 and MRSA. Molecules 2021, 26, 6414. [Google Scholar] [CrossRef] [PubMed]
- Gurunathan, S.; Kalishwaralal, K.; Vaidyanathan, R.; Venkataraman, D.; Pandian, S.R.K.; Muniyandi, J.; Hariharan, N.; Eom, S.H. Biosynthesis, Purification and Characterization of Silver Nanoparticles Using Escherichia coli. Colloids Surf. B Biointerfaces 2009, 74, 328–335. [Google Scholar] [CrossRef] [PubMed]
- Manikprabhu, D.; Cheng, J.; Chen, W.; Sunkara, A.K.; Mane, S.B.; Kumar, R.; Das, M.; Hozzein, W.N.; Duan, Y.Q.; Li, W.J. Sunlight Mediated Synthesis of Silver Nanoparticles by a Novel Actinobacterium (Sinomonas Mesophila MPKL 26) and Its Antimicrobial Activity against Multi Drug Resistant Staphylococcus Aureus. J. Photochem. Photobiol. B 2016, 158, 202–205. [Google Scholar] [CrossRef]
- Arshad, H.; Sadaf, S.; Hassan, U. De-Novo Fabrication of Sunlight Irradiated Silver Nanoparticles and Their Efficacy against E. Coli and S. Epidermidis. Sci. Rep. 2022, 12, 676. [Google Scholar] [CrossRef]
- Satheesh, V.; Mohamed, J.M.M.; El-Sherbiny, M.; Othman, G.; Al-Serwi, R.H.; Thilagar, S. Sunlight-Assisted Green Synthesis of Silver Nanoparticles Using Zizania Latifolia Extract: Toward Antimicrobial Applications. Biomass Convers Biorefin. 2022, 12, 1–11. [Google Scholar] [CrossRef]
- Nallal, V.U.M.; Prabha, K.; VethaPotheher, I.; Ravindran, B.; Baazeem, A.; Chang, S.W.; Otunola, G.A.; Razia, M. Sunlight-Driven Rapid and Facile Synthesis of Silver Nanoparticles Using Allium Ampeloprasum Extract with Enhanced Antioxidant and Antifungal Activity. Saudi J. Biol. Sci. 2021, 28, 3660–3668. [Google Scholar] [CrossRef]
- Patil, H.V.; Nithin, K.S.; Sachhidananda, S.; Chandrashekara, K.T.; Kumar, S. Mycofabrication of Bioactive Silver Nanoparticle: Photo Catalysed Synthesis and Characterization to Attest Its Augmented Bio-Efficacy. Arab. J. Chem. 2019, 12, 4596–4611. [Google Scholar] [CrossRef] [Green Version]
- Sonbol, H.; Mohammed, A.E.; Korany, S.M. Soil Fungi as Biomediator in Silver Nanoparticles Formation and Antimicrobial Efficacy. Int. J. Nanomed. 2022, 17, 2843–2863. [Google Scholar] [CrossRef]
- Bhatnagar, S.; Ogbonna, C.N.; Ogbonna, J.C.; Aoyagi, H. Effect of Physicochemical Factors on Extracellular Fungal Pigment-Mediated Biofabrication of Silver Nanoparticles. Green Chem. Lett. Rev. 2022, 15, 274–284. [Google Scholar] [CrossRef]
- Baltussen, T.J.H.; Zoll, J.; Verweij, P.E.; Melchers, W.J.G. Molecular Mechanisms of Conidial Germination in Aspergillus Spp. Microbiol. Mol. Biol. Rev. 2020, 84, e00049-e19. [Google Scholar] [CrossRef] [PubMed]
- Osherov, N.; May, G.S. The Molecular Mechanisms of Conidial Germination. FEMS Microbiol. Lett. 2001, 199, 153–160. [Google Scholar] [CrossRef]
- Zhou, L.; Zhao, X.; Li, M.; Lu, Y.; Ai, C.; Jiang, C.; Liu, Y.; Pan, Z.; Shi, J. Antifungal Activity of Silver Nanoparticles Synthesized by Iturin against Candida Albicans in Vitro and in Vivo. Appl. Microbiol. Biotechnol. 2021, 105, 3759–3770. [Google Scholar] [CrossRef]
- Li, L.; Pan, H.; Deng, L.; Qian, G.; Wang, Z.; Li, W.; Zhong, C. The Antifungal Activity and Mechanism of Silver Nanoparticles against Four Pathogens Causing Kiwifruit Post-Harvest Rot. Front. Microbiol. 2022, 13, 3418. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Singh, S.; Gaikwad, S.; Nawani, N.; Junnarkar, M.; Pawar, S.V. Optimization of Process Parameters for the Synthesis of Silver Nanoparticles from Piper Betle Leaf Aqueous Extract, and Evaluation of Their Antiphytofungal Activity. Environ. Sci. Pollut. Res. 2020, 27, 27221–27233. [Google Scholar] [CrossRef] [PubMed]
- Gajera, H.P.; Hirpara, D.G.; Bhadani, R.V.; Golakiya, B.A. Green Synthesis and Characterization of Nanosilver Derived from Extracellular Metabolites of Potent Bacillus Subtilis for Antifungal and Eco-Friendly Action against Phytopathogen. BioMetals 2022, 35, 479–497. [Google Scholar] [CrossRef]
- Ibrahim, E.; Zhang, M.; Zhang, Y.; Hossain, A.; Qiu, W.; Chen, Y.; Wang, Y.; Wu, W.; Sun, G.; Li, B. Green-Synthesization of Silver Nanoparticles Using Endophytic Bacteria Isolated from Garlic and Its Antifungal Activity against Wheat Fusarium Head Blight Pathogen Fusarium Graminearum. Nanomaterials 2020, 10, 219. [Google Scholar] [CrossRef] [Green Version]
- Darwesh, O.M.; Elshahawy, I.E. Silver Nanoparticles Inactivate Sclerotial Formation in Controlling White Rot Disease in Onion and Garlic Caused by the Soil Borne Fungus Stromatinia Cepivora. Eur. J. Plant Pathol. 2021, 160, 917–934. [Google Scholar] [CrossRef]
- Darwesh, O.M.; Barakat, K.M.; Mattar, M.Z.; Sabae, S.Z.; Hassan, S.H. Production of Antimicrobial Blue Green Pigment Pyocyanin by Marine Pseudomonas Aeruginosa. Biointerface Res. Appl. Chem. 2019, 9, 4334–4339. [Google Scholar] [CrossRef]
- Ameen, F.; Al-Homaidan, A.A.; Al-Sabri, A.; Almansob, A.; AlNAdhari, S. Anti-Oxidant, Anti-Fungal and Cytotoxic Effects of Silver Nanoparticles Synthesized Using Marine Fungus Cladosporium Halotolerans. Appl. Nanosci. 2021, 13, 623–631. [Google Scholar] [CrossRef]
- Mansoor, S.; Zahoor, I.; Baba, T.R.; Padder, S.A.; Bhat, Z.A.; Koul, A.M.; Jiang, L. Fabrication of Silver Nanoparticles against Fungal Pathogens. Front. Nanotechnol. 2021, 3, 679358. [Google Scholar] [CrossRef]
- Mussin, J.; Giusiano, G. Biogenic Silver Nanoparticles as Antifungal Agents. Front. Chem. 2022, 10, 1270. [Google Scholar] [CrossRef] [PubMed]
- Barros, D.; Pradhan, A.; Pascoal, C.; Cássio, F. Transcriptomics Reveals the Action Mechanisms and Cellular Targets of Citrate-Coated Silver Nanoparticles in a Ubiquitous Aquatic Fungus. Environ. Pollut. 2021, 268, 115913. [Google Scholar] [CrossRef]
- Sultana, T.; Javed, B.; Raja, N.I.; Mashwani, Z.U.R. Silver Nanoparticles Elicited Physiological, Biochemical, and Antioxidant Modifications in Rice Plants to Control Aspergillus Flavus. Green Process. Synth. 2021, 10, 314–324. [Google Scholar] [CrossRef]
- Fouad, F.A.H.; Youssef, D.G.; Shahat, F.M.; Abd El-Ghany, M.N. Role of Microorganisms in Biodegradation of Pollutants. In Handbook of Biodegradable Materials; Springer: Berlin/Heidelberg, Germany, 2023; pp. 221–260. [Google Scholar] [CrossRef]
- Nasirian, V.; Chabok, A.; Barati, A.; Rafienia, M.; Arabi, M.S.; Shamsipur, M. Ultrasensitive Aflatoxin B1 Assay Based on FRET from Aptamer Labelled Fluorescent Polymer Dots to Silver Nanoparticles Labeled with Complementary DNA. Microchim. Acta 2017, 184, 4655–4662. [Google Scholar] [CrossRef]
- Dizaj, S.M.; Lotfipour, F.; Barzegar-Jalali, M.; Zarrintan, M.H.; Adibkia, K. Antimicrobial Activity of the Metals and Metal Oxide Nanoparticles. Mater. Sci. Eng. C 2014, 44, 278–284. [Google Scholar] [CrossRef]
- Cao, J.; Feng, Y.; He, S.; Lin, X. Silver Nanoparticles Deteriorate the Mutual Interaction between Maize (Zea mays L.) and Arbuscular Mycorrhizal Fungi: A Soil Microcosm Study. Appl. Soil Ecol. 2017, 119, 307–316. [Google Scholar] [CrossRef]
- Ammar, H.A.M.; El-Desouky, T.A. Green Synthesis of Nanosilver Particles by Aspergillus Terreus HA1N and Penicillium Expansum HA2N and Its Antifungal Activity against Mycotoxigenic Fungi. J. Appl. Microbiol. 2016, 121, 89–100. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Wang, F.; Yalamarty, S.S.K.; Filipczak, N.; Jin, Y.; Li, X. Nano Silver-Induced Toxicity and Associated Mechanisms. Int. J. Nanomed. 2022, 17, 1851–1864. [Google Scholar] [CrossRef]
- Noga, M.; Milan, J.; Frydrych, A.; Jurowski, K. Toxicological Aspects, Safety Assessment, and Green Toxicology of Silver Nanoparticles (AgNPs)—Critical Review: State of the Art. Int. J. Mol. Sci. 2023, 24, 5133. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, C.A.; Seckler, M.M.; Ingle, A.P.; Gupta, I.; Galdiero, S.; Galdiero, M.; Gade, A.; Rai, M. Silver Nanoparticles: Therapeutical Uses, Toxicity, and Safety Issues. J. Pharm. Sci. 2014, 103, 1931–1944. [Google Scholar] [CrossRef] [PubMed]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abd El-Ghany, M.N.; Hamdi, S.A.; Korany, S.M.; Elbaz, R.M.; Emam, A.N.; Farahat, M.G. Biogenic Silver Nanoparticles Produced by Soil Rare Actinomycetes and Their Significant Effect on Aspergillus-derived mycotoxins. Microorganisms 2023, 11, 1006. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms11041006
Abd El-Ghany MN, Hamdi SA, Korany SM, Elbaz RM, Emam AN, Farahat MG. Biogenic Silver Nanoparticles Produced by Soil Rare Actinomycetes and Their Significant Effect on Aspergillus-derived mycotoxins. Microorganisms. 2023; 11(4):1006. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms11041006
Chicago/Turabian StyleAbd El-Ghany, Mohamed N., Salwa A. Hamdi, Shereen M. Korany, Reham M. Elbaz, Ahmed N. Emam, and Mohamed G. Farahat. 2023. "Biogenic Silver Nanoparticles Produced by Soil Rare Actinomycetes and Their Significant Effect on Aspergillus-derived mycotoxins" Microorganisms 11, no. 4: 1006. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms11041006