
Persistent Dysbiosis, Parasite Rise and Growth Impairment in Aquacultured European Seabass after Oxytetracycline Treatment
 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Aquaculture Setup
2.2. Sampling
2.3. DNA Extraction and rRNA Gene Amplicon Sequencing
2.4. Data Processing
2.5. Calculation of Growth Performance
3. Results
3.1. Primary Data Analysis
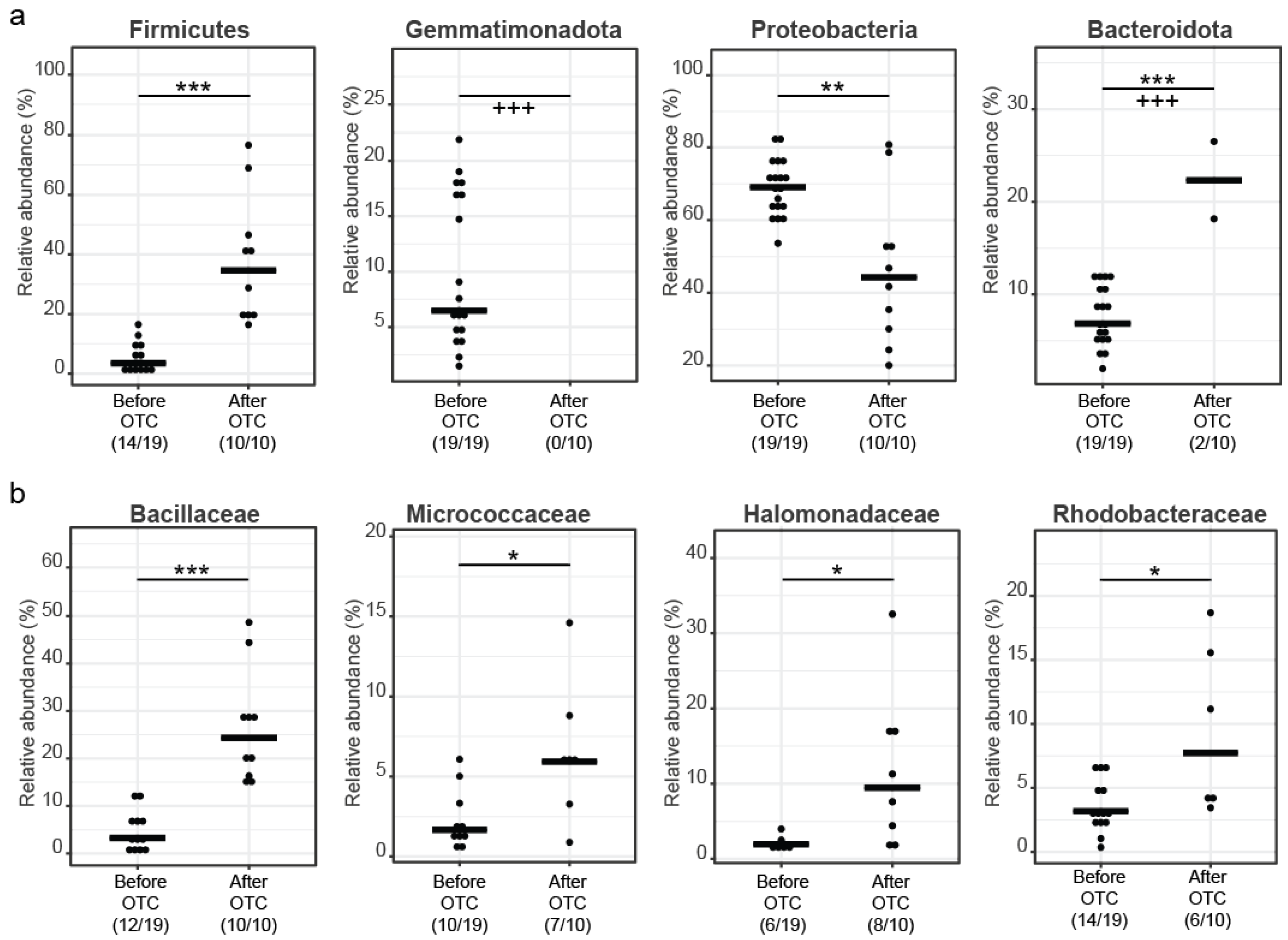
3.2. Microbial Diversity Analysis and Community Profiling
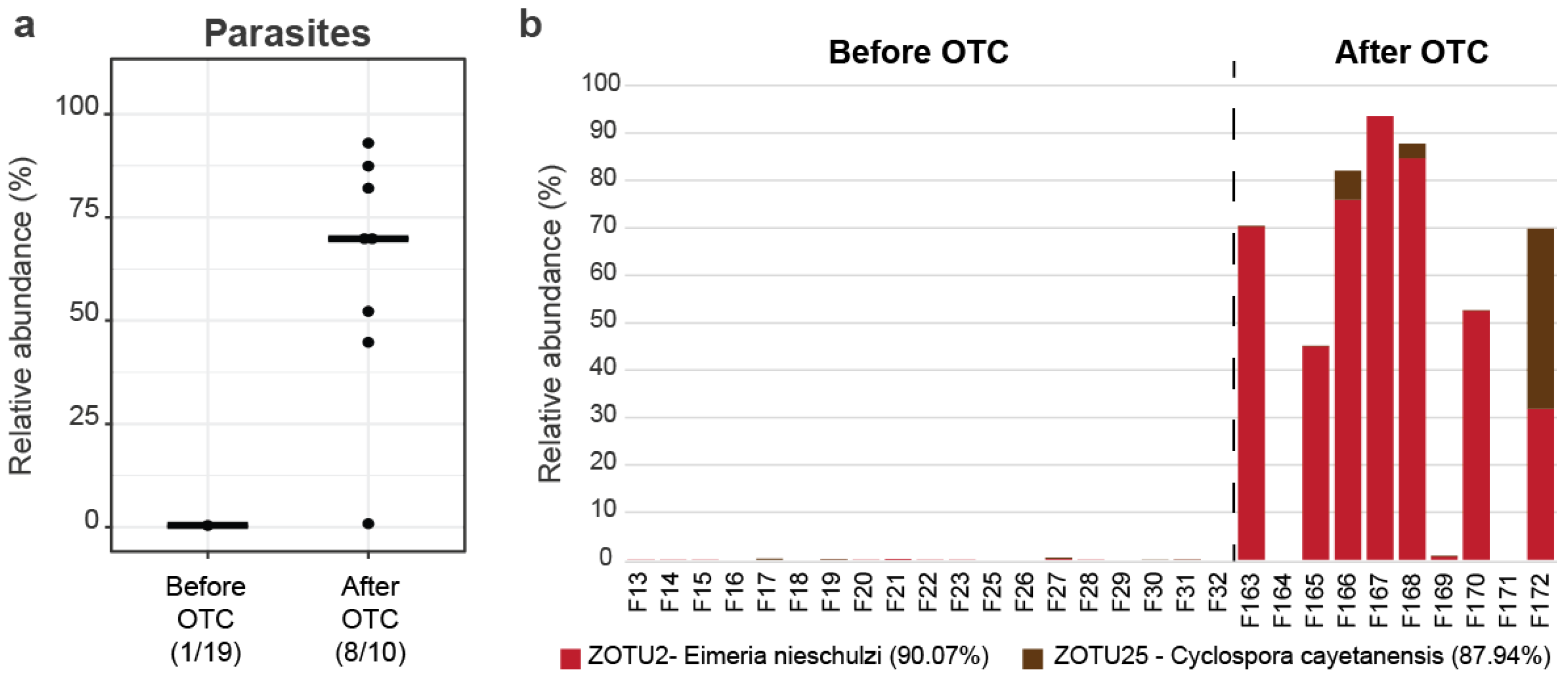
3.3. Apicomplexans Presence
3.4. Growth under OTC Treatment
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture 2020: Sustainability in Action; The State of World Fisheries and Aquaculture (SOFIA); FAO: Rome, Italy, 2020; ISBN 978-92-5-132692-3. [Google Scholar]
- European Commission; Directorate General for Maritime Affairs and Fisheries. The EU Fish Market: 2021 Edition; European Commission: Brussels, Belgium, 2021; ISBN 978-92-76-28905-0. [Google Scholar]
- Vandeputte, M.; Gagnaire, P.-A.; Allal, F. The European Sea Bass: A Key Marine Fish Model in the Wild and in Aquaculture. Anim. Genet. 2019, 50, 195–206. [Google Scholar] [CrossRef]
- Egerton, S.; Culloty, S.; Whooley, J.; Stanton, C.; Ross, R.P. The Gut Microbiota of Marine Fish. Front. Microbiol. 2018, 9, 873. [Google Scholar] [CrossRef]
- Llewellyn, M.S.; Boutin, S.; Hoseinifar, S.H.; Derome, N. Teleost Microbiomes: The State of the Art in Their Characterization, Manipulation and Importance in Aquaculture and Fisheries. Front. Microbiol. 2014, 5, 207. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.R.; Ran, C.; Ringø, E.; Zhou, Z.G. Progress in Fish Gastrointestinal Microbiota Research. Rev. Aquac. 2018, 10, 626–640. [Google Scholar] [CrossRef]
- Sullam, K.E.; Essinger, S.D.; Lozupone, C.A.; O’Connor, M.P.; Rosen, G.L.; Knight, R.; Kilham, S.S.; Russell, J.A. Environmental and Ecological Factors That Shape the Gut Bacterial Communities of Fish: A Meta-Analysis. Mol. Ecol. 2012, 21, 3363–3378. [Google Scholar] [CrossRef] [PubMed]
- Dehler, C.E.; Secombes, C.J.; Martin, S.A.M. Environmental and Physiological Factors Shape the Gut Microbiota of Atlantic Salmon Parr (Salmo Salar L.). Aquaculture 2017, 467, 149–157. [Google Scholar] [CrossRef]
- Walter, J.; Ley, R. The Human Gut Microbiome: Ecology and Recent Evolutionary Changes. Annu. Rev. Microbiol. 2011, 65, 411–429. [Google Scholar] [CrossRef]
- Alberdi, A.; Aizpurua, O.; Bohmann, K.; Zepeda-Mendoza, M.L.; Gilbert, M.T.P. Do Vertebrate Gut Metagenomes Confer Rapid Ecological Adaptation? Trends Ecol. Evol. 2016, 31, 689–699. [Google Scholar] [CrossRef]
- Rimoldi, S.; Terova, G.; Ascione, C.; Giannico, R.; Brambilla, F. Next Generation Sequencing for Gut Microbiome Characterization in Rainbow Trout (Oncorhynchus Mykiss) Fed Animal by-Product Meals as an Alternative to Fishmeal Protein Sources. PLoS ONE 2018, 13, e0193652. [Google Scholar] [CrossRef]
- Payne, C.J.; Turnbull, J.F.; MacKenzie, S.; Crumlish, M. The Effect of Oxytetracycline Treatment on the Gut Microbiome Community Dynamics in Rainbow Trout (Oncorhynchus Mykiss) over Time. Aquaculture 2022, 560, 738559. [Google Scholar] [CrossRef]
- Kamada, N.; Chen, G.Y.; Inohara, N.; Núñez, G. Control of Pathogens and Pathobionts by the Gut Microbiota. Nat. Immunol. 2013, 14, 685–690. [Google Scholar] [CrossRef] [PubMed]
- Brugman, S.; Nieuwenhuis, E.E.S. Mucosal Control of the Intestinal Microbial Community. J. Mol. Med. 2010, 88, 881–888. [Google Scholar] [CrossRef]
- Cerf-Bensussan, N.; Gaboriau-Routhiau, V. The Immune System and the Gut Microbiota: Friends or Foes? Nat. Rev. Immunol. 2010, 10, 735–744. [Google Scholar] [CrossRef]
- Viney, M.; Riley, E. From Immunology to Eco-Immunology: More than a New Name. In Eco-Immunology: Evolutive Aspects and Future Perspectives; Springer: Dordrecht, The Netherlands, 2014; pp. 1–19. ISBN 978-94-017-8711-6. [Google Scholar]
- Luan, Y.; Li, M.; Zhou, W.; Yao, Y.; Yang, Y.; Zhang, Z.; Ringø, E.; Erik Olsen, R.; Liu Clarke, J.; Xie, S.; et al. The Fish Microbiota: Research Progress and Potential Applications. Engineering 2023. [Google Scholar] [CrossRef]
- Reid, G.K.; Gurney-Smith, H.J.; Marcogliese, D.J.; Knowler, D.; Benfey, T.; Garber, A.F.; Forster, I.; Chopin, T.; Brewer-Dalton, K.; Moccia, R.D.; et al. Climate Change and Aquaculture: Considering Biological Response and Resources. Aquac. Environ. Interact. 2019, 11, 569–602. [Google Scholar] [CrossRef]
- Nguyen, F.; Starosta, A.L.; Arenz, S.; Sohmen, D.; Dönhöfer, A.; Wilson, D.N. Tetracycline Antibiotics and Resistance Mechanisms. Biol. Chem. 2014, 395, 559–575. [Google Scholar] [CrossRef]
- Sversut, R.A.; da Silva, A.A.; Cardoso, T.F.M.; Kassab, N.M.; do Amaral, M.S.; Salgado, H.R.N. A Critical Review of Properties and Analytical Methods for the Determination of Oxytetracyline in Biological and Pharmaceutical Matrices. Crit. Rev. Anal. Chem. 2017, 47, 154–171. [Google Scholar] [CrossRef]
- Rigos, G.; Kogiannou, D.; Padrós, F.; Cristòfol, C.; Florio, D.; Fioravanti, M.; Zarza, C. Best Therapeutic Practices for the Use of Antibacterial Agents in Finfish Aquaculture: A Particular View on European Seabass (Dicentrarchus Labrax) and Gilthead Seabream (Sparus Aurata) in Mediterranean Aquaculture. Rev. Aquac. 2021, 13, 1285–1323. [Google Scholar] [CrossRef]
- Francino, M.P. Antibiotics and the Human Gut Microbiome: Dysbioses and Accumulation of Resistances. Front. Microbiol. 2015, 6, 1543. [Google Scholar] [CrossRef]
- Langdon, A.; Crook, N.; Dantas, G. The Effects of Antibiotics on the Microbiome throughout Development and Alternative Approaches for Therapeutic Modulation. Genome Med. 2016, 8, 39. [Google Scholar] [CrossRef]
- Looft, T.; Allen, H.K. Collateral Effects of Antibiotics on Mammalian Gut Microbiomes. Gut Microbes 2012, 3, 463–467. [Google Scholar] [CrossRef] [PubMed]
- Kayani, M.R.; Yu, K.; Qiu, Y.; Yu, X.; Chen, L.; Huang, L. Longitudinal Analysis of Exposure to a Low Concentration of Oxytetracycline on the Zebrafish Gut Microbiome. Front. Microbiol. 2022, 13, 985065. [Google Scholar] [CrossRef]
- Payne, C.J.; Turnbull, J.F.; MacKenzie, S.; Crumlish, M. Investigating the Effect of an Oxytetracycline Treatment on the Gut Microbiome and Antimicrobial Resistance Gene Dynamics in Nile Tilapia (Oreochromis Niloticus). Antibiotics 2021, 10, 1213. [Google Scholar] [CrossRef]
- Limbu, S.M.; Zhou, L.; Sun, S.-X.; Zhang, M.-L.; Du, Z.-Y. Chronic Exposure to Low Environmental Concentrations and Legal Aquaculture Doses of Antibiotics Cause Systemic Adverse Effects in Nile Tilapia and Provoke Differential Human Health Risk. Environ. Int. 2018, 115, 205–219. [Google Scholar] [CrossRef] [PubMed]
- Limbu, S.M.; Zhang, H.; Luo, Y.; Chen, L.-Q.; Zhang, M.; Du, Z.-Y. High Carbohydrate Diet Partially Protects Nile Tilapia (Oreochromis Niloticus) from Oxytetracycline-Induced Side Effects. Environ. Pollut. 2020, 256, 113508. [Google Scholar] [CrossRef]
- Hooper, L.V.; Gordon, J.I. Commensal Host-Bacterial Relationships in the Gut. Science 2001, 292, 1115–1118. [Google Scholar] [CrossRef]
- Ley, R.E.; Peterson, D.A.; Gordon, J.I. Ecological and Evolutionary Forces Shaping Microbial Diversity in the Human Intestine. Cell 2006, 124, 837–848. [Google Scholar] [CrossRef]
- Pérez, T.; Balcázar, J.L.; Ruiz-Zarzuela, I.; Halaihel, N.; Vendrell, D.; de Blas, I.; Múzquiz, J.L. Host–Microbiota Interactions within the Fish Intestinal Ecosystem. Mucosal Immunol. 2010, 3, 355–360. [Google Scholar] [CrossRef]
- Lei, Y.M.K.; Nair, L.; Alegre, M.-L. The Interplay between the Intestinal Microbiota and the Immune System. Clin. Res. Hepatol. Gastroenterol. 2015, 39, 9–19. [Google Scholar] [CrossRef]
- Neish, A.S. Microbes in Gastrointestinal Health and Disease. Gastroenterology 2009, 136, 65–80. [Google Scholar] [CrossRef]
- Nagalingam, N.A.; Lynch, S.V. Role of the Microbiota in Inflammatory Bowel Diseases. Inflamm. Bowel Dis. 2012, 18, 968–984. [Google Scholar] [CrossRef]
- Suez, J.; Korem, T.; Zeevi, D.; Zilberman-Schapira, G.; Thaiss, C.A.; Maza, O.; Israeli, D.; Zmora, N.; Gilad, S.; Weinberger, A.; et al. Artificial Sweeteners Induce Glucose Intolerance by Altering the Gut Microbiota. Nature 2014, 514, 181–186. [Google Scholar] [CrossRef]
- Chassaing, B.; Koren, O.; Goodrich, J.K.; Poole, A.C.; Srinivasan, S.; Ley, R.E.; Gewirtz, A.T. Dietary Emulsifiers Impact the Mouse Gut Microbiota Promoting Colitis and Metabolic Syndrome. Nature 2015, 519, 92–96. [Google Scholar] [CrossRef]
- Zhou, K.; Jiang, M.; Qin, X.; Wang, X. Role of Bilirubin in Digestive Proteases Inactivation in the Lower Intestine. Dig. Liver Dis. 2015, 47, 438–439. [Google Scholar] [CrossRef] [PubMed]
- Qin, X. Changes in Complex Microecosystem of Gut and Pathogenesis of Diseases in Modern Society-Impaired Inactivation of Digestive Proteases May Be the Key Event. World J. Complex Med. 2015, 1, 38–43. [Google Scholar]
- Luna, G.M.; Quero, G.M.; Kokou, F.; Kormas, K. Time to Integrate Biotechnological Approaches into Fish Gut Microbiome Research. Curr. Opin. Biotechnol. 2022, 73, 121–127. [Google Scholar] [CrossRef]
- Xiong, J.-B.; Nie, L.; Chen, J. Current Understanding on the Roles of Gut Microbiota in Fish Disease and Immunity. Zool. Res. 2019, 40, 70–76. [Google Scholar] [CrossRef]
- Shi, F.; Huang, Y.; Yang, M.; Lu, Z.; Li, Y.; Zhan, F.; Lin, L.; Qin, Z. Antibiotic-Induced Alternations in Gut Microflora Are Associated with the Suppression of Immune-Related Pathways in Grass Carp (Ctenopharyngodon Idellus). Front. Immunol. 2022, 13, 970125. [Google Scholar] [CrossRef]
- Lu, T.-H.; Chen, C.-Y.; Wang, W.-M.; Liao, C.-M. A Risk-Based Approach for Managing Aquaculture Used Oxytetracycline-Induced TetR in Surface Water Across Taiwan Regions. Front. Pharmacol. 2021, 12, 803499. [Google Scholar] [CrossRef]
- Wasimuddin; Schlaeppi, K.; Ronchi, F.; Leib, S.L.; Erb, M.; Ramette, A. Evaluation of Primer Pairs for Microbiome Profiling from Soils to Humans within the One Health Framework. Mol. Ecol. Resour. 2020, 20, 1558–1571. [Google Scholar] [CrossRef]
- Edgar, R.C. UNOISE2: Improved Error-Correction for Illumina 16S and ITS Amplicon Sequencing. bioRxiv 2016, 081257. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and Clustering Orders of Magnitude Faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly Accurate OTU Sequences from Microbial Amplicon Reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Lagkouvardos, I.; Joseph, D.; Kapfhammer, M.; Giritli, S.; Horn, M.; Haller, D.; Clavel, T. IMNGS: A Comprehensive Open Resource of Processed 16S rRNA Microbial Profiles for Ecology and Diversity Studies. Sci. Rep. 2016, 6, 33721. [Google Scholar] [CrossRef]
- Reitmeier, S.; Hitch, T.C.A.; Treichel, N.; Fikas, N.; Hausmann, B.; Ramer-Tait, A.E.; Neuhaus, K.; Berry, D.; Haller, D.; Lagkouvardos, I.; et al. Handling of Spurious Sequences Affects the Outcome of High-Throughput 16S rRNA Gene Amplicon Profiling. ISME Commun. 2021, 1, 31. [Google Scholar] [CrossRef]
- Pruesse, E.; Peplies, J.; Glöckner, F.O. SINA: Accurate High-Throughput Multiple Sequence Alignment of Ribosomal RNA Genes. Bioinformatics 2012, 28, 1823–1829. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Yoon, S.-H.; Ha, S.-M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A Taxonomically United Database of 16S rRNA Gene Sequences and Whole-Genome Assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Lagkouvardos, I.; Fischer, S.; Kumar, N.; Clavel, T. Rhea: A Transparent and Modular R Pipeline for Microbial Profiling Based on 16S rRNA Gene Amplicons. PeerJ 2017, 5, e2836. [Google Scholar] [CrossRef]
- Intze, E.; Lagkouvardos, I. DivCom: A Tool for Systematic Partition of Groups of Microbial Profiles into Intrinsic Subclusters and Distance-Based Subgroup Comparisons. Front. Bioinform. 2022, 2, 864382. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Bittinger, K.; Charlson, E.S.; Hoffmann, C.; Lewis, J.; Wu, G.D.; Collman, R.G.; Bushman, F.D.; Li, H. Associating Microbiome Composition with Environmental Covariates Using Generalized UniFrac Distances. Bioinformatics 2012, 28, 2106–2113. [Google Scholar] [CrossRef] [PubMed]
- BacDive|The Bacterial Diversity Metadatabase. Available online: https://bacdive.dsmz.de/ (accessed on 29 August 2023).
- Copatti, C.E.; Baldisserotto, B.; de Souza, C.F.; Monserrat, J.M.; Garcia, L. Water pH and Hardness Alter ATPases and Oxidative Stress in the Gills and Kidney of Pacu (Piaractus Mesopotamicus). Neotrop. Ichthyol. 2019, 17, e190032. [Google Scholar] [CrossRef]
- Goodrich, H.R.; Wilson, R.W.; Smullen, R.; Barnes, A.C.; Franklin, C.E. Acidified Fish Feeds Reduce the Energetic and Physiological Costs of Digestion in Juvenile Barramundi (Lates Calcarifer). Aquaculture 2022, 546, 737400. [Google Scholar] [CrossRef]
- Wang, H.; Gu, Y.; Zhou, H.; Chen, J.; Wang, M.; Jiang, H.; Cao, H. Plesiomonas Shigelloides, a Potential Pathogen of Enteritis in Ictalurus Punctatus. Isr. J. Aquac. Bamidgeh 2020, 72, 1–11. [Google Scholar] [CrossRef]
- Zhang, Y.; Wen, B.; David, M.A.; Gao, J.-Z.; Chen, Z.-Z. Comparative Analysis of Intestinal Microbiota of Discus Fish (Symphysodon Haraldi) with Different Growth Rates. Aquaculture 2021, 540, 736740. [Google Scholar] [CrossRef]
- Zhang, H.; Ding, Q.; Wang, A.; Liu, Y.; Teame, T.; Ran, C.; Yang, Y.; He, S.; Zhou, W.; Olsen, R.E.; et al. Effects of Dietary Sodium Acetate on Food Intake, Weight Gain, Intestinal Digestive Enzyme Activities, Energy Metabolism and Gut Microbiota in Cultured Fish: Zebrafish as a Model. Aquaculture 2020, 523, 735188. [Google Scholar] [CrossRef]
- Padra, J.T.; Sundh, H.; Sundell, K.; Venkatakrishnan, V.; Jin, C.; Samuelsson, T.; Karlsson, N.G.; Lindén, S.K. Aeromonas Salmonicida Growth in Response to Atlantic Salmon Mucins Differs between Epithelial Sites, Is Governed by Sialylated and N-Acetylhexosamine-Containing O-Glycans, and Is Affected by Ca2+. Infect. Immun. 2017, 85, e00189-17. [Google Scholar] [CrossRef]
- Legrand, T.P.R.A.; Catalano, S.R.; Wos-Oxley, M.L.; Wynne, J.W.; Weyrich, L.S.; Oxley, A.P.A. Antibiotic-Induced Alterations and Repopulation Dynamics of Yellowtail Kingfish Microbiota. Anim. Microbiome 2020, 2, 26. [Google Scholar] [CrossRef]
- Rosado, D.; Xavier, R.; Severino, R.; Tavares, F.; Cable, J.; Pérez-Losada, M. Effects of Disease, Antibiotic Treatment and Recovery Trajectory on the Microbiome of Farmed Seabass (Dicentrarchus Labrax). Sci. Rep. 2019, 9, 18946. [Google Scholar] [CrossRef]
- Rigos, G.; Nengas, I.; Alexis, M.; Troisi, G.M. Potential Drug (Oxytetracycline and Oxolinic Acid) Pollution from Mediterranean Sparid Fish Farms. Aquat. Toxicol. 2004, 69, 281–288. [Google Scholar] [CrossRef]
- Reverter, M.; Sarter, S.; Caruso, D.; Avarre, J.-C.; Combe, M.; Pepey, E.; Pouyaud, L.; Vega-Heredía, S.; de Verdal, H.; Gozlan, R.E. Aquaculture at the Crossroads of Global Warming and Antimicrobial Resistance. Nat. Commun. 2020, 11, 1870. [Google Scholar] [CrossRef]
- Cabello, F.C.; Godfrey, H.P.; Tomova, A.; Ivanova, L.; Dölz, H.; Millanao, A.; Buschmann, A.H. Antimicrobial Use in Aquaculture Re-Examined: Its Relevance to Antimicrobial Resistance and to Animal and Human Health. Environ. Microbiol. 2013, 15, 1917–1942. [Google Scholar] [CrossRef] [PubMed]
- Leal, J.F.; Esteves, V.I.; Santos, E.B.H. Use of Sunlight to Degrade Oxytetracycline in Marine Aquaculture’s Waters. Environ. Pollut. 2016, 213, 932–939. [Google Scholar] [CrossRef] [PubMed]
- Leung, H.; Yitbarek, A.; Snyder, R.; Patterson, R.; Barta, J.R.; Karrow, N.; Kiarie, E. Responses of Broiler Chickens to Eimeria Challenge When Fed a Nucleotide-Rich Yeast Extract. Poult. Sci. 2019, 98, 1622–1633. [Google Scholar] [CrossRef] [PubMed]
- Montero, D.; Rimoldi, S.; Torrecillas, S.; Rapp, J.; Moroni, F.; Herrera, A.; Gómez, M.; Fernández-Montero, Á.; Terova, G. Impact of Polypropylene Microplastics and Chemical Pollutants on European Sea Bass (Dicentrarchus Labrax) Gut Microbiota and Health. Sci. Total Environ. 2022, 805, 150402. [Google Scholar] [CrossRef]
- Rangel, F.; Enes, P.; Gasco, L.; Gai, F.; Hausmann, B.; Berry, D.; Oliva-Teles, A.; Serra, C.R.; Pereira, F.C. Differential Modulation of the European Sea Bass Gut Microbiota by Distinct Insect Meals. Front. Microbiol. 2022, 13, 831034. [Google Scholar] [CrossRef]
- Chouayekh, H.; Farhat-Khemakhem, A.; Karray, F.; Boubaker, I.; Mhiri, N.; Abdallah, M.B.; Alghamdi, O.A.; Guerbej, H. Effects of Dietary Supplementation with Bacillus Amyloliquefaciens US573 on Intestinal Morphology and Gut Microbiota of European Sea Bass. Probiotics Antimicro. Prot. 2023, 15, 30–43. [Google Scholar] [CrossRef] [PubMed]
- Busti, S.; Rossi, B.; Volpe, E.; Ciulli, S.; Piva, A.; D’Amico, F.; Soverini, M.; Candela, M.; Gatta, P.P.; Bonaldo, A.; et al. Effects of Dietary Organic Acids and Nature Identical Compounds on Growth, Immune Parameters and Gut Microbiota of European Sea Bass. Sci. Rep. 2020, 10, 21321. [Google Scholar] [CrossRef]
- Liu, Y.; Cheng, J.; Xia, Y.; Li, X.; Liu, Y.; Liu, P. Response Mechanism of Gut Microbiome and Metabolism of European Seabass (Dicentrarchus Labrax) to Temperature Stress. Sci. Total Environ. 2022, 813, 151786. [Google Scholar] [CrossRef]
- Kokou, F.; Sasson, G.; Mizrahi, I.; Cnaani, A. Antibiotic Effect and Microbiome Persistence Vary along the European Seabass Gut. Sci. Rep. 2020, 10, 10003. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| OTU ID | Closest Known Species | Growth Optimal pH | Prevalence | Mean Abundance |
|---|---|---|---|---|
| Zotu20 | Alkalihalobacillus pseudofirmus (100%) | 9.7 | 10 | 20.9 |
| Zotu64 | Anaerobacillus isosaccharinicus (97.76%) | 9.7 | 8 | 5.7 |
| Zotu57 | Halomonas desiderata (98.88) | 9.5 | 8 | 11.7 |
| Zotu26 | Phaeobacter porticola (99.52%) | 7.6 | 4 | 10.7 |
| Zotu60 | Neptuniibacter pectenicola (99.55%) | 7.6 | 3 | 5.6 |
| Zotu70 | Nereida ignava (100%) | 7.6 | 3 | 4.8 |
| Zotu143 | Staphylococcus epidermidis (99.78%) | 7.3 | 4 | 14.6 |
| Zotu86 | Pseudomonas azotoformans (100%) | 7.3 | 4 | 10.3 |
| Zotu152 | Massilia jejuensis (99.32%) | 7.2 | 3 | 6.2 |
| Zotu21 | Pseudomonas aeruginosa (100%) | 7 | 4 | 30.9 |
| Zotu87 | Kocuria palustris (100%) | 7 | 7 | 6.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rigas, D.; Grivas, N.; Nelli, A.; Gouva, E.; Skoufos, I.; Kormas, K.; Tzora, A.; Lagkouvardos, I. Persistent Dysbiosis, Parasite Rise and Growth Impairment in Aquacultured European Seabass after Oxytetracycline Treatment. Microorganisms 2023, 11, 2302. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms11092302
Rigas D, Grivas N, Nelli A, Gouva E, Skoufos I, Kormas K, Tzora A, Lagkouvardos I. Persistent Dysbiosis, Parasite Rise and Growth Impairment in Aquacultured European Seabass after Oxytetracycline Treatment. Microorganisms. 2023; 11(9):2302. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms11092302
Chicago/Turabian StyleRigas, Dimitris, Nikos Grivas, Aikaterini Nelli, Evangelia Gouva, Ioannis Skoufos, Konstantinos Kormas, Athina Tzora, and Ilias Lagkouvardos. 2023. "Persistent Dysbiosis, Parasite Rise and Growth Impairment in Aquacultured European Seabass after Oxytetracycline Treatment" Microorganisms 11, no. 9: 2302. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms11092302