
Isolation and Taxonomic Identity of Bacteriocin-Producing Lactic Acid Bacteria from Retail Foods and Animal Sources


Abstract
:1. Introduction
2. Experimental Section
2.1. Bacterial Cultures, Growth Conditions, and Storage
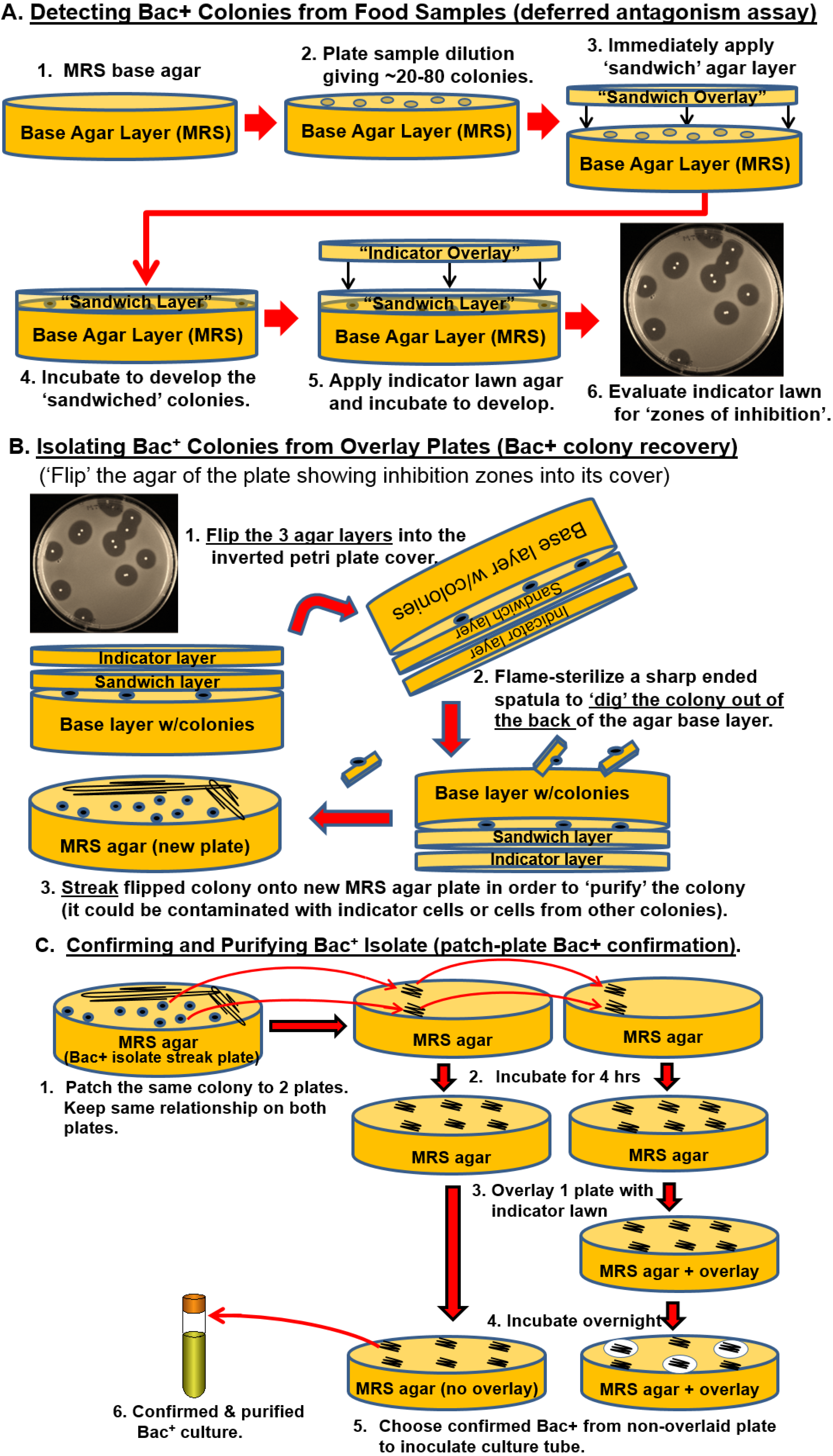
2.2. Isolation of Bacteriocin-Producing (Bac+) LAB from Food and Animal Sources
2.3. Exclusion of Other Inhibitors

2.4. Extraction of Total Bacterial DNA
2.5. Identification of Isolates by 16S rRNA Gene Amplification, Sequencing, and Analysis
3. Results
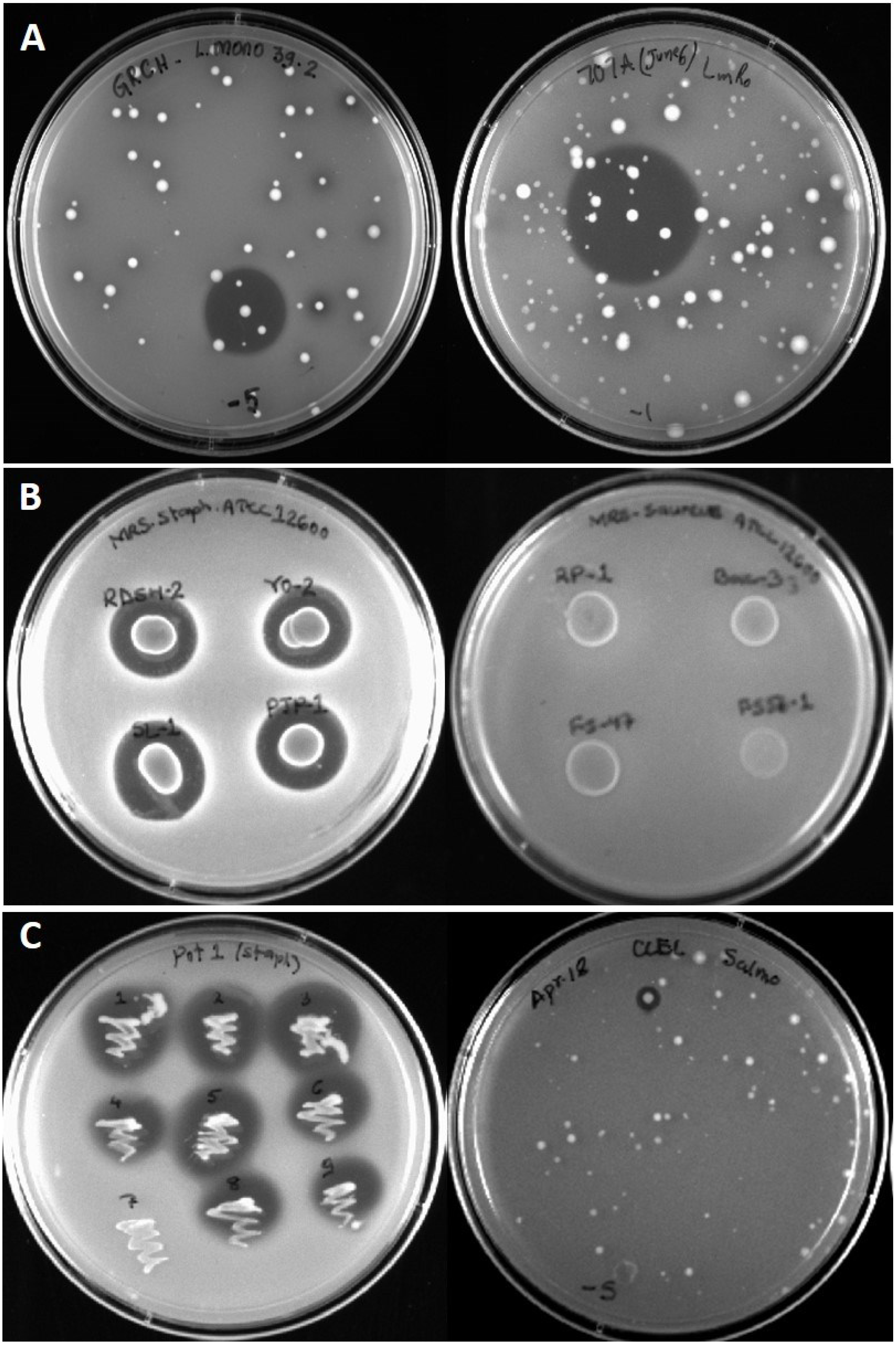
3.1. Isolation of Bac+ Strains of LAB

{kind=link}
{kind=link}
{kind=link}
| Organism | Strain | Source/Reference |
|---|---|---|
| Enterococcus faecium | 326F | This study; bovine fecal sample |
| Enterococcus faecium | FS97-2 | Vegetable [9] |
| Enterococcus faecium | JCP B-5 | This study; pork sausage |
| Enterococcus faecium | JCP M-2 | This study; pork sausage |
| Enterococcus faecium | Thyme 2 | This study; thyme |
| Enterococcus faecium | Thyme 3 | This study; thyme |
| Enterococcus faecium | Milk 5 | This study; raw milk |
| Enterococcus faecium | JCP-9 | This study; pork sausage |
| Enterococcus faecium | Milk 12 | This study; raw milk |
| Enterococcus faecium | Pop 4 | This study; dog feces |
| Enterococcus faecium | FS56-1 | Mushrooms [9] |
| Enterococcus durans | FS707 | This study; bovine fecal sample |
| Enterococcus hirae | 323F | This study; bovine fecal sample |
| Enterococcus hirae | 323 RL1 | This study; bovine rumen fluid |
| Enterococcus hirae | 341 FA | This study; bovine fecal sample |
| Enterococcus thailandicus | RP-1 | Raw pork [5] |
| Enterococcus thailandicus | FS92 | Raw pork [9] |
| Enterococcus faecalis | BJ-12 | Muriana culture collection |
| Enterococcus faecalis | BJ-13 | Muriana culture collection |
| Enterococcus faecalis | BJ-27 | Muriana culture collection |
| Carnobacterium maltaromaticum | LGBF-1 | This study; ground beef |
| Carnobacterium maltaromaticum | GBF-1 | This study; ground beef |
| Carnobacterium maltaromaticum | COG-1 | This study; collard greens |
| Carnobacterium maltaromaticum | COG-2 | This study; collard greens |
| Carnobacterium maltaromaticum | CHW-1 | This study; chicken wings |
| Carnobacterium maltaromaticum | TOF-1 | This study; tofu |
| Carnobacterium maltaromaticum | GPK-1 | This study; ground pork |
| Carnobacterium maltaromaticum | GAC-1 | This study; ground chuck |
| Carnobacterium maltaromaticum | GAC-2 | This study; ground chuck |
| Pediococcus acidilactici | Bac3 | Ground turkey [5] |
| Pediococcus acidilactici | FS707 S4 | This study; bovine fecal sample |
| Lactobacillus ingluviei | FS60 | Cheese [9] |
| Lactobacillus curvatus | BJ-5 | Muriana culture collection |
| Lactobacillus curvatus | BJ-18 | Muriana culture collection |
| Lactobacillus curvatus | BJ-21 | Muriana culture collection |
| Lactobacillus curvatus | BEEF 2L-1 | This study; ground beef |
| Lactobacillus curvatus | BEEF 3 | This study; ground beef |
| Lactobacillus curvatus | FS36-1 | Ground beef [9] |
| Lactobacillus curvatus | FS44-B | Ground pork [9] |
| Lactobacillus curvatus | FS47 | Ground beef [9] |
| Lactobacillus curvatus | FS80-2 | Muriana culture collection |
| Lactobacillus sakei | FS707 S1 | This study; bovine fecal sample |
| Leuconostoc mesenteroides | BFS-1 | This study; breakfast sausage |
| Staphylococcus gallolyticus | 707RS | This study; bovine rumen fluid |
| Staphylococcus gallolyticus | 341RL | This study; bovine rumen fluid |
| Lactococcus lactis | BJ-23 | Muriana culture collection |
| Lactococcus lactis | FL-1 | This study; lettuce (foxy) |
| Lactococcus lactis | FL S-1 | This study; lettuce (foxy) |
| Lactococcus lactis | FS91-1 | Fruit [9] |
| Lactococcus lactis | FS95 | Vegetables [9] |
| Lactococcus lactis | FS162 | Muriana culture collection |
| Lactococcus lactis | ASPG-1 | This study; asparagus |
| Lactococcus lactis | PJP-1 | This study; peppers |
| Lactococcus lactis | RDSH-1 | This study; radish (red) |
| Lactococcus lactis | SL-1 | This study; shredded lettuce |
| Lactococcus lactis | YO-1 | This study; yellow onion |
| Lactococcus lactis | SP-1 | This study; sweet potato |
| Lactococcus lactis | RD | This study; radish |
| Lactococcus lactis | GBN-1 | This study; green beans |
| Lactococcus lactis | BSP | This study; bean sprouts |
| Serratia plymuthica | POT-1 | This study; Russet Potato |
| Serratia ficaria | CCEL-1 | This study; Chinese celery |
3.2. Differentiation of Bacterial Isolates
3.3. Inhibition of Foodborne Pathogens

| Select Bacteriocin-Producing Strains | Listeria monocytogenes 39-2 | Staphylococcus aureus ATCC 12600 | Staphylococcus aureus ISP 178 | Enterococcus faecalis ATCC 19433 |
|---|---|---|---|---|
| Enterococcus thailandicus RP-1 | ++ 1 | - | - | ND 2 |
| Enterococcus faecium FS56-1 | ++++ | - | - | ND |
| Enterococcus durans FS707 | ++++ | - | - | ++ |
| Lactobacillus curvatus FS47 | ++++ | - | - | - |
| Lactobacillus curvatus Beef3 | ++++ | - | - | - |
| Lactobacillus curvatus FS44-B | +++ | - | - | - |
| Lactococcus lactis BSP | +++ | ++ | ++ | + |
| Lactococcus lactis SL1 | ++ | +++ | +++ | + |
| Lactococcus lactis RDSH-1 | +++ | +++ | +++ | + |
| Lactococcus lactis ASPG-1 | +++ | +++ | +++ | + |
| Lactococcus lactis YO-1 | +++ | +++ | +++ | + |
| Lactococcus lactis FLS1 | + | + | + | + |
| Lactococcus lactis GBN-1 | ++ | + | + | + |
| Lactococcus lactis PJP-1 | +++ | +++ | +++ | + |
| Pediococcus acidilactici Bac3 | ++++ | - | - | ND |
| Serratia plymuthica POT-1 3 | ++ | ++ | ++ | + |
| Serratia ficaria CCEL-1 3 | + | + | + | + |
3.4. Phylogenetic Analysis
4. Discussion
5. Conclusions

Acknowledgments
Author Contributions
Conflicts of Interest
References
- Nes, I.F.; Diep, D.B.; Holo, H. Bacteriocin diversity in Streptococcus and Enterococcus. J. Bacteriol. 2006, 189, 1189–1198. [Google Scholar] [CrossRef] [PubMed]
- Nes, I.F.; Johnsborg, O. Exploration of antimicrobial potential in lab by genomics. Curr. Opin. Biotechnol. 2004, 15, 100–104. [Google Scholar] [CrossRef] [PubMed]
- Casaus, P.; Nilsen, T.; Cintas, L.M.; Nes, I.F.; Hernandez, P.E.; Holo, H. Enterocin B, a new bacteriocin from enterococcus faecium T136 which can act synergistically with enterocin A. Microbiology 1997, 143 Pt 7, 2287–2294. [Google Scholar] [CrossRef] [PubMed]
- Knoll, C.; Divol, B.; du Toit, M. Genetic screening of lactic acid bacteria of oenological origin for bacteriocin-encoding genes. Food Microbiol. 2008, 25, 983–991. [Google Scholar] [CrossRef] [PubMed]
- Macwana, S.J.; Muriana, P.M. A “bacteriocin pcr array” for identification of bacteriocin-related structural genes in lactic acid bacteria. J. Microbiol. Methods 2012, 88, 197–204. [Google Scholar] [CrossRef] [PubMed]
- De Jong, A.; van Hijum, S.A.; Bijlsma, J.J.; Kok, J.; Kuipers, O.P. Bagel: A web-based bacteriocin genome mining tool. Nucleic Acids Res. 2006, 34, W273–W279. [Google Scholar] [CrossRef] [PubMed]
- Cleveland, J.; Montville, T.J.; Nes, I.F.; Chikindas, M.L. Bacteriocins: Safe, natural antimicrobials for food preservation. Int. J. Food Microbiol. 2001, 71, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Macwana, S.; Muriana, P.M. Spontaneous bacteriocin resistance in listeria monocytogenes as a susceptibility screen for identifying different mechanisms of resistance and modes of action by bacteriocins of lactic acid bacteria. J. Microbiol. Methods 2012, 88, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Garver, K.I.; Muriana, P.M. Detection, identification and characterization of bacteriocin-producing lactic-acid bacteria from retail food-products. Int. J. Food Microbiol. 1993, 19, 241–258. [Google Scholar] [CrossRef] [PubMed]
- Muriana, P.M.; Klaenhammer, T.R. Purification and partial characterization of lactacin-F, a bacteriocin produced by lactobacillus-acidophilus-11088. Appl. Environ. Microbiol. 1991, 57, 114–121. [Google Scholar] [PubMed]
- Garver, K.I.; Muriana, P.M. Purification and partial amino-acid-sequence of curvaticin FS47, a heat-stable bacteriocin produced by lactobacillus-curvatus FS47. Appl. Environ. Microbiol. 1994, 60, 2191–2195. [Google Scholar] [PubMed]
- Muriana, P.M. Bacteriocins for control of Listeria spp. in food. J. Food Protect. 1996, Supplement, 54–63. [Google Scholar]
- Muriana, P.M.; Klaenhammer, T.R. Conjugal transfer of plasmid-encoded determinants for bacteriocin production and immunity in lactobacillus-acidophilus-88. Appl. Environ. Microbiol. 1987, 53, 553–560. [Google Scholar] [PubMed]
- Coton, E.; Coton, M. Multiplex pcr for colony direct detection of gram-positive histamine- and tyramine-producing bacteria. J. Microbiol. Methods 2005, 63, 296–304. [Google Scholar] [CrossRef] [PubMed]
- Turner, S.; Pryer, K.M.; Miao, V.P.W.; Palmer, J.D. Investigating deep phylogenetic relationships among cyanobacteria and plastids by small subunit RRNA sequence analysis1. J. Eukaryot. Microbiol. 1999, 46, 327–338. [Google Scholar] [CrossRef] [PubMed]
- Henning, C.; Gautam, D.; Muriana, P. Identification of Multiple Bacteriocins in Enterococcus spp. Using an Enterococcus-Specific Bacteriocin PCR Array. Microorganisms 2015, 3, 1–16. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Bosshard, P.P.; Zbinden, R.; Abels, S.; Böddinghaus, B.; Altwegg, M.; Böttger, E.C. 16s RRNA gene sequencing vs. the API 20 NE system and the VITEK 2 ID-GNB card for identification of nonfermenting Gram-negative bacteria in the clinical laboratory. J. Clin. Microbiol. 2006, 44, 1359–1366. [Google Scholar] [CrossRef] [PubMed]
- Kulwichit, W.; Nilgate, S.; Chatsuwan, T.; Krajiw, S.; Unhasuta, C.; Chongthaleong, A. Accuracies of leuconostoc phenotypic identification: A comparison of API systems and conventional phenotypic assays. BMC Infect. Dis. 2007, 7, 69. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Henning, C.; Vijayakumar, P.; Adhikari, R.; Jagannathan, B.; Gautam, D.; Muriana, P.M. Isolation and Taxonomic Identity of Bacteriocin-Producing Lactic Acid Bacteria from Retail Foods and Animal Sources. Microorganisms 2015, 3, 80-93. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms3010080
Henning C, Vijayakumar P, Adhikari R, Jagannathan B, Gautam D, Muriana PM. Isolation and Taxonomic Identity of Bacteriocin-Producing Lactic Acid Bacteria from Retail Foods and Animal Sources. Microorganisms. 2015; 3(1):80-93. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms3010080
Chicago/Turabian StyleHenning, Chris, Paul Vijayakumar, Raj Adhikari, Badrinath Jagannathan, Dhiraj Gautam, and Peter M. Muriana. 2015. "Isolation and Taxonomic Identity of Bacteriocin-Producing Lactic Acid Bacteria from Retail Foods and Animal Sources" Microorganisms 3, no. 1: 80-93. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms3010080