
Correia Repeat Enclosed Elements and Non-Coding RNAs in the Neisseria Species


Abstract
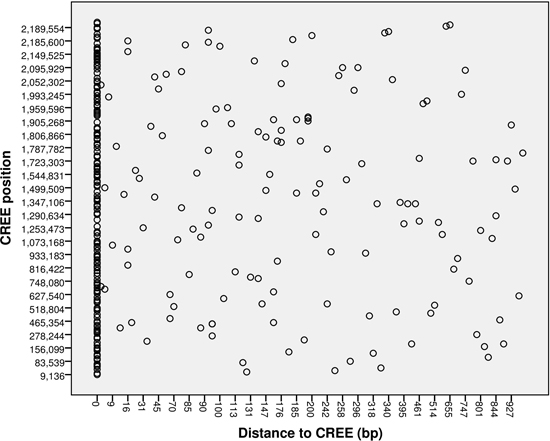
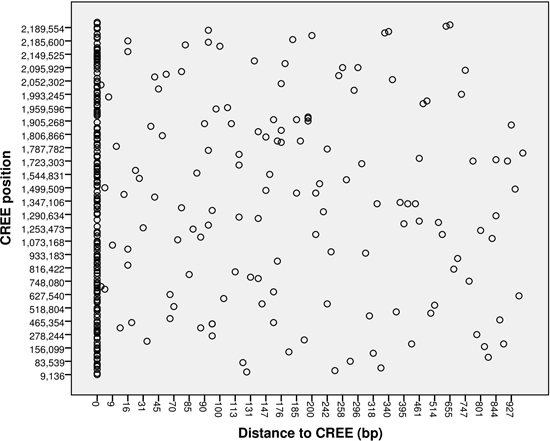
:
1. Introduction
2. Materials and Methods
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Chisholm, S.A.; Wilson, J.; Alexander, S.; Tripodo, F.; Al-Shahib, A.; Schaefer, U.; Lythgow, K.; Fifer, H. An outbreak of high-level azithromycin resistant Neisseria gonorrhoeae in England. Sex. Transm. Infect. 2016, 92, 365–367. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Furuya, R.; Irie, S.; Kanayama, A.; Kobayashi, I. High prevalence of azithromycin-resistant Neisseria gonorrhoeae isolates with a multidrug resistance phenotype in Fukuoka, Japan. Sex. Transm. Dis. 2015, 42, 337–341. [Google Scholar] [CrossRef] [PubMed]
- Demczuk, W.; Martin, I.; Peterson, S.; Bharat, A.; van Domselaar, G.; Graham, M.; Lefebvre, B.; Allen, V.; Hoang, L.; Tyrrell, G.; et al. Genomic epidemiology and molecular resistance mechanisms of azithromycin-resistant Neisseria gonorrhoeae in Canada from 1997 to 2014. J. Clin. Microbiol. 2016, 54, 1304–1313. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.Y.; Cao, W.L.; Li, X.D.; Bi, C.; Yang, R.D.; Liang, Y.H.; Li, P.; Ye, X.D.; Chen, X.X.; Zhang, X.B. Azithromycin-resistant Neisseria gonorrhoeae isolates in Guangzhou, China (2009–2013): Coevolution with decreased susceptibilities to ceftriaxone and genetic characteristics. BMC Infect. Dis. 2016, 16, 152. [Google Scholar] [CrossRef] [PubMed]
- Hoang, L.M.N.; Thomas, E.; Tyler, S.; Pollard, A.J.; Stephens, G.; Gustafson, L.; McNabb, A.; Pocock, I.; Tsang, R.; Tan, R. Rapid and fatal meningococcal disease due to a strain of Neisseria meningitidis containing the capsule null locus. Clin. Infect. Dis. 2005, 40, e38–e42. [Google Scholar] [CrossRef] [PubMed]
- Stephens, D.S.; Greenwood, B.; Brandtzaeg, P. Epidemic meningitis, meningococcaemia, and Neisseria meningitidis. Lancet 2007, 369, 2196–2210. [Google Scholar] [CrossRef]
- Caugant, D.A. Genetics and evolution of Neisseria meningitidis: Importance for the epidemiology of meningococcal disease. Infect. Genet. Evol. 2008, 8, 558–565. [Google Scholar] [CrossRef] [PubMed]
- Azzari, C.; Nieddu, F.; Moriondo, M.; Indolfi, G.; Canessa, C.; Ricci, S.; Bianchi, L.; Serranti, D.; Poggi, G.M.; Resti, M. Underestimation of Invasive Meningococcal Disease in Italy. Emerg. Infect. Dis. 2016, 22, 469–475. [Google Scholar] [CrossRef] [PubMed]
- Dinan, A.M.; Loftus, B.J. Non-translational medicine: Targeting bacterial RNA. Front. Genet. 2013, 4, 230. [Google Scholar] [CrossRef] [PubMed]
- Edson, J.A.; Kwon, Y.J. RNAi for silencing drug resistance in microbes toward development of nanoantibiotics. J. Control. Release 2014, 189, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Turner, M.; Galloway, A.; Vigorito, E. Noncoding RNA and its associated proteins as regulatory elements of the immune system. Nat. Immunol. 2014, 15, 484–491. [Google Scholar] [CrossRef] [PubMed]
- Wagner, E.G.; Romby, P. Small RNAs in bacteria and archaea: Who they are, what they do, and how they do it. Adv. Genet. 2015, 90, 133–208. [Google Scholar] [PubMed]
- Mellin, J.R.; Goswami, S.; Grogan, S.; Tjaden, B.; Genco, C.A. A novel fur- and iron-regulated small RNA, NrrF, is required for indirect fur-mediated regulation of the sdhA and sdhC genes in Neisseria meningitidis. J. Bacteriol. 2007, 189, 3686–3694. [Google Scholar] [CrossRef] [PubMed]
- Cahoon, L.A.; Seifert, H.S. Transcription of a cis-acting, noncoding, small RNA is required for pilin antigenic variation in Neisseria gonorrhoeae. PLoS Pathog. 2013, 9, e1003074. [Google Scholar] [CrossRef] [PubMed]
- Fagnocchi, L.; Bottini, S.; Golfieri, G.; Fantappiè, L.; Ferlicca, F.; Antunes, A.; Guadagnuolo, S.; Del Tordello, E.; Siena, E.; Serruto, D.; et al. Global transcriptome analysis reveals small RNAs affecting Neisseria meningitidis bacteremia. PLoS ONE 2015, 10, e0126325. [Google Scholar] [CrossRef] [PubMed]
- Jackson, L.A.; Pan, J.C.; Day, M.W.; Dyer, D.W. Control of RNA stability by NrrF, an iron-regulated small RNA in Neisseria gonorrhoeae. J. Bacteriol. 2013, 195, 5166–5173. [Google Scholar] [CrossRef] [PubMed]
- McClure, R.; Balasubramanian, D.; Sun, Y.; Bobrovskyy, M.; Sumby, P.; Genco, C.A.; Vanderpool, C.K.; Tjaden, B. Computational analysis of bacterial RNA-Seq data. Nucleic Acids Res. 2013, 41, e140. [Google Scholar] [CrossRef] [PubMed]
- Ramsey, M.E.; Bender, T.; Klimowicz, A.K.; Hackett, K.T.; Yamamoto, A.; Jolicoeur, A.; Callaghan, M.M.; Wassarman, K.M.; van der Does, C.; Dillard, J.P. Targeted mutagenesis of intergenic regions in the Neisseria gonorrhoeae gonococcal genetic island reveals multiple regulatory mechanisms controlling type IV secretion. Mol. Microbiol. 2015, 97, 1168–1185. [Google Scholar] [CrossRef] [PubMed]
- Tan, F.Y.; Wörmann, M.E.; Loh, E.; Tang, C.M.; Exley, R.M. Characterization of a novel antisense RNA in the major pilin locus of Neisseria meningitidis influencing antigenic variation. J. Bacteriol. 2015, 197, 1757–1768. [Google Scholar] [CrossRef] [PubMed]
- Wachter, J.; Hill, S.A. Small transcriptome analysis indicates that the enzyme RppH influences both the quality and quantity of sRNAs in Neisseria gonorrhoeae. FEMS Microbiol. Lett. 2015, 362, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Gunesekere, I.C.; Kahler, C.M.; Ryan, C.S.; Snyder, L.A.; Saunders, N.J.; Rood, J.I.; Davies, J.K. Ecf, an alternative sigma factor from Neisseria gonorrhoeae, controls expression of msrAB, which encodes methionine sulfoxide reductase. J. Bacteriol. 2006, 188, 3463–3469. [Google Scholar] [CrossRef] [PubMed]
- Overton, T.W.; Whitehead, R.; Li, Y.; Snyder, L.A.; Saunders, N.J.; Smith, H.; Cole, J.A. Coordinated regulation of the Neisseria gonorrhoeae-truncated denitrification pathway by the nitric oxide-sensitive repressor, NsrR, and nitrite-insensitive NarQ-NarP. J. Biol. Chem. 2006, 281, 33115–33126. [Google Scholar] [CrossRef] [PubMed]
- Whitehead, R.N.; Overton, T.W.; Snyder, L.A.; McGowan, S.J.; Smith, H.; Cole, J.A.; Saunders, N.J. The small FNR regulon of Neisseria gonorrhoeae: Comparison with the larger Escherichia coli FNR regulon and interaction with the NarQ-NarP regulon. BMC Genom. 2007, 8, 35. [Google Scholar] [CrossRef] [PubMed]
- Fantappiè, L.; Oriente, F.; Muzzi, A.; Serruto, D.; Scarlato, V.; Delany, I. A novel Hfq-dependent sRNA that is under FNR control and is synthesized in oxygen limitation in Neisseria meningitidis. Mol. Microbiol. 2011, 80, 507–523. [Google Scholar] [CrossRef] [PubMed]
- Isabella, V.M.; Clark, V.L. Deep sequencing-based analysis of the anaerobic stimulon in Neisseria gonorrhoeae. BMC Genom. 2011, 12, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Saunders, N.J.; Davies, J.K. The use of the pan-Neisseria microarray and experimental design for transcriptomics studies of Neisseria. Methods Mol. Biol. 2012, 799, 295–317. [Google Scholar] [PubMed]
- Haas, B.J.; Chin, M.; Nusbaum, C.; Birren, B.W.; Livny, J. How deep is deep enough for RNA-Seq profiling of bacterial transcriptomes? BMC Genom. 2012, 13, 734. [Google Scholar] [CrossRef] [PubMed]
- Remmele, C.W.; Xian, Y.; Albrecht, M.; Faulstich, M.; Fraunholz, M.; Heinrichs, E.; Dittrich, M.T.; Müller, T.; Reinhardt, R.; Rudel, T. Transcriptional landscape and essential genes of Neisseria gonorrhoeae. Nucleic Acids Res. 2014, 42, 10579–10595. [Google Scholar] [CrossRef] [PubMed]
- Snyder, L.A.; Cole, J.A.; Pallen, M.J. Comparative analysis of two Neisseria gonorrhoeae genome sequences reveals evidence of mobilization of Correia Repeat Enclosed Elements and their role in regulation. BMC Genom. 2009, 10, 70. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.V.; Saunders, N.J.; Jeffries, A.; Rest, R.F. Genome analysis and strain comparison of correia repeats and correia repeat-enclosed elements in pathogenic Neisseria. J. Bacteriol. 2002, 184, 6163–6173. [Google Scholar] [CrossRef] [PubMed]
- Bentley, S.D.; Vernikos, G.S.; Snyder, L.A.; Churcher, C.; Arrowsmith, C.; Chillingworth, T.; Cronin, A.; Davis, P.H.; Holroyd, N.E.; Jagels, K.; et al. Meningococcal genetic variation mechanisms viewed through comparative analysis of serogroup C strain FAM18. PLoS Genet. 2007, 3, e23. [Google Scholar] [CrossRef] [PubMed]
- Correia, F.F.; Inouye, S.; Inouye, M. A 26-base-pair repetitive sequence specific for Neisseria gonorrhoeae and Neisseria meningitidis genomic DNA. J. Bacteriol. 1986, 167, 1009–1015. [Google Scholar] [PubMed]
- Correia, F.F.; Inouye, S.; Inouye, M. A family of small repeated elements with some transposon-like properties in the genome of Neisseria gonorrhoeae. J. Biol. Chem. 1988, 263, 12194–12198. [Google Scholar] [PubMed]
- Livny, J. Bioinformatic discovery of bacterial regulatory RNAs using SIPHT. Methods Mol. Biol. 2012, 905, 3–14. [Google Scholar] [PubMed]
- Rice, P.; Longden, L.; Bleasby, A. EMBOSS: The European molecular biology open software suite. Trends Genet. 2000, 16, 276–277. [Google Scholar] [CrossRef]
- Chaudhuri, R.R.; Loman, N.J.; Snyder, L.A.S.; Bailey, C.M.; Stekel, D.J.; Pallen, M.J. xBASE2: A comprehensive resource for comparative bacterial genomics. Nucleic Acids Res. 2008, 36, D543–D546. [Google Scholar] [CrossRef] [PubMed]
- De Gregorio, E.; Abrescia, C.; Carlomagno, M.S.; di Nocera, P.P. Asymmetrical distribution of Neisseria miniature insertion sequence DNA repeats among pathogenic and nonpathogenic Neisseria strains. Infect. Immun. 2003, 71, 4217–4221. [Google Scholar] [CrossRef] [PubMed]
- Marri, P.R.; Paniscus, M.; Weyand, N.J.; Rendón, M.A.; Calton, C.M.; Hernández, D.R.; Higashi, D.L.; Sodergren, E.; Weinstock, G.M.; Rounsley, S.D.; et al. Genome sequencing reveals widespread virulence gene exchange among human Neisseria species. PLoS ONE 2010, 5, e11835. [Google Scholar] [CrossRef] [PubMed]
- Huis in ′t Veld, R.A.; Willemsen, A.M.; van Kampen, A.H.; Bradley, E.J.; Baas, F.; Pannekoek, Y.; van der Ende, A. Deep sequencing whole transcriptome exploration of the σE regulon in Neisseria meningitidis. PLoS ONE 2011, 6, e29002. [Google Scholar] [CrossRef] [PubMed]
- Rouquette-Loughlin, C.E.; Balthazar, J.T.; Hill, S.A.; Shafer, W.M. Modulation of the mtrCDE-encoded efflux pump gene complex of Neisseria meningitidis due to a Correia element insertion sequence. Mol. Microbiol. 2004, 54, 731–741. [Google Scholar] [CrossRef] [PubMed]
- Mazzone, M.; de Gregorio, E.; Lavitola, A.; Pagliarulo, C.; Alifano, P.; di Nocera, P.P. Whole-genome organization and functional properties of miniature DNA insertion sequences conserved in pathogenic Neisseriae. Gene 2001, 278, 211–222. [Google Scholar] [CrossRef]
- De Gregorio, E.; Abrescia, C.; Carlomagno, M.S.; di Nocera, P.P. The abundant class of nemis repeats provides RNA substrates for ribonuclease III in Neisseriae. Biochim. Biophys. Acta 2002, 1576, 39–44. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Strain | Intergenic Regions | SIPHT ncRNAs 1 |
|---|---|---|
| N. gonorrhoeae strain NCCP11945 | 2255 | 760 |
| N. gonorrhoeae strain FA1090 | 1806 | 976 |
| N. meningitidis strain MC58 | 2015 | 912 |
| N. meningitidis strain Z2491 | 1846 | 996 |
| N. meningitidis strain FAM18 | 1833 | 959 |
| N. meningitidis strain alpha14 | 1782 | 976 |
| N. meningitidis strain 53442 | 1881 | 846 |
| N. lactamica strain 020-06 | 1873 | 890 |
| Strain | Intergenic Regions | CREE 1 |
|---|---|---|
| N. gonorrhoeae strain NCCP11945 | 2255 | 131 |
| N. gonorrhoeae strain FA1090 | 1806 | 123 |
| N. meningitidis strain MC58 | 2015 | 248 |
| N. meningitidis strain Z2491 | 1846 | 260 |
| N. meningitidis strain FAM18 | 1833 | 249 |
| N. meningitidis strain alpha14 | 1782 | 255 |
| N. meningitidis strain 53442 | 1881 | 234 |
| N. lactamica strain 020-06 | 1873 | 92 |
| Strain | Intergenic Regions | SIPHT ncRNAs 1 | CREE 2 | within 1 kb 3 | 1 kb % 4 | within 300 bp 5 | 300 bp % 6 | Overlap 7 | Overlap % 8 |
|---|---|---|---|---|---|---|---|---|---|
| N. gonorrhoeae strain NCCP11945 | 2255 | 760 | 131 | 96 | 73% | 86 | 66% | 75 | 57% |
| N. gonorrhoeae strain FA1090 | 1806 | 976 | 123 | 100 | 81% | 92 | 75% | 73 | 59% |
| N. meningitidis strain MC58 | 2015 | 912 | 248 | 164 | 66% | 131 | 53% | 87 | 35% |
| N. meningitidis strain Z2491 | 1846 | 996 | 260 | 188 | 72% | 151 | 58% | 99 | 38% |
| N. meningitidis strain FAM18 | 1833 | 959 | 249 | 183 | 74% | 143 | 57% | 98 | 39% |
| N. meningitidis strain alpha14 | 1782 | 976 | 255 | 191 | 75% | 147 | 58% | 95 | 37% |
| N. meningitidis strain 53442 | 1881 | 846 | 234 | 151 | 65% | 127 | 54% | 85 | 36% |
| N. lactamica strain 020-06 | 1873 | 890 | 92 | 72 | 78% | 59 | 64% | 36 | 39% |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roberts, S.B.; Spencer-Smith, R.; Shah, M.; Nebel, J.-C.; Cook, R.T.; Snyder, L.A.S. Correia Repeat Enclosed Elements and Non-Coding RNAs in the Neisseria Species. Microorganisms 2016, 4, 31. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms4030031
Roberts SB, Spencer-Smith R, Shah M, Nebel J-C, Cook RT, Snyder LAS. Correia Repeat Enclosed Elements and Non-Coding RNAs in the Neisseria Species. Microorganisms. 2016; 4(3):31. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms4030031
Chicago/Turabian StyleRoberts, Sabrina B., Russell Spencer-Smith, Mahwish Shah, Jean-Christophe Nebel, Richard T. Cook, and Lori A. S. Snyder. 2016. "Correia Repeat Enclosed Elements and Non-Coding RNAs in the Neisseria Species" Microorganisms 4, no. 3: 31. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms4030031