Geographic Distribution of Avirulence Genes of the Rice Blast Fungus Magnaporthe oryzae in the Philippines


Abstract
:1. Introduction
2. Materials and Mathods
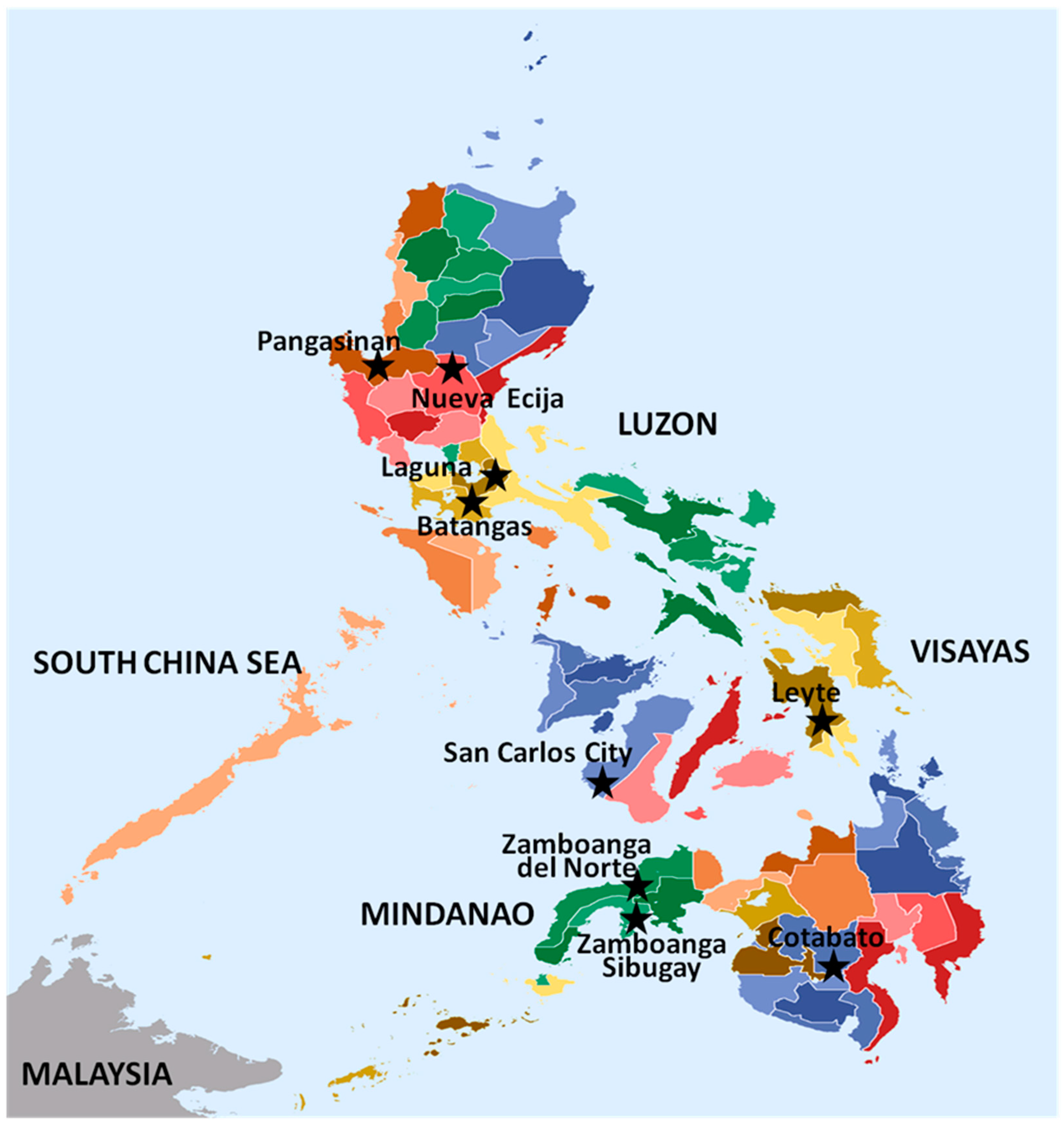
2.1. Magnaporthe oryzae Isolates and DNA Extraction
2.2. PCR Assay for the Presence of Avirulence Genes
2.3. Data Analysis
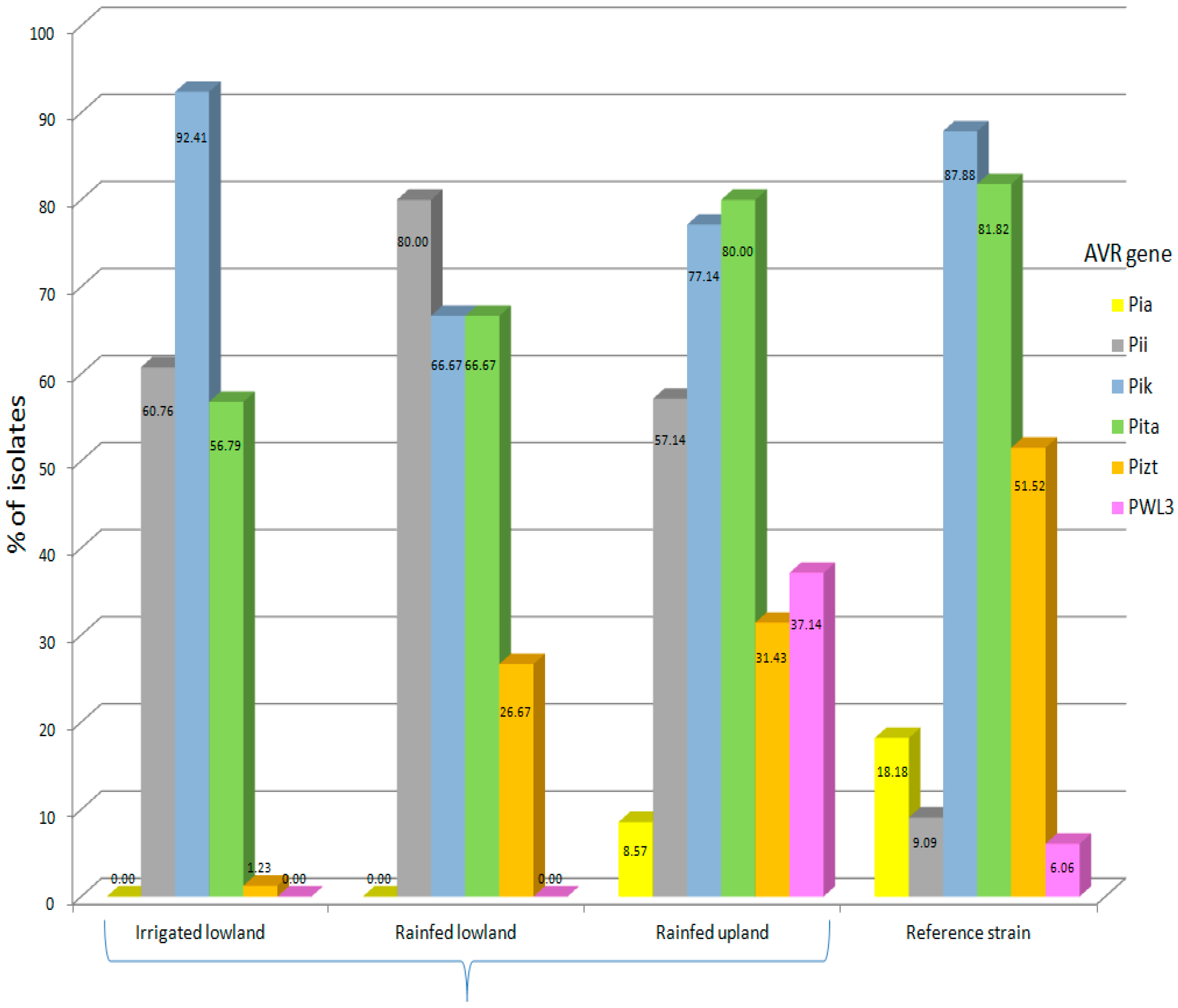
3. Results and Discussion
3.1. Avirulence Genes in the Isolates
3.2. Resistance Genes Present in Rice Genotypes
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Valent, B. Rice blast as a model system for plant pathology. Phytopathology 1990, 80, 33–36. [Google Scholar] [CrossRef]
- Liu, E.-M.; Xiao, Y.-I.; Yi, Y.J.; Zhuang, J.Y.; Zheng, K.I.; Lou, F. Relationship between blast resistance phenotypes and resistance gene analogue profiles in rice. Rice Sci. 2005, 2, 75–82. [Google Scholar]
- Silué, D.; Notteghem, J.L.; Tharreau, D. Evidence of a gene-for-gene relationship in the Oryza sativa Magnaporthe grisea pathosystem. Phytopathology 1992, 82, 577–580. [Google Scholar] [CrossRef]
- Rouxel, T.; Balesdent, M.-H. Avirulence genes. In Encyclopedia of Life Sciences (ELS); John Wiley & Sons, Ltd.: Chichester, UK, 2010. [Google Scholar] [CrossRef]
- Flor, H.H. Current status of the gene-for-gene concept. Annu. Rev. Phytopathol. 1971, 9, 275–296. [Google Scholar] [CrossRef]
- Lauge, R.; De Wit, P.J.G.M. Fungal avirulence genes: Structure and possible functions. Fungal Genet. Biol. 1998, 24, 285–297. [Google Scholar] [CrossRef] [PubMed]
- Hua, L.; Chen, C.; Wu, W.; He, X.; Lin, F.; Wang, X.; Wang, L. The isolation of Pi1, an allele at the Pik locus which confers broad spectrum resistance to rice blast. Appl. Gen. 2012, 125, 1047–1055. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; McAdams, S.A.; Bryan, G.T.; Hershey, H.P.; Valent, B. Direct interaction of resistance gene and avirulence gene. Products confers rice blast resistance. EMBO J. 2000, 19, 4004–4014. [Google Scholar] [CrossRef]
- Kang, S.; Sweigard, J.A.; Valent, B. The PWL host specificity gene family in the blast fungus Magnaporthe grisea. Mol. Plant Microbe Interact. 1995, 8, 939–948. [Google Scholar] [CrossRef]
- Sweigard, J.A.; Carroll, A.M.; Kang, S.; Farrall, L.; Chumley, F.G.; Valent, B. Identification, cloning, and characterization of PWL2, a gene for host species specificity in the rice blast fungus. Plant Cell 1995, 7, 1221–1233. [Google Scholar] [CrossRef]
- Orbach, M.J.; Farrall, L.; Sweigard, J.A.; Chumley, F.G.; Valent, B. A telomeric avirulence gene determines efficacy for the rice blast resistance gene Pi-ta. Plant Cell 2000, 12, 2019–2032. [Google Scholar] [CrossRef]
- Bohnert, H.; Fudal, I.; Dioh, W.; Tharreau, D.; Notteghem, J.L.; Lebrun, M.H. A putative polyketide synthase/peptide synthetase from Magnaporthe grisea signals pathogen attack to resistant rice. Plant Cell 2004, 16, 2499–2513. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Wang, B.; Wu, J.; Lu, G.; Hu, Y.; Zhang, X.; Zhang, Z.; Zhao, Q.; Feng, Q.; Zhang, H.; et al. The Magnaporthe oryzae avirulence gene AvrPiz-t encodes a predicted secreted protein that triggers the immunity in rice mediated by the blast resistance gene Piz-t. Mol. Plant Microbe Interact. 2009, 22, 411–420. [Google Scholar] [CrossRef] [PubMed]
- Miki, S.; Matsui, K.; Kito, H.; Otsuka, K.; Ashizawa, T. Molecular cloning and characterization of the AVR-Pia locus from a Japanese field isolate of Magnaporthe Oryzae. Mol. Plant Pathol. 2009, 10, 361–374. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.; Saitoh, H.; Fujisawa, S.; Kanzaki, H.; Matsumura, H.; Yoshida, K.; Tosa, Y.; Chuma, I.; Takano, Y.; Win, J.; et al. Association genetics reveals three novel avirulence genes from the rice blast fungal pathogen Magnaporthe oryzae. Plant Cell 2009, 21, 1573–1591. [Google Scholar] [CrossRef] [PubMed]
- Ribot, C.; Césari, S.; Abidi, I.; Chalvon, V.; Bournaud, C.; Vallet, J.; Lebrun, M.-H.; Morel, J.-B.; Kroj, T. The Magnaporthe oryzae effector AVR1-CO39 is translocated into rice cells independently of a fungal-derived machinery. Plant J. 2013, 74, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Kou, Y.; Bao, J.; Li, Y.; Tang, M.; Zhu, X.; Ponaya, A.; Xiao, G.; Li, J.; Li, C.; et al. Comparative genomics identifies the Magnaporthe oryzae avirulence effector AvrPi9 that triggers Pi9-mediated blast resistance in rice. New Phytol. 2015, 206, 1463–1467. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, N.; Masako, T.; Noguchi, T.; Fujita, Y. Partial mapping of avirulence genes AVR-Pii and AVR-Pia in the rice blast fungus Magnaporthe oryzae. Can. J. Plant Pathol. 2006, 28, 494–498. [Google Scholar] [CrossRef]
- Sone, T.; Takeuchi, S.; Miki, Y.; Satoh, K.; Ohtsuka, A.; Abe, A.; Asano, K. Homologous recombination causes the spontaneous deletion of AVR-Pia in Magnaporthe oryzae. FEMS Microbiol. Lett. 2012, 339, 102–109. [Google Scholar] [CrossRef]
- Li, P.; Bai, B.; Zhang, H.; Zhou, H.; Zhou, B. Genomic organization and sequence dynamics of the AvrPiz-t locus in Magnaporthe oryzae. J. Zhejiang Univ. Sci. B 2012, 13, 452–464. [Google Scholar] [CrossRef]
- Kanzaki, H.; Yoshida, K.; Saitoh, H.; Fujisaka, K.; Hirabuchi, A.; Alaux, L.; Fournier, E.; Tharreau, D.; Terauchi, R. Arms race co-evolution of Magnaporthe oryzae AVR-Pik and rice Pik genes driven by their physical interactions. Plant J. 2012, 72, 894–907. [Google Scholar] [CrossRef]
- Chuma, I.; Isobe, C.; Hotta, Y.; Ibaragi, K.; Futamata, N.; Kusaba, M.; Yoshida, K.; Terauchi, R.; Fujita, Y.; Nakayashiki, H.; et al. Multiple translocation of the AVR-Pita effector gene among chromosomes of the rice blast fungus Magnaporthe oryzae and related species. PLoS Pathog. 2011, 7, e1002147. [Google Scholar] [CrossRef] [PubMed]
- Van der Does, H.C.; Rep, M. Virulence genes and the evolution of host specificity in plant-pathogenic fungi. Mol. Plant–Microbe Interact. 2007, 20, 1175–1182. [Google Scholar] [CrossRef] [PubMed]
- Inoue, Y.; Vy, T.P.P.; Yoshida, K.; Asano, H.; Mitsuoka, C.; Asuke, S.; Anh, V.L.; Cumagun, C.J.R.; Chuma, I.; Terauchi, R.; et al. Evolution of the wheat blast fungus through functional losses in a host specificity determinant. Science 2017, 357, 80–83. [Google Scholar] [CrossRef] [PubMed]
- Lopez, A.L.C.; Cumagun, C.J.R.; Didier, T. Diversity of ACE1 genotypes of the rice blast fungus (Magnaporthe oryzae B.C. Couch) in the Philippines. IAMURE Int. J. Ecol. Conserv. 2015, 16, 80–92. [Google Scholar] [CrossRef]
- Bonman, J.M.; Khush, G.S.; Nelson, R.J. Breeding rice for resistance to pests. Annu. Rev. Phytopathol. 1992, 30, 507–528. [Google Scholar] [CrossRef]
- Ou, S.H. Pathogen variability and host resistance in rice blast disease. Annu. Rev. Phytopathol. 1980, 18, 167–187. [Google Scholar] [CrossRef]
- Sweigard, J.A.; Orbach, M.J.; Valent, B.; Chumley, F.G. A miniprep procedure for isolating genomic DNA from Magnaporthe grisea. Fungal Genet. Rep. 1990, 37, 28. [Google Scholar] [CrossRef]
- Chen, D.; Zeigler, R.S.; Leung, H.; Nelson, R.J. Population structure of Pyricularia grisea at two screening sites in the Philippines. Phytopathology 1995, 85, 1011–1020. [Google Scholar] [CrossRef]
- Stukenbrock, E.H.; McDonald, B.A. Population genetics of fungal and oomycete effectors involved in gene-for-gene interactions. Mol. Plant Microbe Interact. 2009, 22, 371–380. [Google Scholar] [CrossRef]
- Tharreau, D.; Fudal, I.; Andriantsimialona, D.; Kadrawi, S.; Utami, D.W.; Fournier, E.; Lebrun, M.-H.; Nottéghem, J.L. World Population Structure and Migration of the Rice Blast Fungus, Magnaporthe oryzae. In Advances in Genetics, Genomics and Control of Rice Blast Disease; Wang, G.L., Valent, B., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 209–215. [Google Scholar]
- Jia, Y.; Zhang, Z.; Xing, J.; Wang, J.; Correll, J.C.; Cartwright, R. Natural Variation and Evolution of the Avirulence Genes in Magnaporthe oryzae. 2012. Available online: https://www.ars.usda.gov/research/publications/publication/?seqNo115=279490 (accessed on 16 January 2019).
- Lu, L.; Li, C.; Fan, H.; Jia, Y.; Li, J. Selection and mutation of the avirulence gene AVR-Pii of the rice blast fungus Magnaporthe oryzae. Plant Pathol. 2018, 68, 127–134. [Google Scholar] [CrossRef]
- Imam, J.; Alam, S.; Mandal, N.P.; Shukla, P.; Sharma, T.R.; Variar, M. Molecular identification and virulence analysis of AVR genes in rice blast pathogen, Magnaporthe oryzae from Eastern India. Euphytica 2015, 206, 21–31. [Google Scholar] [CrossRef]
- Sirisathaworn, T.; Srirat, T.; Longya, A.; Jantasuriyarat, C. Evaluation of mating type distribution and genetic diversity of three Magnaporthe oryzae avirulence genes, PWL-2, AVR-Pii and Avr-Piz-t, in Thailand rice blast isolates. Agric. Nat. Resour. 2017, 51, 7–14. [Google Scholar] [CrossRef]
- Lai, X.H.; Marchetti, M.A.; Petersen, H.D. Comparative slow-blasting in rice grown under upland and flooded blast nursery culture. Plant Dis. 1999, 83, 681–684. [Google Scholar] [CrossRef]
- Li, J.; Yang, J.; Liu, L.; He, H.; He, X.; Zhu, Y.; Li, C. Geographic distribution of avirulence genes in rice blast fungus in Yunnan Province, China. J. Res. Ecol. 2011, 2, 181–185. [Google Scholar]
- Lopez, A.L.C.; Cumagun, C.J.R. Genetic structure of Magnaporthe oryzae populations in three island groups in the Philippines. Eur. J. Plant Pathol. 2019, 153, 181–198. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence 5’–3’ | AVR Gene | Expected Product Size (bp) |
|---|---|---|---|
| YL169b | CGACCCGTTTCCGCC | Avr-Pita | 1086 |
| YL149b | TGACCGCGATTCCCTCCATT | ||
| Z21 | AATCCCGTCACTTTCATTCTCCA | Avr-Pizt | 637 |
| Z22 | GTCGCAAGCCTCGTACTACCTTT | ||
| Z23 | TCCAATTTATTCAACTGCCACTC | Avr-Pik | 532 |
| Z24 | GTAAACCTCGTCAAACCTCCCTA | ||
| Z27 | CCCATTATCTTACCAGTCGCTTGA | Avr-Pia | 868 |
| Z28 | ATTCCTCCCGTAAACAGTAAACC | ||
| ZZ41 | TGCAGGCCCAAATCCGTAGGAA | Avr-Pii | 508 |
| ZZ42 | ACTGTCCGCCGCTCGTTTGG | ||
| PW3-F | TGCGTGCTCATTTGTAAACC | Avr-PWL3 | 937 |
| PW3-R | TTGACGGTACTAGGGCTGCT |
| χ2 Value* | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Geographical Location/Type of Agroecosystem | Avr-Pita | Avr-Pia | Avr-Pizt | Avr-Pik | Avr-Pii | Avr-PWL3 | ||||||
| 0 | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 1 | |
| A. Geographic Location | ||||||||||||
| Luzon | ||||||||||||
| E | 28.60 | 53.39 | 80.09 | 1.91 | 71.83 | 10.17 | 12.07 | 69.92 | 31.15 | 50.86 | 73.74 | 8.26 |
| χ2 | 0.74 | 0.40 | 0.02 | 0.63 | 0.37 | 2.63 | 0.07 | 0.01 | 0.04 | 0.03 | 0.25 | 2.20 |
| Mindanao | ||||||||||||
| E | 6.98 | 13.02 | 19.53 | 0.47 | 17.52 | 2.48 | 2.95 | 17.05 | 7.60 | 12.40 | 17.98 | 2.02 |
| χ2 | 7.07 | 3.79 | 0.01 | 0.46 | 0.36 | 2.56 | 0.30 | 0.05 | 0.76 | 0.47 | 0.23 | 2.02 |
| Visayas | ||||||||||||
| E | 9.42 | 17.58 | 26.37 | 0.63 | 23.65 | 3.34 | 3.97 | 23.02 | 10.26 | 16.74 | 24.28 | 2.72 |
| χ2 | 0.62 | 0.33 | 0.02 | 0.63 | 0.30 | 2.10 | 0.00 | 0.00 | 0.15 | 0.09 | 1.62 | 14.92 |
| B. Type of Agroecosystem | ||||||||||||
| Irrigated lowland | ||||||||||||
| E | 27.56 | 51.44 | 77.16 | 1.84 | 69.20 | 9.79 | 11.64 | 67.36 | 39.24 | 60.76 | 71.04 | 7.96 |
| χ2 | 1.075 | 0.58 | 0.04 | 1.84 | 1.12 | 7.90 | 2.73 | 0.47 | 0.03 | 0.02 | 0.89 | 7.96 |
| Rainfed lowland | ||||||||||||
| E | 5.23 | 9.77 | 14.65 | 0.35 | 13.14 | 1.86 | 2.21 | 12.79 | 20.00 | 80.00 | 13.49 | 1.51 |
| χ2 | 0.01 | 0.01 | 0.01 | 0.35 | 0.35 | 2.46 | 3.53 | 0.61 | 1.28 | 0.78 | 0.17 | 1.15 |
| Rainfed upland | ||||||||||||
| E | 12.21 | 22.79 | 34.19 | 0.82 | 30.66 | 4.34 | 5.16 | 29.84 | 42.86 | 57.14 | 31.47 | 3.53 |
| χ2 | 2.22 | 1.19 | 0.14 | 5.87 | 1.45 | 10.21 | 1.57 | 0.27 | 0.22 | 0.13 | 2.85 | 25.44 |
| Host Genotype | Resistance Gene * | |||||
|---|---|---|---|---|---|---|
| Pita | Pia | Pizt | Pik | Pii | PWL3 | |
| A. Irrigated Lowland | ||||||
| 012–N | 0 | 0 | 0 | 1 | 1 | 0 |
| 15 IL | 1 | 0 | 0 | 1 | 1 | 0 |
| Au 108 | 1 | 0 | 0 | 1 | 1 | 0 |
| Au 63 | 1 | 0 | 0 | 1 | 1 | 0 |
| Bigante | 0 | 0 | 0 | 1 | 1 | 0 |
| C9305-B-9-2 | 0 | 0 | 0 | 1 | 0 | 0 |
| Dinorado | 1 | 0 | 0 | 1 | 0 | 0 |
| ILMAS 540 | 1 | 0 | 0 | 1 | 1 | 0 |
| IR 50 | 1 | 0 | 0 | 1 | 1 | 0 |
| IR83140-B-36-B | 0 | 0 | 0 | 1 | 0 | 0 |
| IR86781-3-3-1-1 | 1 | 0 | 1 | 1 | 1 | 0 |
| LMAS 544 | 1 | 0 | 0 | 1 | 1 | 0 |
| MS 11 | 0 | 0 | 0 | 1 | 0 | 0 |
| NSIC Rc130 | 1 | 0 | 0 | 1 | 0 | 0 |
| NSIC Rc212 | 0 | 0 | 0 | 1 | 1 | 0 |
| NSIC Rc216 | 1 | 0 | 0 | 1 | 1 | 0 |
| PNG 719 | 1 | 0 | 0 | 1 | 1 | 0 |
| PR34350-4-POKKALI-24-M5R-10 | 0 | 0 | 0 | 1 | 1 | 0 |
| PR36723-B-13-3-3-3 | 0 | 0 | 0 | 1 | 0 | 0 |
| PR36930-B-7-3 | 0 | 0 | 0 | 1 | 0 | 0 |
| PR37088-B-9-1-1 | 0 | 0 | 0 | 1 | 0 | 0 |
| PR37624-1-5-1-2-1 | 0 | 0 | 0 | 1 | 0 | 0 |
| PR37801-15-1-1-3-2-B-B | 0 | 0 | 0 | 0 | 1 | 0 |
| PYT 132 | 1 | 0 | 0 | 1 | 0 | 0 |
| PYT 172 | 1 | 0 | 0 | 1 | 1 | 0 |
| PYT 20 | 1 | 0 | 0 | 1 | 1 | 0 |
| PYT 210 | 1 | 0 | 0 | 1 | 1 | 0 |
| PYT 218 | 1 | 0 | 0 | 1 | 0 | 0 |
| PYT 238 | 1 | 0 | 0 | 1 | 0 | 0 |
| PYT 307 | 1 | 0 | 0 | 1 | 1 | 0 |
| PYT 324 | 1 | 0 | 0 | 0 | 1 | 0 |
| PYT 41 | 1 | 0 | 0 | 1 | 1 | 0 |
| B. Rainfed Lowland | ||||||
| Bigante | 1 | 0 | 1 | 1 | 1 | 0 |
| C4 | 0 | 0 | 0 | 0 | 1 | 0 |
| NSIC Rc222 | 1 | 0 | 0 | 1 | 1 | 0 |
| C. Rainfed upland | ||||||
| Japanese | 1 | 0 | 1 | 1 | 0 | 1 |
| Milagrosa | 1 | 1 | 1 | 1 | 1 | 1 |
| NSIC Rc152 | 1 | 0 | 1 | 1 | 1 | 1 |
| NSIC Rc216 | 1 | 0 | 0 | 1 | 1 | 0 |
| PhilRice M3 | 1 | 0 | 1 | 1 | 1 | 1 |
| Waray | 1 | 1 | 1 | 1 | 1 | 1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lopez, A.L.C.; Yli-Matilla, T.; Cumagun, C.J.R. Geographic Distribution of Avirulence Genes of the Rice Blast Fungus Magnaporthe oryzae in the Philippines. Microorganisms 2019, 7, 23. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7010023
Lopez ALC, Yli-Matilla T, Cumagun CJR. Geographic Distribution of Avirulence Genes of the Rice Blast Fungus Magnaporthe oryzae in the Philippines. Microorganisms. 2019; 7(1):23. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7010023
Chicago/Turabian StyleLopez, Ana Liza C., Tapani Yli-Matilla, and Christian Joseph R. Cumagun. 2019. "Geographic Distribution of Avirulence Genes of the Rice Blast Fungus Magnaporthe oryzae in the Philippines" Microorganisms 7, no. 1: 23. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7010023