Bioremediation of Artificial Diesel-Contaminated Soil Using Bacterial Consortium Immobilized to Plasma-Pretreated Wood Waste



Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Diesel Emulsion in Mineral Medium
2.2. Acquisition of a Diesel-Degrading Microbial Consortium from Oil-Polluted Soil
2.3. Measuring Diesel Concentration with Gas Chromatography−Flame Ionization Detector (GC−FID) Analysis
2.4. Exposure to Cold Low-Pressure Nitrogen Plasma
2.5. Biofilm Formation on the Wood Waste
2.6. Measurements of the Biofilm Viability
2.7. Measuring the Turbidity of the Bacteria in the Biofilm Attached to the Wood Waste
2.8. Preparation of Artificially Diesel-Contaminated Soil for Biodegradation Experiments
2.9. Microbial Community Based on 16S rRNA Analysis
2.10. Statistics
3. Results and Discussion
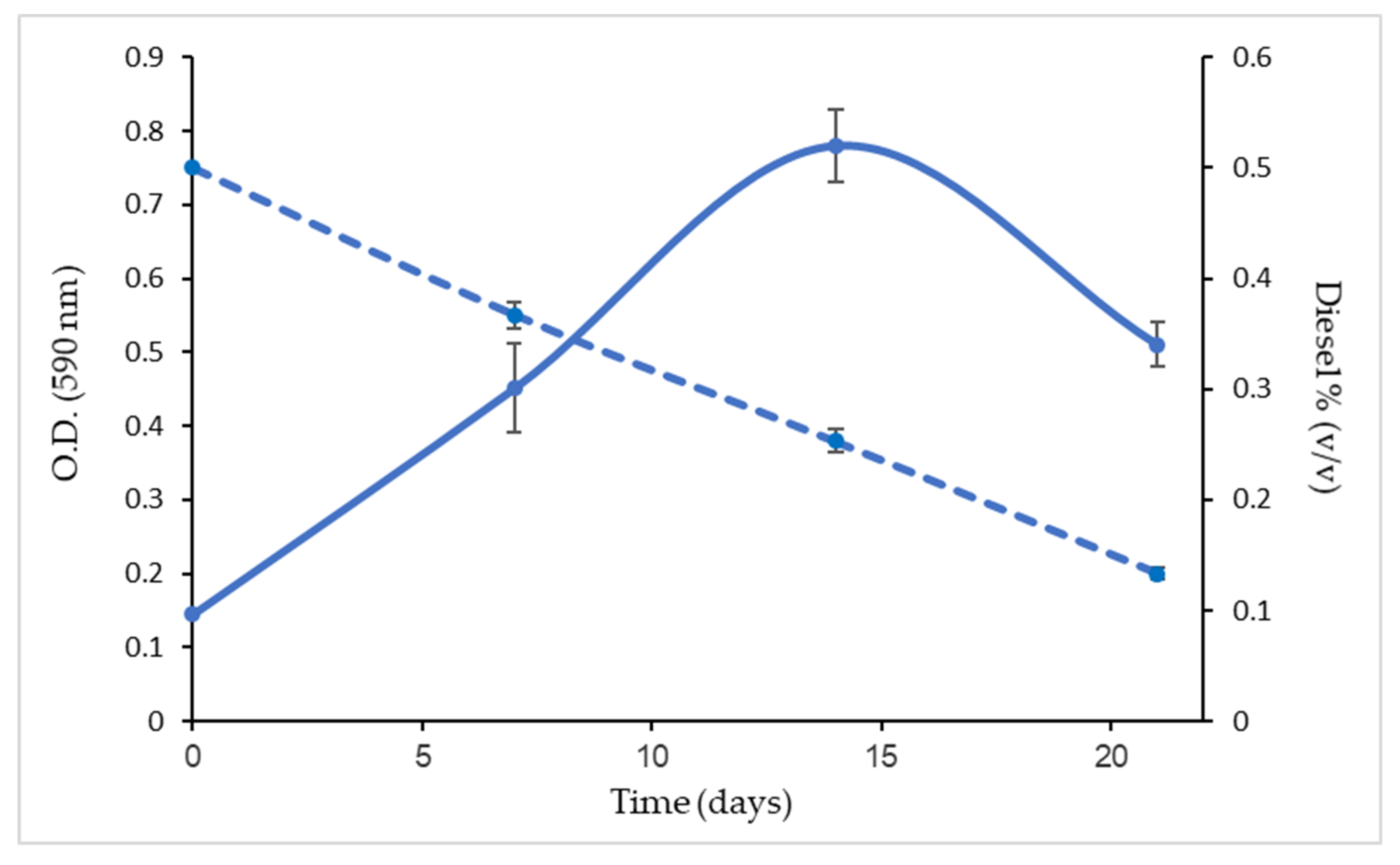
3.1. Bacterial Consortium Growth Curve in MM with Diesel as the Carbon Source
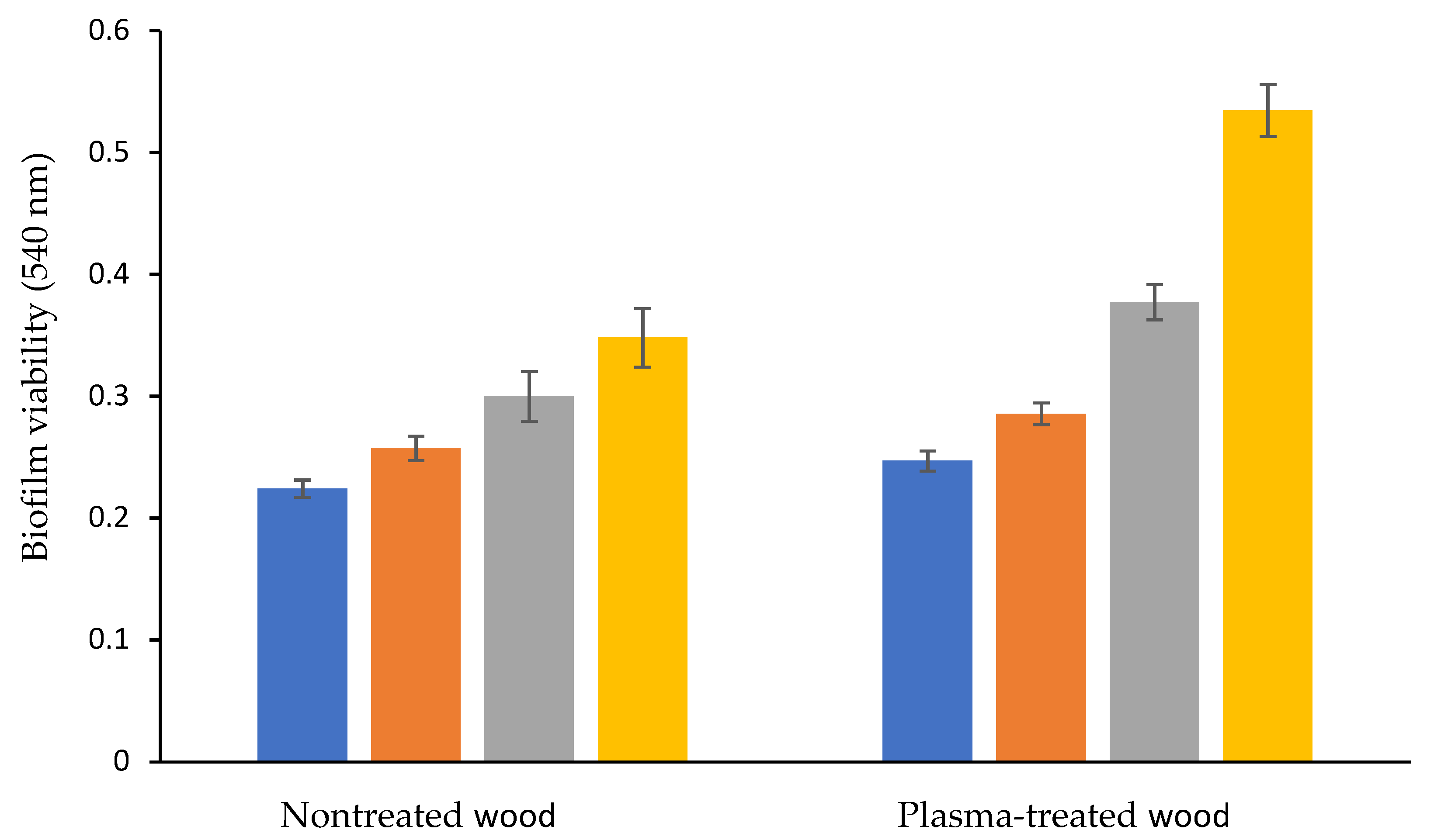
3.2. Biofilm Formation on Plasma-Treated and Nontreated Wood Waste
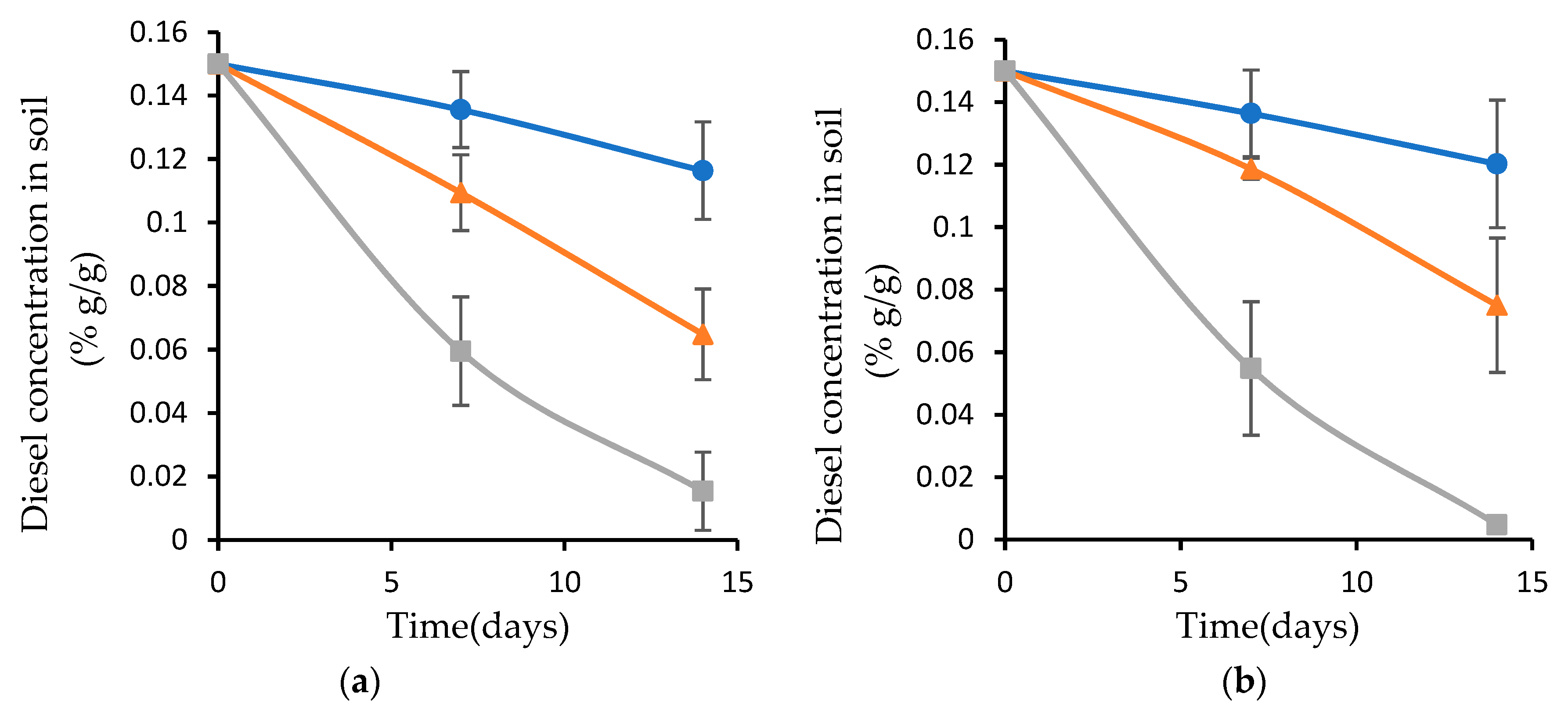
3.3. Diesel Concentration in Artificially Contaminated Soil
3.4. Diesel Concentration in Artificially Contaminated Soil Inoculated with Biofilm Attached to Plasma-Treated and Nontreated Wood Waste
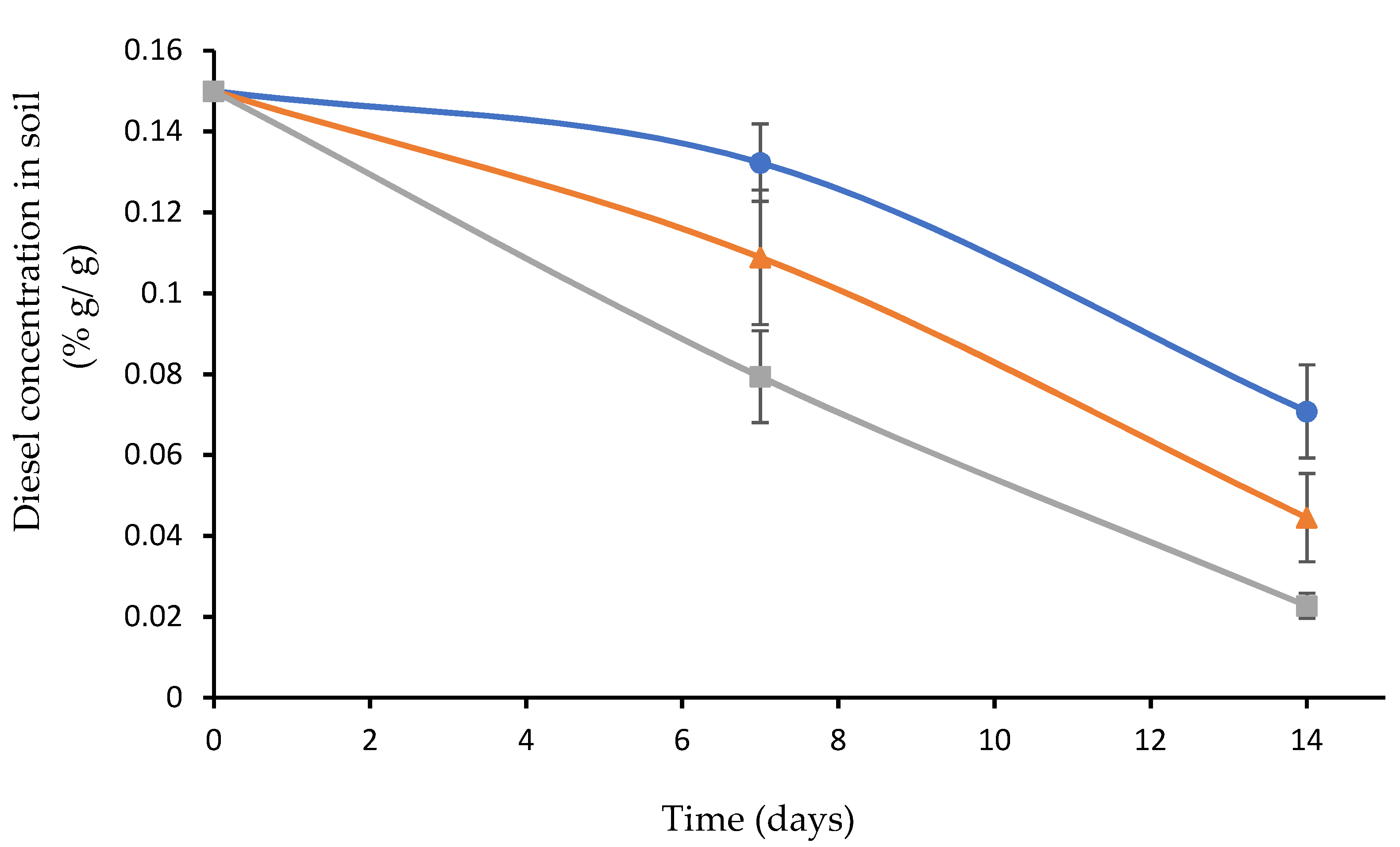
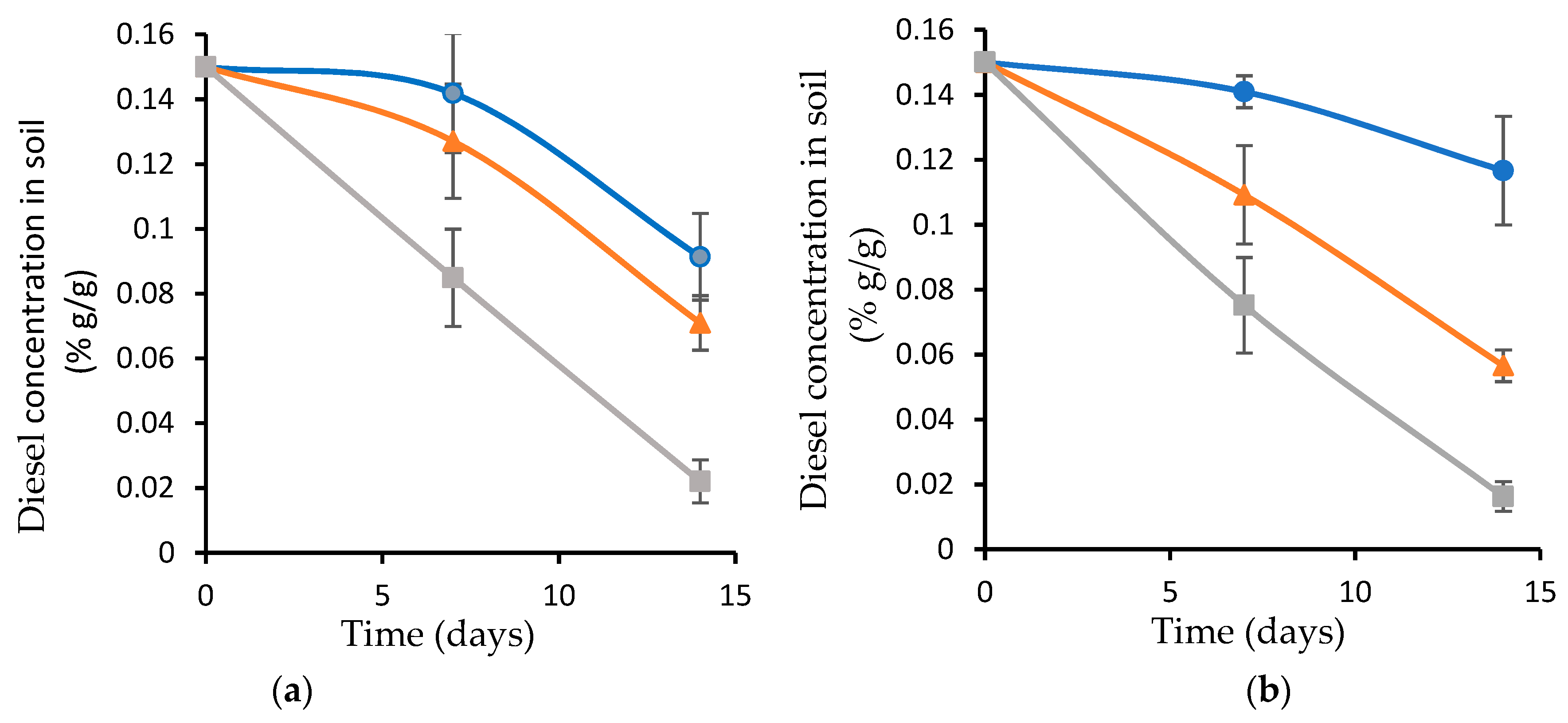
3.5. Diesel Biodegradation as a Function of a Pre-Exposure to High Temperature
3.6. Diesel Biodegradation as a Function of Soil Acidity
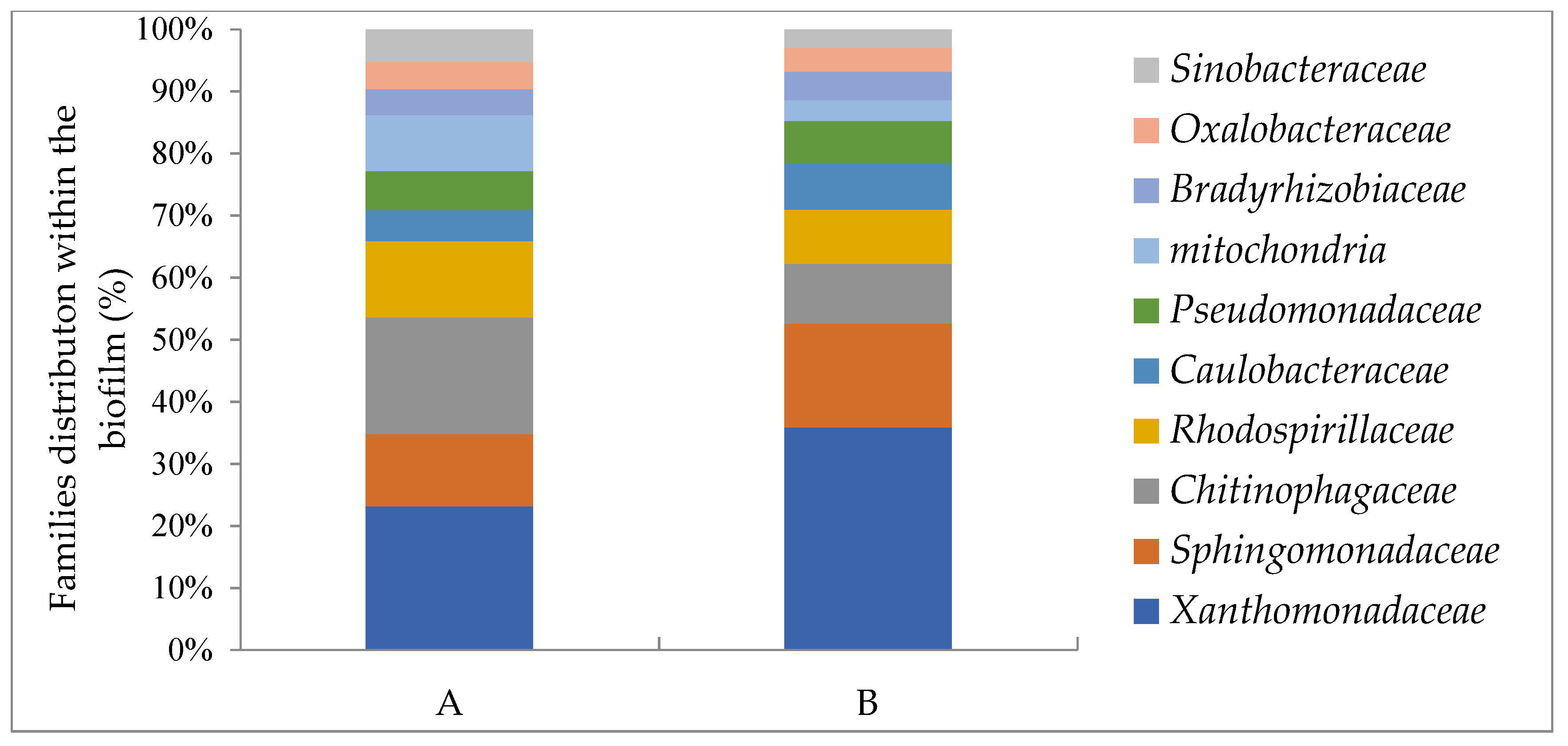
3.7. Microbial Distribution Analysis (Based on 16S rRNA) in the Original Oil-Contaminated Site and in the Biofilms

{kind=link}
{kind=link}
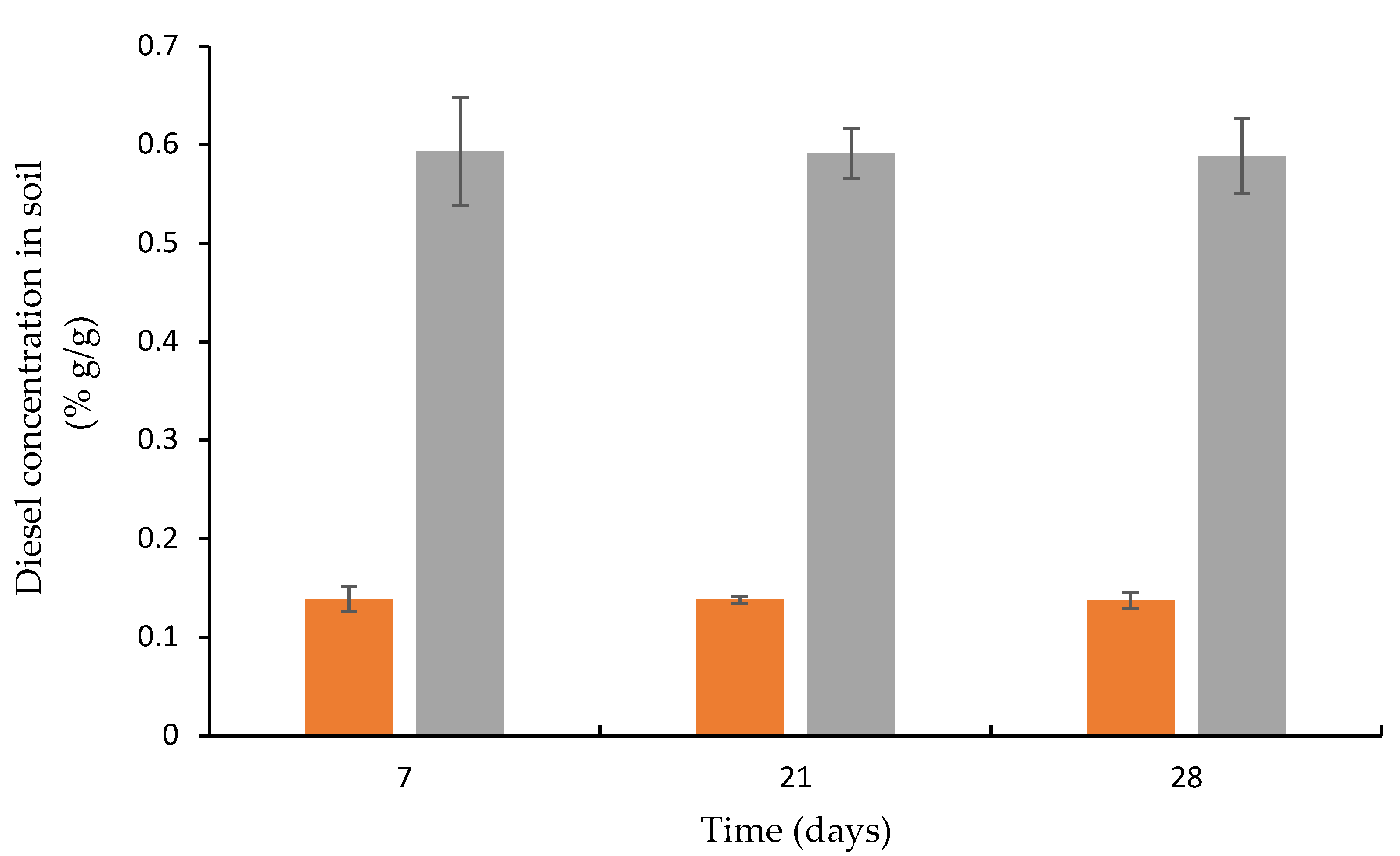{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Families | Soil (%) | Biofilm on the Plasma-Treated Wood Waste (%) | Biofilm on the Untreated Wood Waste (%) | Hydrocarbon Activity Reported Previously |
|---|---|---|---|---|
| Xanthomonadaceae | 4 | 29 | 16 | [47,48] |
| Sphingomonadaceae | 3 | 13 | 8 | [49,50] |
| Chitinophagaceae | 0.29 | 8 | 13 | [51] |
| Rhodospirillaceae | 1 | 7 | 8 | [52,53] |
| Caulobacteraceae | 2 | 6 | 3 | [52] |
| Pseudomonadaceae | 4 | 6 | 4 | [48] |
| Bradyrhizobiaceae | 0.04 | 4 | 3 | [53] |
| mitochondria | 0.22 | 3 | 6 | |
| Oxalobacteraceae | 0.04 | 3 | 3 | [47] |
| Sinobacteraceae | 11 | 2 | 2 | [54] |
| sum | 26.09% | 81% | 66% |
4. Conclusions
Author Contributions
Conflicts of Interest
References
- Gallego, J.L.R.; Loredo, J.; Llamas, J.F.; Vázquez, F.; Sánchez, J. Bioremediation of diesel-contaminated soils: Evaluation of potential in situ techniques by study of bacterial degradation. Biodegradation 2001, 12, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Silverman, D.T.; Samanic, C.M.; Lubin, J.H.; Blair, A.E.; Stewart, P.A.; Vermeulen, R.; Coble, J.B.; Rothman, N.; Schleiff, P.L.; Travis, W.D.; et al. The Diesel Exhaust in Miners Study: A Nested Case-Control Study of Lung Cancer and Diesel Exhaust. JNCI J. Natl. Cancer Inst. 2012, 104, 855–868. [Google Scholar] [CrossRef] [PubMed]
- Khan, F.I.; Husain, T.; Hejazi, R. An overview and analysis of site remediation technologies. J. Environ. Manag. 2004, 71, 95–122. [Google Scholar] [CrossRef] [PubMed]
- Vidali, M. Bioremediation. An overview. Pure Appl. Chem. 2001, 73, 1163–1172. [Google Scholar] [CrossRef]
- Donlan, R.M. Biofilms: microbial life on surfaces. Emerg. Infect. Dis. 2002, 8, 881–890. [Google Scholar] [CrossRef]
- Swissa, N.; Nitzan, Y.; Anker, Y.; Cahan, R. Biofilter based on a biofilm immobilized on geo-textile sheets for rapid atrazine biodegradation. Int. Biodeterior. Biodegrad. 2015, 105, 146–152. [Google Scholar] [CrossRef]
- Gomila, M.; Busquets, A.; García-Valdés, E.; Michael, E.; Cahan, R.; Nitzan, Y.; Lalucat, J. Draft Genome Sequence of the Toluene-Degrading Pseudomonas stutzeri Strain ST-9. Genome Announc. 2015, 3, e00567-15. [Google Scholar] [CrossRef] [Green Version]
- Michael, E.; Gomila, M.; Lalucat, J.; Nitzan, Y.; Pechatnikov, I.; Cahan, R. Proteomic Assessment of the Expression of Genes Related to Toluene Catabolism and Porin Synthesis in Pseudomonas stutzeri ST-9. J. Proteome Res. 2017, 16, 1683–1692. [Google Scholar] [CrossRef]
- Innemanová, P.; Filipová, A.; Michalíková, K.; Wimmerová, L.; Cajthaml, T. Bioaugmentation of PAH-contaminated soils: A novel procedure for introduction of bacterial degraders into contaminated soil. Ecol. Eng. 2018, 118, 93–96. [Google Scholar] [CrossRef]
- Sun, G.-D.; Xu, Y.; Jin, J.-H.; Zhong, Z.-P.; Liu, Y.; Luo, M.; Liu, Z.-P. Pilot scale ex-situ bioremediation of heavily PAHs-contaminated soil by indigenous microorganisms and bioaugmentation by a PAHs-degrading and bioemulsifier-producing strain. J. Hazard. Mater. 2012, 233–234, 72–78. [Google Scholar] [CrossRef]
- Tahhan, R.A.; Ammari, T.G.; Goussous, S.J.; Al-Shdaifat, H.I. Enhancing the biodegradation of total petroleum hydrocarbons in oily sludge by a modified bioaugmentation strategy. Int. Biodeterior. Biodegrad. 2011, 65, 130–134. [Google Scholar] [CrossRef]
- Teng, Y.; Luo, Y.; Sun, M.; Liu, Z.; Li, Z.; Christie, P. Effect of bioaugmentation by Paracoccus sp. strain HPD-2 on the soil microbial community and removal of polycyclic aromatic hydrocarbons from an aged contaminated soil. Bioresour. Technol. 2010, 101, 3437–3443. [Google Scholar] [PubMed] [Green Version]
- Venkata Mohan, S.; Prasanna, D.; Purushotham Reddy, B.; Sarma, P.N. Ex situ bioremediation of pyrene contaminated soil in bio-slurry phase reactor operated in periodic discontinuous batch mode: Influence of bioaugmentation. Int. Biodeterior. Biodegrad. 2008, 62, 162–169. [Google Scholar] [CrossRef]
- Thomassin-Lacroix, E.; Eriksson, M.; Reimer, K.; Mohn, W. Biostimulation and bioaugmentation for on-site treatment of weathered diesel fuel in Arctic soil. Appl. Microbiol. Biotechnol. 2002, 59, 551–556. [Google Scholar]
- Launen, L.A.; Buggs, V.H.; Eastep, M.E.; Enriquez, R.C.; Leonard, J.W.; Blaylock, M.J.; Huang, J.-W.; Häggblom, M.M. Bioremediation of Polyaromatic Hydrocarbon-Contaminated Sediments in Aerated Bioslurry Reactors. Bioremediat. J. 2002, 6, 125–141. [Google Scholar] [CrossRef]
- Silva, Í.S.; dos Santos, E.D.; de Menezes, C.R.; de Faria, A.F.; Franciscon, E.; Grossman, M.; Durrant, L.R. Bioremediation of a polyaromatic hydrocarbon contaminated soil by native soil microbiota and bioaugmentation with isolated microbial consortia. Bioresour. Technol. 2009, 100, 4669–4675. [Google Scholar] [CrossRef]
- Mrozik, A.; Piotrowska-Seget, Z. Bioaugmentation as a strategy for cleaning up of soils contaminated with aromatic compounds. Microbiol. Res. 2010, 165, 363–375. [Google Scholar] [CrossRef]
- Kjelleberg, S.; Molin, S. Is there a role for quorum sensing signals in bacterial biofilms? Curr. Opin. Microbiol. 2002, 5, 254–258. [Google Scholar] [CrossRef]
- Stoodley, P.; Cargo, R.; Rupp, C.J.; Wilson, S.; Klapper, I. Biofilm material properties as related to shear-induced deformation and detachment phenomena. J. Ind. Microbiol. Biotechnol. 2002, 29, 361–367. [Google Scholar] [CrossRef]
- Kumar, C.G.; Anand, S.K. Significance of microbial biofilms in food industry: a review. Int. J. Food Microbiol. 1998, 42, 9–27. [Google Scholar] [CrossRef]
- Koch, B.; Worm, J.; Jensen, L.E.; Højberg, O.; Nybroe, O. Carbon limitation induces sigma(S)-dependent gene expression in Pseudomonas fluorescens in soil. Appl. Environ. Microbiol. 2001, 67, 3363–3370. [Google Scholar] [CrossRef] [PubMed]
- Schechter, M.; Schechter, A.; Rozenfeld, S.; Efrat, E.; Cahan, R. Anode Biofilm. In Technology and Application of Microbial Fuel Cells; Chin-Tgan, W., Ed.; IntechOpen: London, UK, 2014; pp. 57–75. [Google Scholar]
- Svensson, S.L.; Pryjma, M.; Gaynor, E.C. Flagella-Mediated Adhesion and Extracellular DNA Release Contribute to Biofilm Formation and Stress Tolerance of Campylobacter jejuni. PLoS ONE 2014, 9, e106063. [Google Scholar] [CrossRef] [PubMed]
- Farber, R.; Dabush-Busheri, I.; Chaniel, G.; Rozenfeld, S.; Bormashenko, E.; Multanen, V.; Cahan, R. Biofilm grown on wood waste pretreated with cold low-pressure nitrogen plasma: Utilization for toluene remediation. Int. Biodeterior. Biodegradation 2019, 139, 62–69. [Google Scholar] [CrossRef]
- Dursun, A.Y.; Tepe, O. Internal mass transfer effect on biodegradation of phenol by Ca-alginate immobilized Ralstonia eutropha. J. Hazard. Mater. 2005, 126, 105–111. [Google Scholar] [CrossRef]
- Abu-Salah, K.; Shelef, G.; Levanon, D.; Armon, R.; Dosoretz, C.G. Microbial degradation of aromatic and polyaromatic toxic compounds adsorbed on powdered activated carbon. J. Biotechnol. 1996, 51, 265–272. [Google Scholar] [CrossRef]
- Yordanova, G.; Ivanova, D.; Godjevargova, T.; Krastanov, A. Biodegradation of phenol by immobilized Aspergillus awamori NRRL 3112 on modified polyacrylonitrile membrane. Biodegradation 2009, 20, 717–726. [Google Scholar] [CrossRef]
- Cahan, R.; Stein, M.; Anker, Y.; Langzam, Y.; Nitzan, Y. Innovative utilization of coal bottom ash for bioremediation of toxic organic pollutants. Int. Biodeterior. Biodegrad. 2013, 85, 421–428. [Google Scholar] [CrossRef]
- Bormashenko, E.; Chaniel, G.; Grynyov, R. Towards understanding hydrophobic recovery of plasma treated polymers: Storing in high polarity liquids suppresses hydrophobic recovery. Appl. Surf. Sci. 2013, 273, 549–553. [Google Scholar] [CrossRef]
- Carter, M.R.; Gregorich, E.G. Soil Sampling and Methods of Analysis, 2nd ed.; Taylor & Francis Group, LLC: Boca Raton, FL, USA, 2008. [Google Scholar]
- Sayara, T.; Sarrà, M.; Sánchez, A. Preliminary screening of co-substrates for bioremediation of pyrene-contaminated soil through composting. J. Hazard. Mater. 2009, 172, 1695–1698. [Google Scholar] [CrossRef] [Green Version]
- Sayara, T.; Sarrà, M.; Sánchez, A. Effects of compost stability and contaminant concentration on the bioremediation of PAHs-contaminated soil through composting. J. Hazard. Mater. 2010, 179, 999–1006. [Google Scholar] [CrossRef] [Green Version]
- Betancur-Galvis, L.A.; Alvarez-Bernal, D.; Ramos-Valdivia, A.C.; Dendooven, L. Bioremediation of polycyclic aromatic hydrocarbon-contaminated saline–alkaline soils of the former Lake Texcoco. Chemosphere 2006, 62, 1749–1760. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Chi, X.-Q.; Zhang, J.-J.; Sun, D.-L.; Zhou, N.-Y. Bioaugmentation of a methyl parathion contaminated soil with Pseudomonas sp. strain WBC-3. Int. Biodeterior. Biodegrad. 2014, 87, 116–121. [Google Scholar] [CrossRef]
- Ghazali, F.M.; Rahman, R.N.Z.A.; Salleh, A.B.; Basri, M. Biodegradation of hydrocarbons in soil by microbial consortium. Int. Biodeterior. Biodegrad. 2004, 54, 61–67. [Google Scholar] [CrossRef]
- Rodrigues, J.L.M.; Kachel, C.A.; Aiello, M.R.; Quensen, J.F.; Maltseva, O.V.; Tsoi, T.V.; Tiedje, J.M. Degradation of Aroclor 1242 Dechlorination Products in Sediments by Burkholderia xenovorans LB400(ohb) and Rhodococcus sp. Strain RHA1(fcb). Appl. Environ. Microbiol. 2006, 72, 2476–2482. [Google Scholar] [CrossRef]
- Nwankwegu, A.S.; Onwosi, C.O. Bioremediation of gasoline contaminated agricultural soil by bioaugmentation. Environ. Technol. Innov. 2017, 7, 1–11. [Google Scholar] [CrossRef]
- Poi, G.; Aburto-Medina, A.; Mok, P.C.; Ball, A.S.; Shahsavari, E. Large scale bioaugmentation of soil contaminated with petroleum hydrocarbons using a mixed microbial consortium. Ecol. Eng. 2017, 102, 64–71. [Google Scholar] [CrossRef]
- Gallo, M.; Speranza, B.; Corbo, M.R.; Sinigaglia, M.; Bevilacqua, A. Novel Microbial Immobilization Techniques. In Novel Food Fermentation Technologies; Springer: Cham, Switzerland, 2016; pp. 35–55. [Google Scholar]
- Das, N.; Chandran, P. Microbial Degradation of Petroleum Hydrocarbon Contaminants: An Overview. Biotechnol. Res. Int. 2011, 2011, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Kuyukina, M.S.; Ivshina, I.B.; Serebrennikova, M.K.; Krivorutchko, A.B.; Podorozhko, E.A.; Ivanov, R.V.; Lozinsky, V.I. Petroleum-contaminated water treatment in a fluidized-bed bioreactor with immobilized Rhodococcus cells. Int. Biodeterior. Biodegrad. 2009, 63, 427–432. [Google Scholar] [CrossRef]
- Chen, W.; Li, J.; Sun, X.; Min, J.; Hu, X. High efficiency degradation of alkanes and crude oil by a salt-tolerant bacterium Dietzia species CN-3. Int. Biodeterior. Biodegrad. 2017. [Google Scholar] [CrossRef]
- Freidman, B.L.; Gras, S.L.; Snape, I.; Stevens, G.W.; Mumford, K.A. The performance of ammonium exchanged zeolite for the biodegradation of petroleum hydrocarbons migrating in soil water. J. Hazard. Mater. 2016, 313, 272–282. [Google Scholar] [CrossRef]
- Wang, X.; Wang, X.; Liu, M.; Bu, Y.; Zhang, J.; Chen, J.; Zhao, J. Adsorption–synergic biodegradation of diesel oil in synthetic seawater by acclimated strains immobilized on multifunctional materials. Mar. Pollut. Bull. 2015, 92, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Margesin, R.; Labbé, D.; Schinner, F.; Greer, C.W.; Whyte, L.G. Characterization of Hydrocarbon-Degrading Microbial Populations in Contaminated and Pristine Alpine Soils. Appl. Environ. Microbiol. 2003, 69, 3085–3092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehetre, G.T.; Dastager, S.G.; Dharne, M.S. Biodegradation of mixed polycyclic aromatic hydrocarbons by pure and mixed cultures of biosurfactant producing thermophilic and thermo-tolerant bacteria. Sci. Total Environ. 2019, 679, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Bell, T.H.; Yergeau, E.F.; Juck, D.G.; Whyte, L.W.; Greer, C. Alteration of microbial community structure affects diesel biodegradation in an Arctic soil. FEMS Microbiol. Ecol. 2013, 85, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.; Choi, S.; Hong, H.; Sung, J.-S.; Park, W. Effect of Red Clay on Diesel Bioremediation and Soil Bacterial Community. Microb. Ecol. 2014, 68, 314–323. [Google Scholar] [CrossRef]
- Gran-Scheuch, A.; Fuentes, E.; Bravo, D.M.; Jiménez, J.C.; Pérez-Donoso, J.M. Isolation and Characterization of Phenanthrene Degrading Bacteria from Diesel Fuel-Contaminated Antarctic Soils. Front. Microbiol. 2017, 8, 1634. [Google Scholar] [CrossRef] [Green Version]
- Garrido-Sanz, D.; Redondo-Nieto, M.; Guirado, M.; Pindado Jiménez, O.; Millán, R.; Martin, M.; Rivilla, R. Metagenomic Insights into the Bacterial Functions of a Diesel-Degrading Consortium for the Rhizoremediation of Diesel-Polluted Soil. Genes Basel 2019, 10. [Google Scholar] [CrossRef]
- Sarkar, J.; Kazy, S.K.; Gupta, A.; Dutta, A.; Mohapatra, B.; Roy, A.; Bera, P.; Mitra, A.; Sar, P. Biostimulation of Indigenous Microbial Community for Bioremediation of Petroleum Refinery Sludge. Front. Microbiol. 2016, 7, 1407. [Google Scholar] [CrossRef] [Green Version]
- Fuentes, S.; Barra, B.; Caporaso, J.G.; Seeger, M. From Rare to Dominant: A Fine-Tuned Soil Bacterial Bloom during Petroleum Hydrocarbon Bioremediation. Appl. Environ. Microbiol. 2016, 82, 888–896. [Google Scholar] [CrossRef]
- Jung, J.; Philippot, L.; Park, W. Metagenomic and functional analyses of the consequences of reduction of bacterial diversity on soil functions and bioremediation in diesel-contaminated microcosms. Sci. Rep. 2016, 6, 23012. [Google Scholar] [CrossRef] [Green Version]
- Korotkevych, O.; Josefiova, J.; Praveckova, M.; Cajthaml, T.; Stavelova, M.; Brennerova, M.V. Functional adaptation of microbial communities from jet fuel-contaminated soil under bioremediation treatment: simulation of pollutant rebound. FEMS Microbiol. Ecol. 2011, 78, 137–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prince, R.C.; Gramain, A.; McGenity, T.J. Prokaryotic Hydrocarbon Degraders. In Handbook of Hydrocarbon and Lipid Microbiology; Springer Berlin Heidelberg: Berlin, Heidelberg, 2010; pp. 1669–1692. [Google Scholar]
- Yang, S.; Wen, X.; Jin, H.; Wu, Q. Pyrosequencing Investigation into the Bacterial Community in Permafrost Soils along the China-Russia Crude Oil Pipeline (CRCOP). PLoS ONE 2012, 7, e52730. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, S.; Méndez, V.; Aguila, P.; Seeger, M. Bioremediation of petroleum hydrocarbons: catabolic genes, microbial communities, and applications. Appl. Microbiol. Biotechnol. 2014, 98, 4781–4794. [Google Scholar] [CrossRef] [PubMed]
- Popp, N.; Schlomann, M.; Mau, M. Bacterial diversity in the active stage of a bioremediation system for mineral oil hydrocarbon-contaminated soils. Microbiology 2006, 152, 3291–3304. [Google Scholar] [CrossRef]
- Hamamura, N.; Ward, D.M.; Inskeep, W.P. Effects of petroleum mixture types on soil bacterial population dynamics associated with the biodegradation of hydrocarbons in soil environments. FEMS Microbiol. Ecol. 2013, 85, 168–178. [Google Scholar] [CrossRef] [Green Version]
- Ferrenberg, S.; O’neill, S.P.; Knelman, J.E.; Todd, B.; Duggan, S.; Bradley, D.; Robinson, T.; Schmidt, S.K.; Townsend, A.R.; Williams, M.W.; et al. Changes in assembly processes in soil bacterial communities following a wildfire disturbance. ISME J. 2013, 7, 1102–1111. [Google Scholar] [CrossRef] [Green Version]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Farber, R.; Rosenberg, A.; Rozenfeld, S.; Banet, G.; Cahan, R. Bioremediation of Artificial Diesel-Contaminated Soil Using Bacterial Consortium Immobilized to Plasma-Pretreated Wood Waste. Microorganisms 2019, 7, 497. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7110497
Farber R, Rosenberg A, Rozenfeld S, Banet G, Cahan R. Bioremediation of Artificial Diesel-Contaminated Soil Using Bacterial Consortium Immobilized to Plasma-Pretreated Wood Waste. Microorganisms. 2019; 7(11):497. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7110497
Chicago/Turabian StyleFarber, Ravit, Alona Rosenberg, Shmuel Rozenfeld, Gabi Banet, and Rivka Cahan. 2019. "Bioremediation of Artificial Diesel-Contaminated Soil Using Bacterial Consortium Immobilized to Plasma-Pretreated Wood Waste" Microorganisms 7, no. 11: 497. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7110497